Effects of Dietary Supplementation of Lauric Acid on Lactation Function, Mammary Gland Development, and Serum Lipid Metabolites in Lactating Mice

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture and Treatment
2.3. Animals and In Vivo Study
2.4. H&E Staining
2.5. Western Blot Analysis
2.6. qRT-PCR
2.7. Lipidomics Analysis
2.8. Statistical Analysis
3. Results
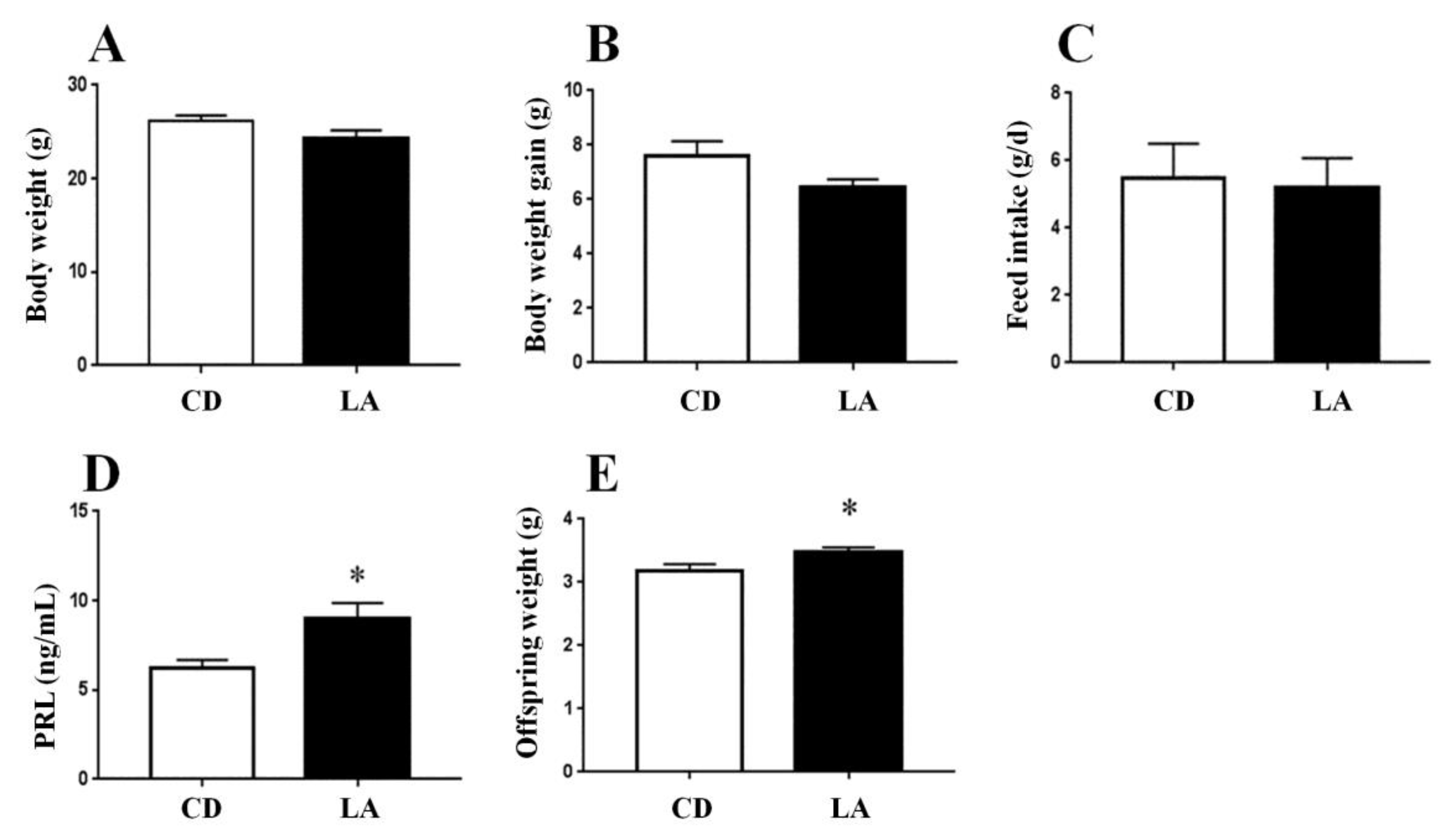
3.1. Dietary Supplementation of LA Improves Lactation Function in Breast-Feeding Mice
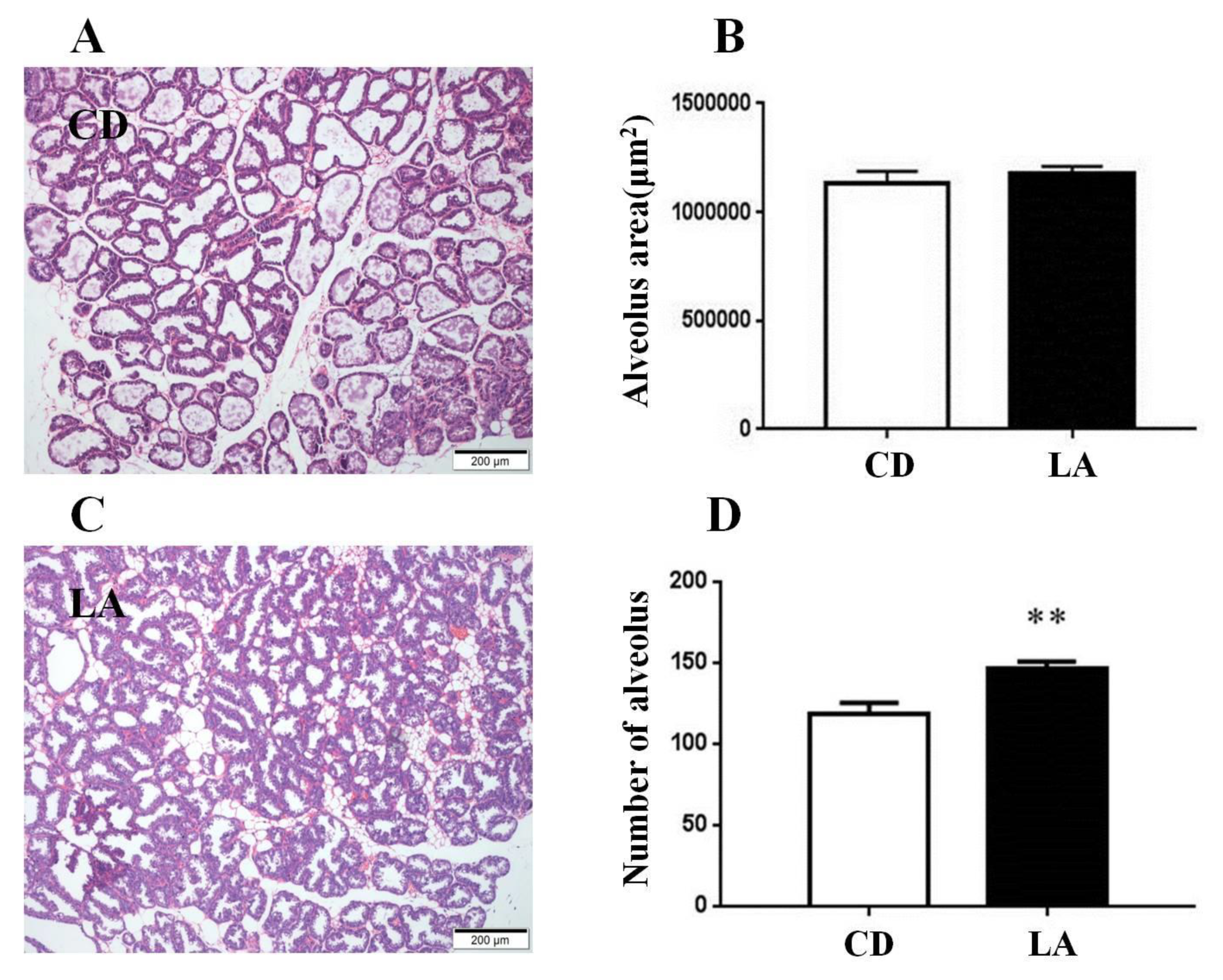
3.2. Dietary Supplementation of LA Promotes Mammary Gland Development in Breast-Feeding Mice
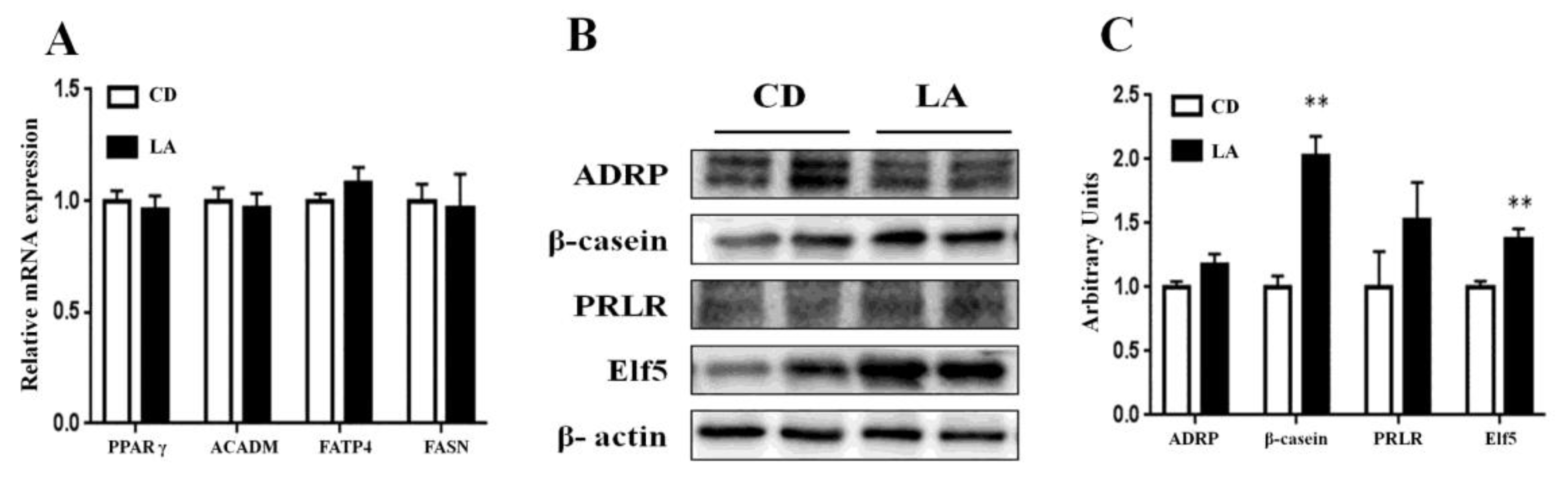
3.3. Gene and Protein Expression in Mammary Glands of Lactating Mice
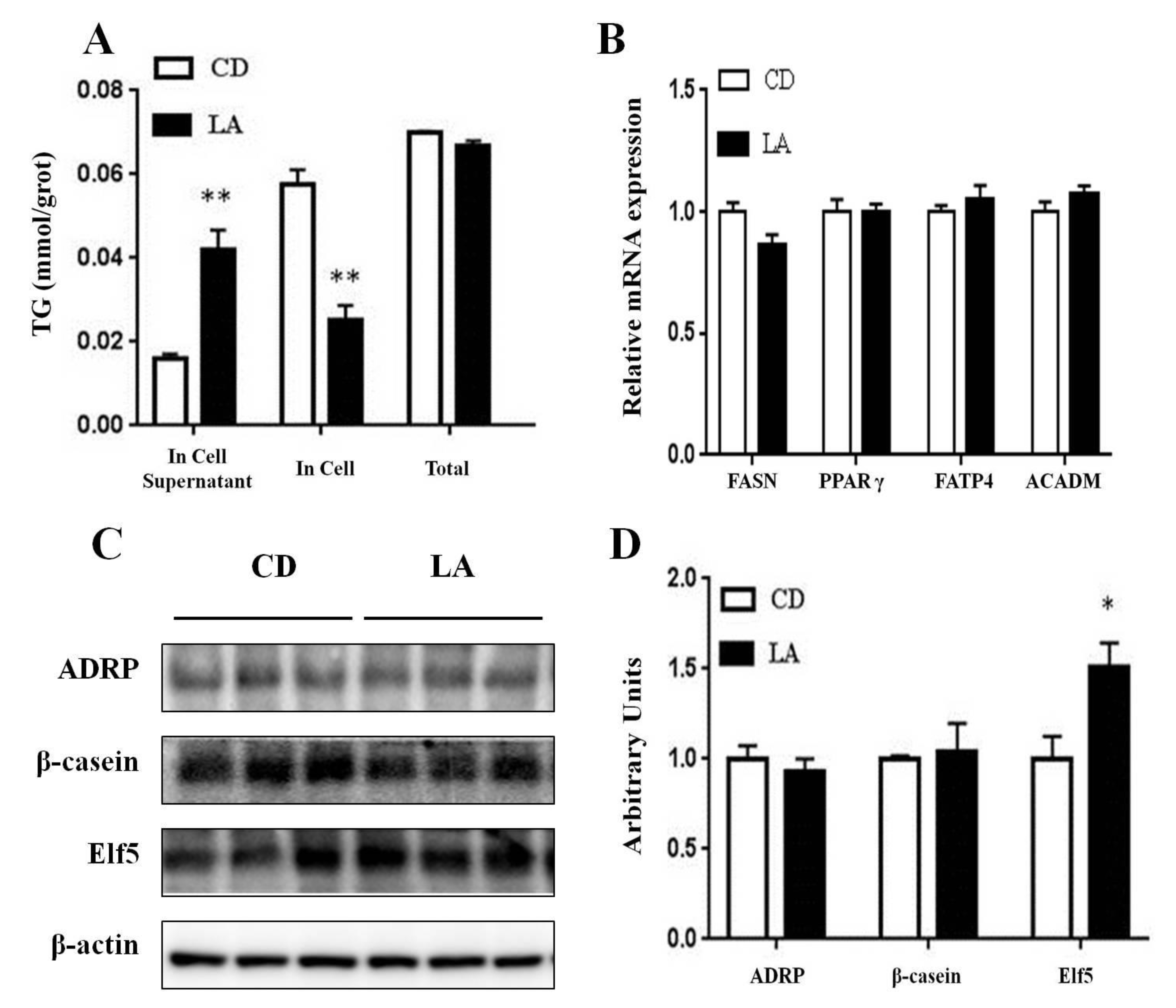
3.4. TG Contents in Mammary Epithelial Cells and Cell Supernatants
3.5. Gene and Protein Expression Related to Milk-fat Synthesis
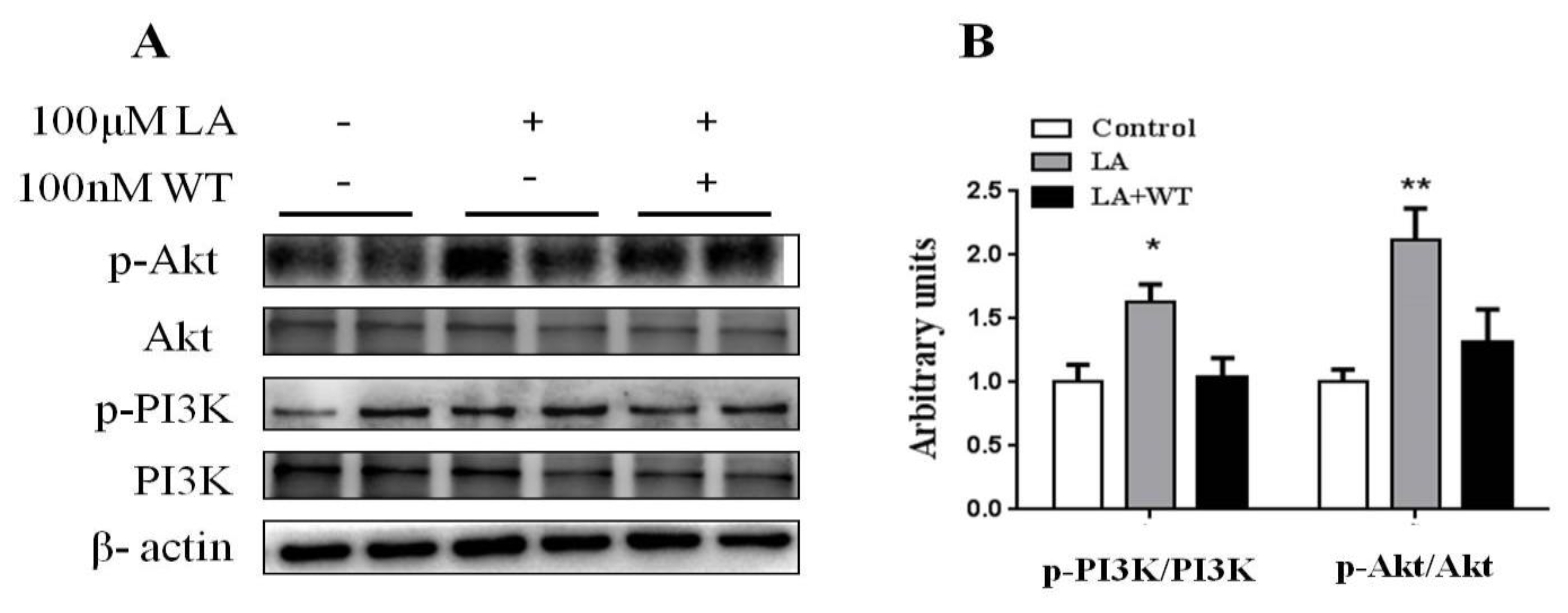
3.6. Inhibition of PI3K/Akt Blocked the Promotion of Differentiated HC11 Development by LA
3.7. Screening of Differential Lipid Metabolites in Serum of LA-Treated Lactating Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oftedal, O.T. The mammary gland and its origin during synapsid evolution. J. Mammary Gland Biol. Neoplasia 2002, 7, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Macias, H.; Hinck, L. Mammary gland development. WIREs Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei, R.; Wu, Z.L.; Hou, Y.Q.; Bazer, F.W.; Wu, G.Y. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musumeci, G.; Castrogiovanni, P.; Szychlinska, M.A.; Aiello, F.C.; Vecchio, G.M.; Salvatorelli, L.; Magro, G.; Imbesi, R. Mammary gland: From embryogenesis to adult life. Acta Histochem. 2015, 117, 379–385. [Google Scholar] [CrossRef]
- Watson, C.J.; Kreuzaler, P.A. Remodeling mechanisms of the mammary gland during involution. Int. J. Dev. Biol. 2011, 55, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Cowley, F.C.; Barber, D.G.; Houlihan, A.V.; Poppi, D.P. Immediate and residual effects of heat stress and restricted intake on milk protein and casein composition and energy metabolism. J. Dairy Sci. 2015, 98, 2356–2368. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G.E.; Tao, S.; Monteiro, A.P.A. Effects of late-gestation heat stress on immunity and performance of calves. J. Dairy Sci. 2016, 99, 3193–3198. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, Y.D.; Zheng, N.; Cheng, J.B.; Wang, J.Q. The effect of heat stress on gene expression and synthesis of heat-shock and milk proteins in bovine mammary epithelial cells. Anim. Sci. J. 2016, 87, 84–91. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, J.Y.; Yuan, C.; Fu, Q.; Zhang, F.L.; Zhu, X.T.; Wang, L.N.; Gao, P.; Shu, G.; Jiang, Q.Y.; et al. Exogenous H2S exerts biphasic effects on porcine mammary epithelial cells proliferation through PI3K/Akt-mTOR signaling pathway. J. Cell. Physiol. 2018, 233, 7071–7081. [Google Scholar] [CrossRef]
- Atwood, C.S.; Hovey, R.C.; Glover, J.P.; Chepko, G.; Ginsburg, E.; Robsin, W.G., Jr.; Vonderhaar, B.K. Progesterone induces side-branching of the ductal epithelium in the mammary glands of peripubertal mice. J. Endocrinol. 2000, 167, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.X.; Manka, D.; Wagner, K.U.; Khan, S.A. Estrogen receptor-α expression in the mammary epithelium is required for ductal and alveolar morphogenesis in mice. Proc. Natl. Acad. Sci. USA 2007, 104, 14718–14723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinberg, D.L.; Barcellos-Hoff, M.H. The pivotal role of Insulin-Like growth factor I in normal mammary development. Endocrinol. Metab. Clin. N. Am. 2011, 40, 461–471. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, K.A.; Shea, M.P.; Salituro, S.; Blohm, C.E.; Schuler, L.A. Prolactin alters the mammary epithelial hierarchy, increasing progenitors and facilitating ovarian steroid action. Stem Cell Rep. 2017, 9, 1167–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.Y.; Zhang, J.; Yuan, C.; Zhang, F.L.; Fu, Q.; Su, H.; Zhu, X.T.; Wang, L.N.; Gao, P.; Shu, G.; et al. Oleic acid stimulates HC11 mammary epithelial cells proliferation and mammary gland development in peripubertal mice through activation of CD36-Ca2+ and PI3K/Akt signaling pathway. Oncotarget 2018, 9, 12982–12994. [Google Scholar] [CrossRef]
- Rodriguez-Cruz, M.; Sánchez, R.; Sánchez, A.M.; Kelleher, S.L.; Sánchez-Muñoz, F.; Maldonado, J.; López-Alarcón, M. Participation of mammary gland in long-chain polyunsaturated fatty acid synthesis during pregnancy and lactation in rats. BBA Mol. Cell Biol. Lipids 2011, 1811, 284–293. [Google Scholar] [CrossRef]
- Tygesen, M.P.; Nielsen, M.O.; NØrgaard, P.; Ranvig, H.; Harrison, A.P.; Tauson, A.H. Late gestational nutrient restriction: Effects on ewes’ metabolic and homeorhetic adaptation, consequences for lamb birth weight and lactation performance. Arch. Anim. Nutr. 2008, 62, 44–59. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Fehrmann-Cartes, K.; Iñiguez-González, G.; Toro-Mujica, P.; Garnsworthy, P.C. Chemical composition, fatty acid composition, and sensory characteristics of Chanco cheese from dairy cows supplemented with soybean and hydrogenated vegetable oils. J. Dairy Sci. 2015, 98, 111–117. [Google Scholar] [CrossRef]
- Meng, Y.Y.; Zhang, J.; Zhang, F.L.; Ai, W.; Zhu, X.T.; Shu, G.; Wang, L.N.; Gao, P.; Xi, Q.Y.; Zhang, Y.L.; et al. Lauric acid stimulates mammary gland development of pubertal mice through activation of GPR84 and PI3K/Akt signaling pathway. J. Agric. Food Chem. 2017, 65, 95–103. [Google Scholar] [CrossRef]
- Dayrit, F.M. The properties of lauric acid and their significance in coconut oil. J. Am. Oil Chem. Soc. 2015, 92, 1–15. [Google Scholar] [CrossRef]
- Bragdon, J.H.; Karmen, A. The fatty acid composition of chylomicrons of chyle and serum following the ingestion of different oils. J. Lipid Res. 1960, 1, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Ong, M.H.L.; Wong, H.K.; Tengku-Muhammad, T.; Choo, Q.C.; Chew, C.H. Pro-atherogenic proteoglycanase ADAMTS-1 is down-regulated by lauric acid through PI3K and JNK signaling pathways in THP-1 derived macrophages. Mol. Biol. Rep. 2019, 46, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B.; Wang, G.Q.; Zhang, M.Y.; Zhuang, L.; Wang, X.J.; Xu, J.R.; Wang, L.N.; Zhu, X.T.; Gao, P.; Xi, Q.Y.; et al. The dipeptide Pro-Asp promotes IGF-1 secretion and expression in hepatocytes by enhancing JAK2/STAT5 signaling pathway. Mol. Cell. Endocrinol. 2016, 436, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B.; Zhou, G.X.; Shu, G.; Wang, L.N.; Zhu, X.T.; Gao, P.; Xi, Q.Y.; Zhang, Y.L.; Yuan, L.; Jiang, Q.Y. Glucose utilization, lipid metabolism and BMP-Smad signaling pathway of porcine intramuscular preadipocytes compared with subcutaneous preadipocytes. Cell. Physiol. Biochem. 2013, 31, 981–996. [Google Scholar] [CrossRef] [PubMed]
- Chin, S.F.; Storkson, J.M.; Albright, K.J.; Cook, M.E.; Pariza, M.W. Conjugated linoleic acid is a growth factor for rats as shown by enhanced weight gain and improved feed efficiency. J. Nutr. 1994, 124, 2344–2349. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.K.B.; Queiroga, R.C.R.E.; Bomfim, M.A.D.; Pessoa, D.C.N.P.; Barbosa, E.A.; Souza, D.L.; Cabral-Filho, J.E.; Medeiros, M.C. Acceleration of reflex maturation and physical development in suckling rats: Effects of a maternal diet containing lipids from goat milk. Nutr. Neurosci. 2014, 17, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bernard, L.; Toral, P.G.; Chilliard, Y. Comparison of mammary lipid metabolism in dairy cows and goats fed diets supplemented with starch, plant oil, or fish oil. J. Dairy Sci. 2017, 100, 9338–9351. [Google Scholar] [CrossRef] [Green Version]
- Oakes, S.R.; Naylor, M.J.; Asselin-Labat, M.; Blazek, K.D.; Gardiner-Garden, M.; Hilton, H.N.; Kazlauskas, M.; Pritchard, M.A.; Chodosh, L.A.; Pfeffer, P.L.; et al. The Ets transcription factor Elf5 specifies mammary alveolar cell fate. Gene Dev. 2008, 22, 581–586. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chehab, R.; Tkalcevic, J.; Naylor, M.J.; Harris, J.; Wilson, T.J.; Tsao, S.; Tellis, I.; Zavarsek, S.; Xu, D.K.; et al. Elf5 is essential for early embryogenesis and mammary gland development during pregnancy and lactation. EMBO J. 2005, 24, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Prizant, R.L.; Barash, I. Negative effects of the amino acids Lys, His, and Thr on S6K1 phosphorylation in mamary epithelial cells. J. Cell. Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef]
- Appuhamy, J.A.; Knoebel, N.A.; Nayananjalie, W.A.; Escobar, J.; Haniqan, M.D. Isoleucine and leucine independently regulate mTOR signaling and protein synthesis in MAC-T cells and bovine mamary tissue slices. J. Nutr. 2012, 142, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.T.; Zhao, F.Q.; Liu, J.X.; Liu, H.Y. Seryl-tRNA synthetase is involved in methionine stimulation of β-casein synthesis in bovine mammary epithelial cells. Br. J. Nutr. 2019, 123, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, Y.; Bar-Peled, L.; Zoncu, R.; Markhard, A.L.; Nada, S.; Sabatini, D.M. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 2010, 141, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Mol. Cell 2010, 40, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, T.D.; Palmer, C.A.; Orlicky, D.J.; Fischer, A.; Rudolph, M.C.; Neville, M.C.; McManaman, J.L. Cytoplasmic lipid droplet accumulation in developing mammary epithelial cells: Roles of adipophilin and lipid metabolism. J. Lipid Res. 2007, 48, 1463–1475. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.B.; Yu, K.; Luo, J.; Li, J.; Tian, H.B.; Zhu, J.J.; Sun, Y.T.; Yao, D.W.; Xu, H.F.; Shi, H.P.; et al. Adipocyte differentiation-related protein promotes lipid accumulation in goat mammary epithelial cells. J. Dairy Sci. 2015, 98, 6954–6964. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.J.; Kollmeyer, J.; Symolon, H.; Momin, A.; Munter, E.; Wang, E.; Kelly, S.; Allegood, J.C.; Liu, Y.; Peng, Q.; et al. Ceramides and other bioactive sphingolipid backbones in health and disease: Lipidomic analysis, metabolism and roles in membrane structure, dynamics, signaling and autophagy. BBA Biomembr. 2006, 1758, 1864–1884. [Google Scholar] [CrossRef] [Green Version]
- Mousley, C.J.; Tyeryar, K.; Ile, K.E.; Schaaf, G.; Brost, R.L.; Boone, C.; Guan, X.L.; Wenk, M.R.; Bankaitis, V.A. Trans-Golgi network and endosome dynamics connect ceramide homeostasis with regulation of the unfolded protein response and TOR signaling in yeast. Mol. Biol. Cell 2008, 19, 4785–4803. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Chen, X.; Zhong, G.; Chen, Q.; Hu, C. Mitofusin 2 downregulation triggers pulmonary artery smooth muscle cell proliferation and apoptosis imbalance in rats with hypoxic pulmonary hypertension via the PI3K/Akt and mitochondrial apoptosis pathways. J. Cardiovasc. Pharmacol. 2016, 67, 164–174. [Google Scholar] [CrossRef]
- Zhang, E.; Gao, B.; Yang, L.; Wu, X.; Wang, Z. Notoginsenoside Ft1 promotes fibroblast proliferation via PI3K/Akt/mTOR signaling pathway and benefits wound healing in genetically diabetic mice. J. Pharmacol. Exp. Ther. 2016, 356, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bai, M.; Ning, C.; Xie, B.; Zhang, J.; Liao, H.; Xiong, J.; Tao, X.; Yan, D.; Xi, X.; et al. Gankyrin facilitates follicle-stimulating hormone-driven ovarian cancer cell proliferation through the PI3K/ AKT/HIF-1α/cyclin D1 pathway. Oncogene 2016, 35, 2506–2517. [Google Scholar] [CrossRef]
- Wickenden, J.A.; Watson, C.J. Key signaling nodes in mammary gland development and cancer. Signaling downstream of PI3 kinase in mammary epithelium: A play in 3 Akts. Breast Cancer Res. 2010, 12, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.W.; Wehde, B.L.; Sakamoto, K.; Triplett, A.A.; Anderson, S.M.; Tsichlis, P.N.; Leone, G.; Wagner, K.U. Stat5 regulates the phosphatidylinositol 3-kinase/Akt1 pathway during mammary gland development and tumorigenesis. Mol. Cell Biol. 2014, 34, 1363–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.H.; Liu, X.; Wang, Z.H.; Lin, X.Y.; Yan, Z.G.; Cao, Q.Q.; Zhao, M.; Shi, K.R. MEN1/Menin regulates milk protein synthesis through mTOR signaling in mammary epithelial cells. Sci. Rep. 2017, 7, 5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.C.; Stairs, D.B.; Boxer, R.B.; Belka, G.K.; Horseman, N.D.; Alvarez, J.V.; Chodosh, L.A. Autocrine prolactin induced by the Pten-Akt pathway is required for lactation initiation and provides a direct link between the Akt and Stat5 pathways. Genes Dev. 2012, 26, 2154–2168. [Google Scholar] [CrossRef] [Green Version]
- Belugin, S.; Diogenes, A.R.; Patil, M.J.; Ginsburg, E.; Henry, M.A.; Akopian, A.N. Mechanisms of transient signaling via short and long prolactin receptor isoforms in female and male sensory neurons. J. Biol. Chem. 2013, 288, 34943–34955. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | gm | kcal |
|---|---|---|
| Casein, 30 Mesh | 200 | 800 |
| L-Cystine | 3 | 12 |
| Corn Starch | 397.5 | 1590 |
| Maltodextrin | 132 | 528 |
| Sucrose | 100 | 400 |
| Cellulose | 50 | 0 |
| Soybean Oil | 70 | 630 |
| Mineral Mix #210025 | 35 | 0 |
| Vitamin Mix #310025 | 10 | 40 |
| Choline Bitartrate | 2.5 | 0 |
| Total | 1000 | 4000 |
| gm% | kcal% | |
| Protein | 18.7 | 19.3 |
| Carbohydrate | 64.7 | 64 |
| Fat | 7 | 16.7 |
| kcal/gm | 4 | |
| Gene | Primer Sequences (5′–3′) | Amplification Length |
|---|---|---|
| ACADM | F: GGCCAGAAGATGTGGATAAC R: GTCGGCTTCCACAATGAA | 123 bp |
| FASN | F: GGAGGTGGTGATAGCCGGTAT R: TGGGTAATCCATAGAGCCCAG | 140 bp |
| FATP4 | F:TCTGTTCTGATTCGTGTTCG R:CAGCATATACCACTACTGG | 137bp |
| PPARγ | F: GGAAGACCACTCGCATTCCTT R: GTAATCAGCAACCATTGGGTCA | 121 bp |
| β-actin | F: GGTCATCACTATTGGCAACGAG R: GAGGTCTTTACGGATGTCAACG | 142 bp |
| Index | Compounds | Class | Type |
|---|---|---|---|
| LIPID-N-0006 | 9,10-EpOME | Eicosanoid | down |
| LIPID-N-0407 | PE (18:2/16:0) | PE (Phosphatidyl ethanolamine) | down |
| LIPID-N-0410 | PE (18:0/18:2) | PE | down |
| LIPID-N-0525 | PE (22:6/16:0) | PE | down |
| LIPID-P-0272 | LPE (0:0/22:1) | LPE (Lysophosphatidyl ethanolamine) | down |
| LIPID-P-0281 | LPE (0:0/20:5) | LPE | down |
| LIPID-P-0528 | PC(O-16:0/14:2) | PC (Phosphatidyl choline) | down |
| LIPID-P-0749 | TG (14:0/16:0/16:0) | TG (Triglyceride) | down |
| LIPID-P-0792 | TG (14:0/18:0/18:1) | TG | down |
| LIPID-P-0804 | TG (18:0/18:0/18:1) | TG | down |
| LIPID-P-0834 | TG (14:1/14:1/16:0) | TG | down |
| LIPID-P-0863 | TG (14:0/18:2/22:0) | TG | down |
| LIPID-P-0894 | TG (14:1/14:1/18:1) | TG | down |
| LIPID-P-0901 | TG (14:0/16:1/18:2) | TG | down |
| LIPID-P-0902 | TG (14:0/16:0/18:3) | TG | down |
| LIPID-P-0969 | TG (14:1/14:1/18:2) | TG | down |
| LIPID-P-1036 | TG (14:0/18:2/18:3) | TG | down |
| LIPID-P-1178 | TG (14:0/20:5/22:4) | TG | down |
| LIPID-P-0716 | SM(d18:0/12:0) | SM (Sphingomyelin) | up |
| LIPID-P-0724 | SM(d18:1/12:0) | SM | up |
| LIPID-P-0833 | TG (12:0/12:0/18:2) | TG | up |
| LIPID-P-0838 | TG (14:0/14:0/18:2) | TG | up |
| LIPID-P-0839 | TG (14:1/16:0/16:1) | TG | up |
| LIPID-P-0896 | TG (12:0/16:0/18:3) | TG | up |
| LIPID-P-0970 | TG (12:0/18:2/18:2) | TG | up |
| LIPID-P-0972 | TG (12:0/18:1/18:3) | TG | up |
| LIPID-P-1031 | TG (14:1/16:1/18:3) | TG | up |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Yang, Q.; Li, F.; Yi, W.; Liu, F.; Wang, S.; Jiang, Q. Effects of Dietary Supplementation of Lauric Acid on Lactation Function, Mammary Gland Development, and Serum Lipid Metabolites in Lactating Mice. Animals 2020, 10, 529. https://doi.org/10.3390/ani10030529
Yang L, Yang Q, Li F, Yi W, Liu F, Wang S, Jiang Q. Effects of Dietary Supplementation of Lauric Acid on Lactation Function, Mammary Gland Development, and Serum Lipid Metabolites in Lactating Mice. Animals. 2020; 10(3):529. https://doi.org/10.3390/ani10030529
Chicago/Turabian StyleYang, Lin, Qiang Yang, Fan Li, Wuzhou Yi, Fangfang Liu, Songbo Wang, and Qingyan Jiang. 2020. "Effects of Dietary Supplementation of Lauric Acid on Lactation Function, Mammary Gland Development, and Serum Lipid Metabolites in Lactating Mice" Animals 10, no. 3: 529. https://doi.org/10.3390/ani10030529