Crocin Improves the Quality of Cryopreserved Goat Semen in Different Breeds

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farm, Animals and Management
2.2. Semen Collection
2.3. Assessment of Sperm Motility
2.4. Assessment of Sperm Viability and Morphology by Trypan Blue/Giemsa Technique
2.5. Assessment of Sperm Membrane Integrity
2.6. Evaluation of DNA Fragmentation by Tunel Assay
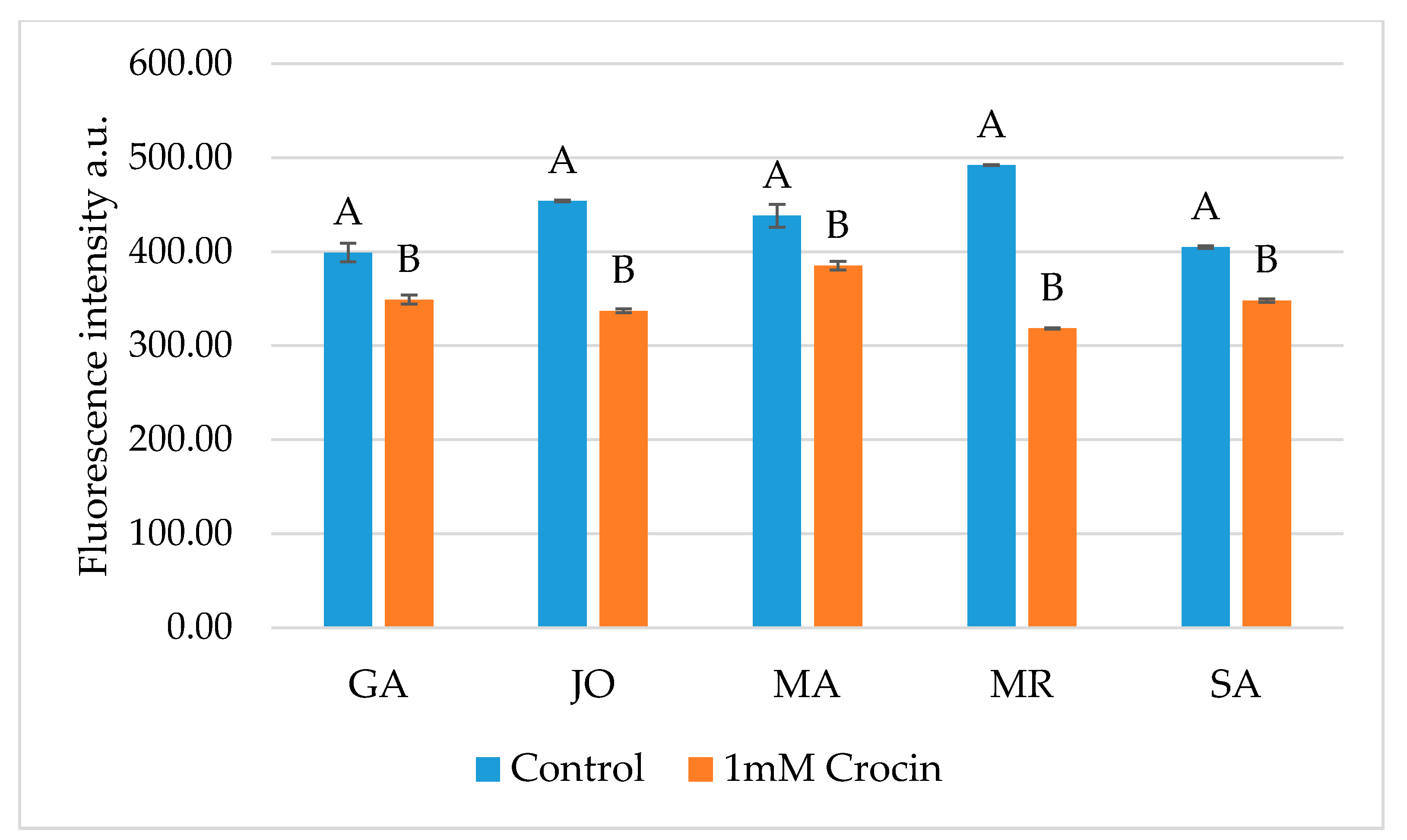
2.7. Evaluation of ROS Levels
2.8. Experimental Design
2.9. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Leboeuf, B.; Restall, B.; Salamon, S. Production and storage of goat semen for artificial insemination. Anim. Reprod. Sci. 2000, 62, 113–141. [Google Scholar] [CrossRef]
- Gama, L.; Bressan, M.C. Biotechnology applications for the sustainable management of goat genetic resources. Small Rumin. Res. 2011, 98, 133–146. [Google Scholar] [CrossRef]
- Gangwar, C.; Kharche, S.D.; Kumar, S.; Jindal, S.K. Cryopreservation of goat semen: Status and prospects. Indian J. Small Rumin. 2016, 22, 1–10. [Google Scholar] [CrossRef]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar] [CrossRef]
- Al-Ghalban, A.M.; Tabaa, M.J.; Kridli, R.T. Factors affecting semen characteristics and scrotal circumference in Damascus bucks. Small Rumin. Res. 2004, 53, 141–149. [Google Scholar] [CrossRef]
- Arrebola, F.; Abecia, J.A. Effects of season and artificial photoperiod on semen and seminal plasma characteristics in bucks of two goat breeds maintained in a semen collection center. Vet. World 2017, 10, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, M.S.; Khan, D.; Mustaq, A.; Afridi, S.S. Effect of extenders, post dilution intervals and seasons on semen quality in dairy goats. Turk. J. Vet. Anim. Sci. 2013, 35, 147–152. [Google Scholar] [CrossRef]
- Hanmante, A.A.; Barbind, R.P.; Mule, R.S.; Baswade, S.V.; Andhare, B.C. Effect of management system on semen quality attributes of Osmanabadi goat bucks. Indian J. Small Rumin. 2009, 15, 115–117. [Google Scholar]
- Abdi-Benemar, H.; Khalili, B.; Zamiri, M.J.; Ezazi, H. Seasonal variation in seminal characteristics, testicular measurements and plasma testosterone concentration in Iranian Khalkhali bucks. J. Livest. Sci. Technol. 2018, 6, 33–39. [Google Scholar] [CrossRef]
- Ritar, A.J.; Salamon, S. Effects of seminal plasma and of its removal and of egg yolk in the diluents on the survival of fresh and frozen-thawed spermatozoa of the Angora goat. Aust. J. Biol. Sci. 1982, 35, 305–312. [Google Scholar] [CrossRef]
- Ferreira, V.; Mello, M.; Fonseca, C.; Dias, Á.; Cardoso, J.; Silva, R.; Junior, W. Effect of seminal plasma and egg yolk concentration on freezability of goat semen. Rev. Bras. De Zootec. 2014, 43, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Majzoub, A. Laboratory tests for oxidative stress. Indian J. Urol 2017, 33, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Tuncer, P.B.; Sarıözkan, S.; Başpınar, N.; Taşpınar, M.; Coyan, K.; Bilgili, A.; Akalın, P.P.; Büyükleblebici, S.; Aydos, S.; et al. Effects of antioxidants on post-thawed bovine sperm and oxidative stress parameters: Antioxidants protect DNA integrity against cryodamage. Cryobiology 2010, 61, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Estrada, E.; Rodríguez-Gil, J.E.; Rocha, L.G.; Balasch, S.; Bonet, S.; Yeste, M. Supplementing cryopreservation media with reduced glutathione increases fertility and prolificacy of sows inseminated with frozen-thawed boar semen. Andrology 2014, 2, 88–99. [Google Scholar] [CrossRef]
- Longobardi, V.; Zullo, G.; Salzano, A.; De Canditiis, C.; Cammarano, A.; De Luise, L.; Puzio, M.V.; Neglia, G.; Gasparrini, B. Resveratrol prevents capacitation-like changes and improves in vitro fertilizing capability of buffalo frozen-thawed sperm. Theriogenology 2017, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nouri, H.; Shojaeian, K.; Samadian, F.; Lee, S.; Kohram, H.; Lee, J.I. Using resveratrol and Epigallocatechin-3-Gallate to improve cryopreservation of stallion spermatozoa with low quality. J. Equine Vet. Sci. 2018, 70, 18–25. [Google Scholar] [CrossRef]
- Sánchez-Rubio, F.; Fernández-Santos, M.; Castro-Vázquez, L.; García-Álvarez, O.; Maroto-Morales, A.; Soler, A.J.; Martínez-Pastor, F.; Garde, J.J. Cinnamtannin B-1, a novel antioxidant for sperm in red deer. Anim. Reprod. Sci. 2018, 195, 44–52. [Google Scholar] [CrossRef]
- Daramola, J.O.; Adekunle, E.O. Cryosurvival of goat spermatozoa in Tris-egg yolk extender supplemented with vitamin C. Arch. Zootec. 2015, 64, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Larbi, A.; Wu, G.; Hong, Q.; Quan, G. Improving the quality of cryopreserved goat semen with a commercial bull extender supplemented with resveratrol. Anim. Reprod. Sci. 2019, 208, 106127. [Google Scholar] [CrossRef]
- Seifi-Jamadi, A.; Ahmad, E.; Ansari, M.; Kohram, H. Antioxidant effect of quercetin in an extender containing DMA or glycerol on freezing capacity of goat semen. Cryobiology 2017, 75, 15–20. [Google Scholar] [CrossRef]
- Ghyasvand, T.; Goodarzi, M.T.; Amiri, I.; Karimi, J.; Ghorbani, M. Serum levels of lycopene, beta-carotene, and retinol and their correlation with sperm DNA damage in normospermic and infertile men. Int. J. Reprod. Biomed. (Yazd) 2015, 13, 787–792. [Google Scholar] [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Zervos, I.; Tsantarliotou, M. Antioxidant effect of crocin on bovine sperm quality and in vitro fertilization. Theriogenology 2015, 84, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Rahaiee, S.; Moini, S.; Hashemi, M.; Shojaosadati, S.A. Evaluation of antioxidant activities of bioactive compounds and various extracts obtained from saffron (Crocus sativus L.): A review. J. Food Sci. Technol. 2015, 52, 1881–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Rebolledo, A.E.; Fernández-Santos, M.R.; Bisbal, A.; Ros-Santaella, J.L.; Ramón, M.; Carmona, M.; Martínez-Pastor, F.; Garde, J.J. Improving the effect of incubation and oxidative stress on thawed spermatozoa from red deer by using different antioxidant treatments. Reprod. Fertil. Dev. 2010, 22, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Mata-Campuzano, M.; Álvarez-Rodríguez, M.; Álvarez, M.; Tamayo-Canul, J.; Anel, L.; de Paz, P.; Martínez-Pastor, F. Post-thawing quality and incubation resilience of cryopreserved ram spermatozoa are affected by antioxidant supplementation and choice of extender. Theriogenology 2015, 83, 520–528. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids, 6th ed.; National Academy Press: Washington, DC, USA, 2007.
- Garde, J.J.; Soler, A.J.; Cassinello, J.; Crespo, C.; Malo, A.F.; Espeso, G.; Gomendio, M.; Roldan, E.M.S. Sperm cryopreservation in three species of endangered gazelles (Gazella cuvieri, G. dama mhorr, and G. dorcas neglecta). Biol. Reprod. 2003, 69, 602–611. [Google Scholar] [CrossRef] [Green Version]
- Boccia, L.; Di Palo, R.; De Rosa, A.; Attanasio, L.; Mariotti, E.; Gasparrini, B. Evaluation of buffalo semen by Trypan blue/Giemsa staining and related fertility in vitro. Ital. J. Anim. Sci. 2007, 6, 739–742. [Google Scholar] [CrossRef]
- World Health Organization Department of Reproductive Health and Research. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO: Geneva, Switzerland, 2010; Available online: http://www.who.int/reproductivehealth/publications/infertility/9789241547789/en/ (accessed on 25 February 2010).
- Correa, J.R.; Zavos, P.M. The Hypoosmotic swelling test: Its employment as an assay to evaluate the functional integrity of the frozen-thawed bovine sperm membrane. Theriogenology 1994, 42, 351–360. [Google Scholar] [CrossRef]
- Nazarewicz, R.; Bikineyeva, A.; Dikalov, S. Rapid and specific measurements of superoxide using fluorescence spectroscopy. J. Biomol. Screen 2013, 18, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Karagiannidis, A.; Varsakeli, S.; Karatzas, G. Characteristics and seasonal variations in the semen of Alpine, Saanen and Damascus goat bucks born and raised in Greece. Theriogenology 2000, 53, 1285–1293. [Google Scholar] [CrossRef]
- Sultana, F.; Husain, S.S.; Khatun, A.; Apu, A.S.; Khandoker, M.A.M. Study on buck evaluation based on semen quality and fertility. Bang. J. Anim. Sci. 2013, 42, 101–108. [Google Scholar] [CrossRef]
- Delgadillo, J.A.; Fitz-Rodríguez, G.; Duarte, G.; Véliz, F.G.; Carrillo, E.; Flores, J.A.; Vielma, J.; Hernandez, H.; Malpaux, B. Management of photoperiod to control caprine reproduction in the subtropics. Reprod. Fertil. Dev. 2004, 16, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.; Maxwell, W.M.C. Salamon’s Artificial Insemination of Sheep and Goats; Buttetworths: Sydney, Australia, 1987; pp. 85–91. [Google Scholar] [CrossRef]
- Saacke, R.G.; White, J.M. Semen quality tests and their relationship to fertility. In Proceedings of the 4th National Association of Animal Breeders, Technology Conference Artificial Insemination and Reproduction; National Association of Animal Breeders: Columbia, MO, USA, 1972; pp. 22–27. [Google Scholar]
- Adeniji, D.A.; Oyeyemi, M.O.; Olugbemi, J.B. Sperm morphological characteristic and mating behaviour of Proviron® treated west African dwarf bucks with testicular degeneration. Int. J. Morphol. 2010, 28, 623–626. [Google Scholar] [CrossRef]
- Barbas, J.; Mascarenhas, R. Cryopreservation of domestic animal sperm cells. Cell Tissue Bank. 2009, 1, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dorado, J.; Munoz-Serrano, A.; Hidalgo, M. The effect of cryopreservation on goat semen characteristics related to sperm freezability. Anim. Reprod. Sci. 2010, 121, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Simões, R.; Feitosa, W.B.; Siqueira, A.F.; Nichi, M.; Paula-Lopes, F.F.; Marques, M.G.; Peres, M.A.; Barnabe, V.H.; Visintin, J.A.; Assumpção, M.E. Influence of bovine sperm DNA fragmentation and oxidative stress on early embryo in vitro development outcome. Reproduction 2013, 146, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Üstüner, B.; Nur, Z.; Alçay, S.; Toker, M.B.; Sağirkaya, H.; Soylu, M.K. Effect of freezing rate on goat sperm morphology and DNA integrity. Turk. J. Vet. Anim. Sci. 2015, 39, 110–114. [Google Scholar] [CrossRef]
- Vijayaraghavan, S. Sperm Motility: Patterns and Regulation. In Introduction to Mammalian Reproduction; Springer: Boston, MA, USA, 2003; pp. 79–91. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Caballero-Ortega, H.; Riveron-Nigrete, L.; Pereda-miranda, R.; Rivera- Luna, R.; Manuel Hernandez, J.; Perez-Lopez, I.; Espinosa-Aguirre, J.J. In vitro evaluation of chemopreventive potential of saffron. Rev. Invest. Clin. 2002, 54, 430–436. [Google Scholar]
- Mardani, M.; Vaez, A.; Razavi, S. Effect of saffron on rat sperm chromatin integrity. Int. J. Reprod. Biomed. 2014, 12, 343–350. [Google Scholar]
- Mehdipour, M.; Kia, D.H.; Najafi, A. Effect of crocin and naringenin supplementation in cryopreservation medium on post-thawed rooster sperm quality and expression of apoptosis associated genes. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Tsantarliotou, M.P.; Poutahidis, T.; Markala, D.; Kazakos, G.; Sapanidou, V.; Lavrentiadou, S.; Zervos, I.; Taitzoglou, I.; Sinakos, Z. Crocetin administration ameliorates endotoxin-induced disseminated intravascular coagulation in rabbits. Blood Coagul. Fibrinolysis 2013, 24, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Martorana, K.; Klooster, K.; Meyers, S. Suprazero cooling rate, rather than freezing rate, determines post thaw quality of rhesus macaque sperm. Theriogenology 2014, 81, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualtieri, R.; Barbato, V.; Fiorentino, I.; Braun, S.; Rizos, D.; Longobardi, S.; Talevi, R. Treatment with zinc, d-aspartate, and co-enzyme Q10 protects bull sperm against damage and improves their ability to support embryo development. Theriogenology 2014, 82, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Mokhber Maleki, E.; Eimani, H.; Bigdeli, M.R.; Golkar Narenji, A.; Abedi, R. Effects of crocin supplementation during in vitro maturation of mouse oocytes on glutathione synthesis and cytoplasmic maturation. Int. J. Fertil. Steril. 2016, 10, 53–61. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Crocin | Motility (%) | AIL * (%) | ALL + ALD ** (%) | HOS+ *** (%) | Tunel+ **** (%) |
|---|---|---|---|---|---|
| 0 mM | 63.1 ± 0.2 A | 74.8 ± 1.8 | 25.2 ± 1.7 | 63.3 ±1.5 | 21.4 ± 4.1 A |
| 0.5 mM | 65.0 ± 1.2 A | 75.2 ± 1.9 | 24.8 ± 1.8 | 62.9 ± 2.3 | 18.4 ± 2.7 A |
| 1 mM | 73.3 ± 0.5 B | 75.1 ± 2.1 | 24.9 ± 2.0 | 66.3 ± 2.1 | 9.8 ± 1.6 B |
| 2 mM | 57.5 ± 0.8 AC | 74.8 ± 2.0 | 25.2 ± 1.5 | 65.1 ± 1.6 | 22.8 ± 3.4 A |
| Breed | Volume (mL) | Concentration (109/mL) | Motility (%) | AIL (%) | Abnormalities (%) |
|---|---|---|---|---|---|
| GA | 0.7 ± 0.1 B | 6.0 ± 0.9 | 70.0 ± 3.0 | 83.1 ± 2.1 | 21.3 ± 4.5 a |
| JO | 0.9 ± 0.1 AB | 4.7 ± 1.0 | 68.0 ± 1.3 | 76.2 ± 3.9 | 19.5 ± 3.5 a |
| MA | 0.8 ± 0.1 b | 6.5 ± 0.6 a | 70.7 ± 3.6 | 82.0 ± 3.5 | 10.9 ± 2.1 b |
| MR | 1.1 ± 0.1 Aa | 5.8 ± 1.0 | 67.8 ± 5.6 | 85.1 ± 3.6 a | 12.1 ± 2.4 b |
| SA | 0.7 ± 0.1 B | 3.8 ± 0.9 b | 70.5 ± 2.9 | 69.7 ± 6.4 b | 23.0 ± 6.0 a |
| Breed | Group | Motility (%) | AIL * (%) | ALL + ALD ** (%) | HOS+ *** (%) | Tunel+ **** (%) |
|---|---|---|---|---|---|---|
| GA | Fresh | 70.0 ± 3.0 | 83.1 ± 2.1 A | 6.1 ± 1.7 A | 67.8 ± 2.4 | 7.8 ± 2.0 A |
| Frozen | 65.0 ± 1.5 | 70.8 ± 2.9 B | 23.7 ± 2.0 B | 61.0 ± 3.3 | 23.2 ± 4.0 B | |
| JO | Fresh | 68.0 ± 1.3 | 76.2 ± 3.9 | 4.7 ± 1.5 A | 61.0 ± 2.8 | 12.1 ± 3.1 a |
| Frozen | 60.0 ± 3.8 | 70.2 ± 2.4 | 21.8 ± 2.4 B | 58.2 ± 1.9 | 22.8 ± 3.4 b | |
| MA | Fresh | 70.7 ± 3.6 | 82.0 ± 3.5 A | 7.1 ± 2.5 A | 67.3 ± 3.3 | 9.7 ± 2.8 a |
| Frozen | 65.7 ± 2.5 | 67.1 ± 3.2 B | 22.5 ± 2.8 B | 64.9 ± 3.2 | 22.1 ± 4.4 b | |
| MR | Fresh | 67.3 ± 5.6 | 85.1 ± 3.6 a | 3.5 ± 1.0 A | 68.4 ± 2.1 | 9.9 ± 1.6 a |
| Frozen | 66.0 ± 3.4 | 76.0 ± 2.1 b | 15.2 ± 1.6 B | 66.6 ± 3.6 | 20.4 ± 4.6 b | |
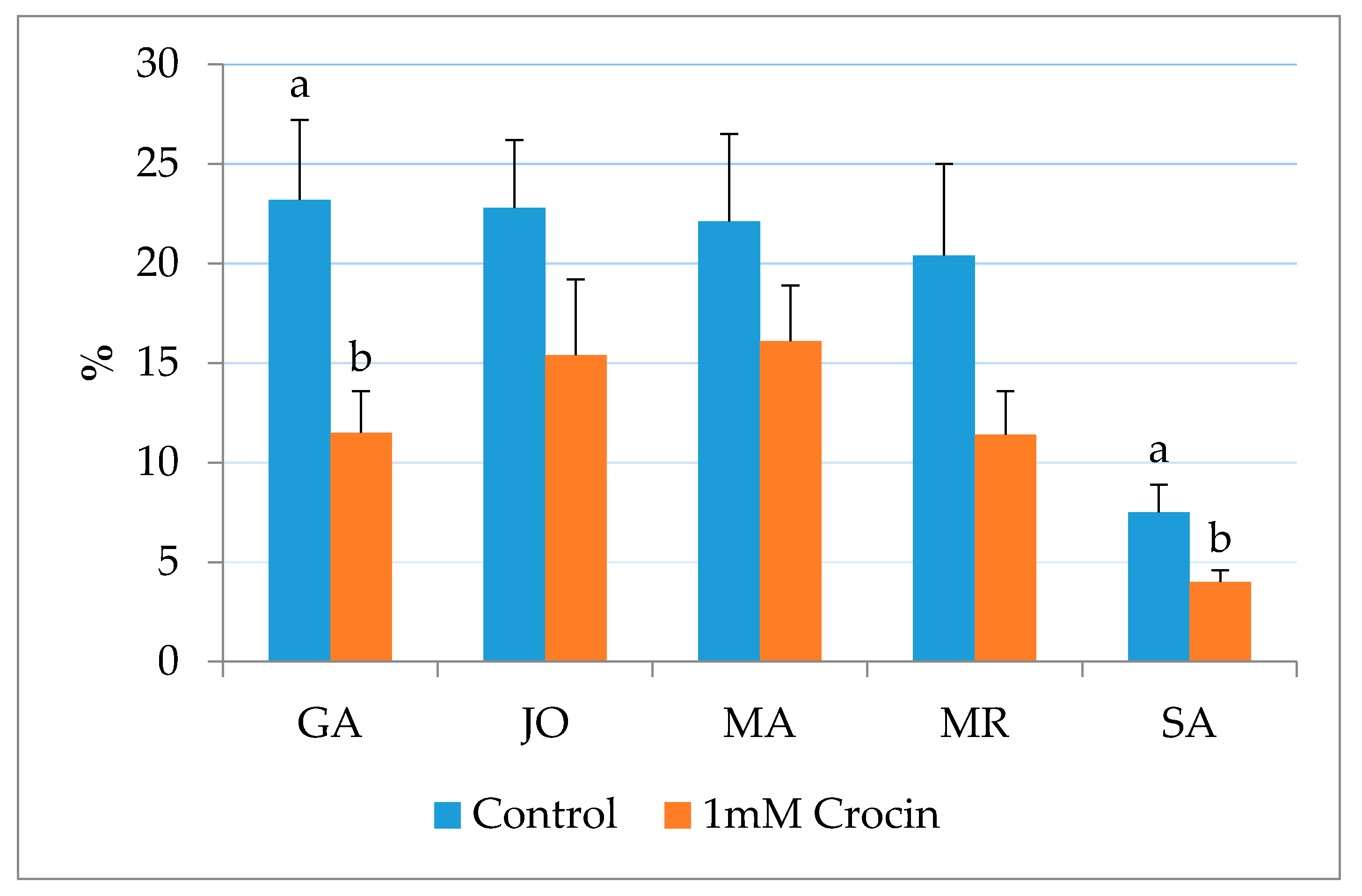
| SA | Fresh | 70.5 ± 2.9 A | 69.7 ± 6.4 | 4.1 ± 1.6 A | 64.6 ± 4.9 | 7.5 ± 1.4 |
| Frozen | 50.0 ± 6.6 B | 62.5 ± 3.2 | 26.0 ± 5.2 B | 61.5 ± 6.1 | 11.6 ± 3.1 | |
| TOTAL | Fresh | 69.3 ± 3.3 a | 79.2. ±3.9 A | 5.1 ± 1.7 A | 65.8 ± 3.1 | 9.4 ± 2.2 A |
| Frozen | 61.3 ± 3.6 b | 69.3 ± 2.8 B | 21.8 ± 2.8 B | 62.4 ± 3.6 | 20.0 ± 3.9 B |
| Breed | Group | Motility (%) | AIL * (%) | ALL + ALD ** (%) | HOS+ *** (%) |
|---|---|---|---|---|---|
| GA | Control | 65.0 ± 1.5 A | 70.8 ± 2.9 | 23.7 ± 2.0 | 61.0 ± 3.3 |
| 1 mM crocin | 74.2 ± 1.8 B | 78.1 ± 3.0 | 18.0 ± 2.7 | 63.2 ± 1.9 | |
| JO | Control | 60.0 ± 3.8 A | 70.2 ± 2.4 | 21.8 ± 2.4 | 58.2 ± 1.9 |
| 1 mM crocin | 74.0 ± 2.7 B | 71.4 ± 1.3 | 23.8 ± 1.0 | 65.0 ± 3.9 | |
| MA | Control | 65.7 ± 2.5 A | 67.1 ±3.2 | 22.5 ± 2.8 | 64.9 ± 3.2 |
| 1 mM crocin | 74.3 ± 2.0 B | 69.1 ±.2.0 | 24.1 ± 2.0 | 63.6 ± 2.4 | |
| MR | Control | 66.0 ± 3.4 | 76.0 ± 2.1 | 15.2 ± 1.6 | 66.6 ± 3.6 |
| 1 mM crocin | 64.0 ± 1.6 | 76.2 ± 1.9 | 15.0 ± 1.1 | 67.6 ± 2.4 | |
| SA | Control | 50.0 ± 6.6 a | 62.5 ± 3.2 | 26.0 ± 5.2 | 61.5 ± 6.1 |
| 1 mM crocin | 60.0 ± 5.8 b | 73.5 ± 6.0 | 21.5 ± 4.9 | 75.0 ± 3.5 | |
| TOTAL | Control | 61.3 ± 3.6 A | 69.3 ± 2.8 | 21.8 ± 2.8 | 62.4 ± 3.6 |
| 1 mM crocin | 69.3 ± 2.8 B | 73.7 ± 2.9 | 20.5 ± 2.3 | 66.9 ± 2.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longobardi, V.; Zullo, G.; Cotticelli, A.; Salzano, A.; Albero, G.; Navas, L.; Rufrano, D.; Claps, S.; Neglia, G. Crocin Improves the Quality of Cryopreserved Goat Semen in Different Breeds. Animals 2020, 10, 1101. https://doi.org/10.3390/ani10061101
Longobardi V, Zullo G, Cotticelli A, Salzano A, Albero G, Navas L, Rufrano D, Claps S, Neglia G. Crocin Improves the Quality of Cryopreserved Goat Semen in Different Breeds. Animals. 2020; 10(6):1101. https://doi.org/10.3390/ani10061101
Chicago/Turabian StyleLongobardi, Valentina, Gianluigi Zullo, Alessio Cotticelli, Angela Salzano, Giuseppe Albero, Luigi Navas, Domenico Rufrano, Salvatore Claps, and Gianluca Neglia. 2020. "Crocin Improves the Quality of Cryopreserved Goat Semen in Different Breeds" Animals 10, no. 6: 1101. https://doi.org/10.3390/ani10061101