Herbicide Diuron as Endocrine Disrupting Chemicals (EDCs) through Histopathalogical Analysis in Gonads of Javanese Medaka (Oryzias javanicus, Bleeker 1854)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Maintenance of Javanese Medaka
2.2. Chronic Toxicity Testing
2.3. Ethics Statement
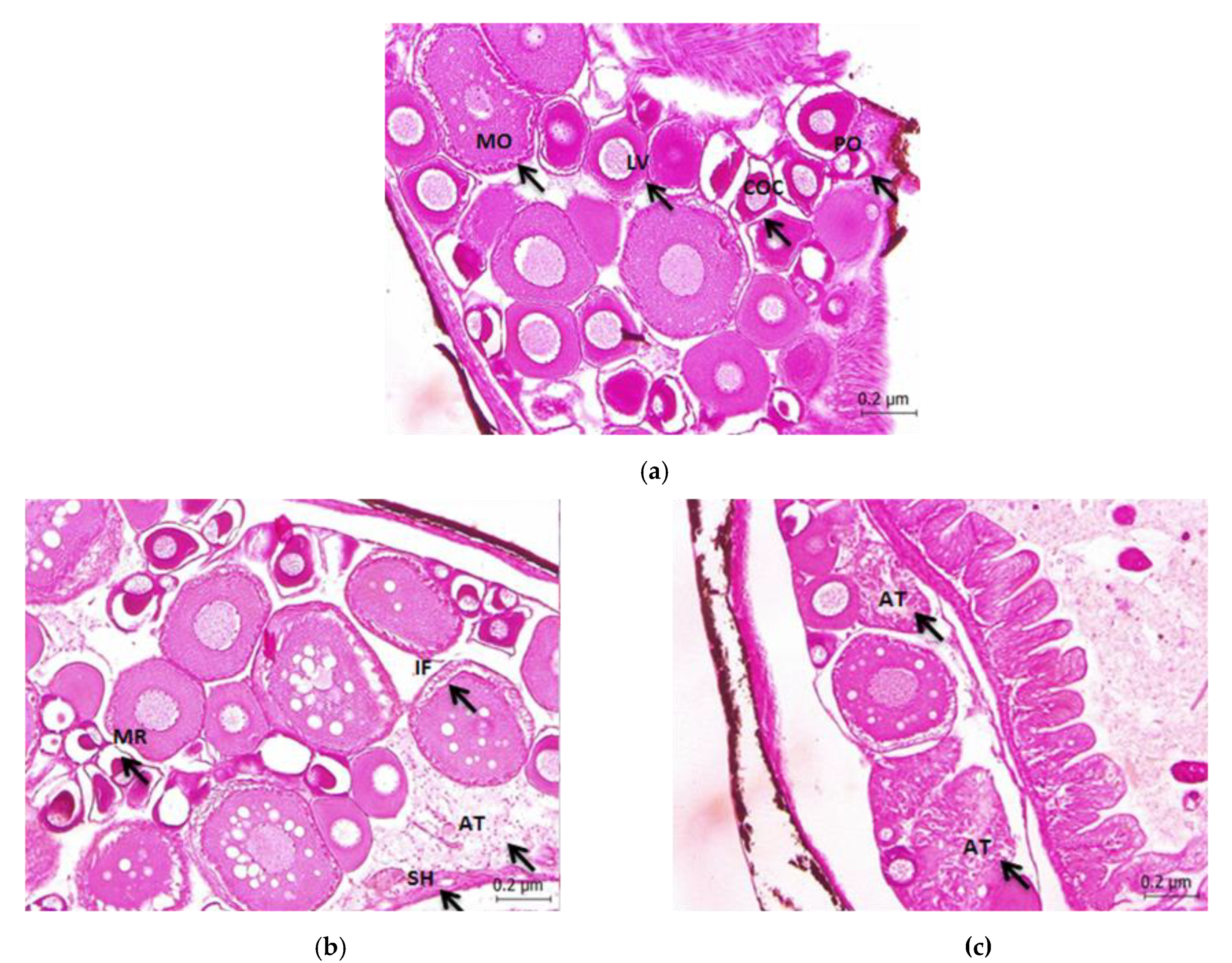
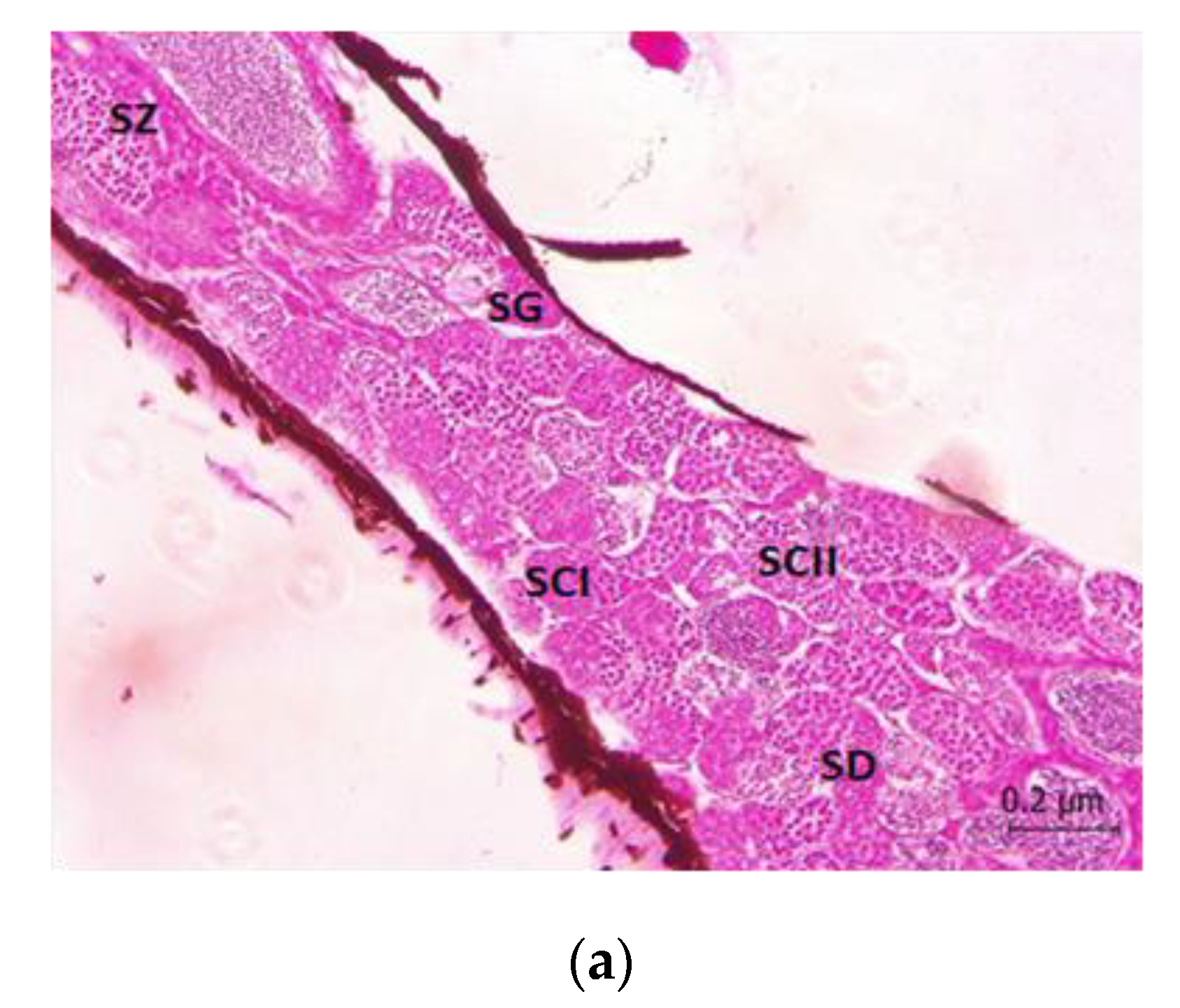
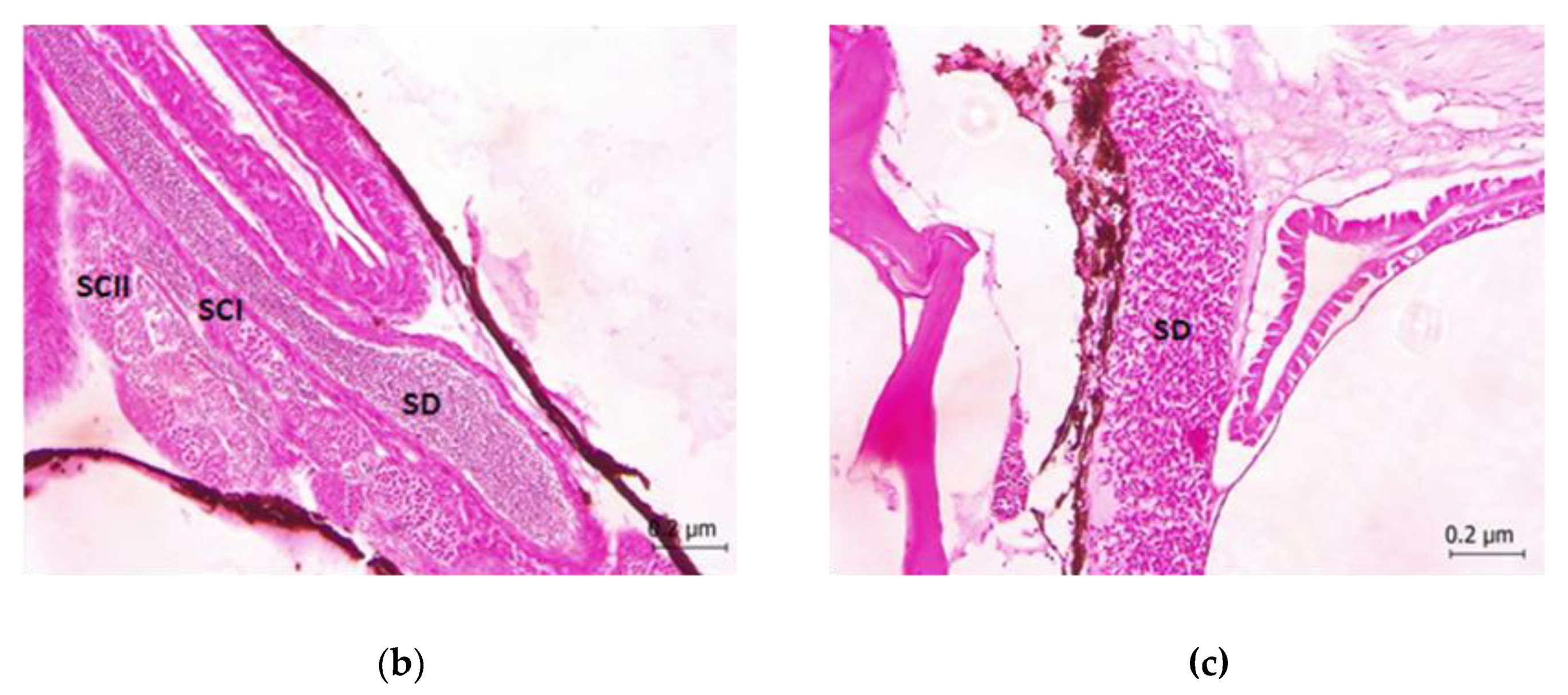
2.4. Histopathological Studies
2.5. Statistical Analysis
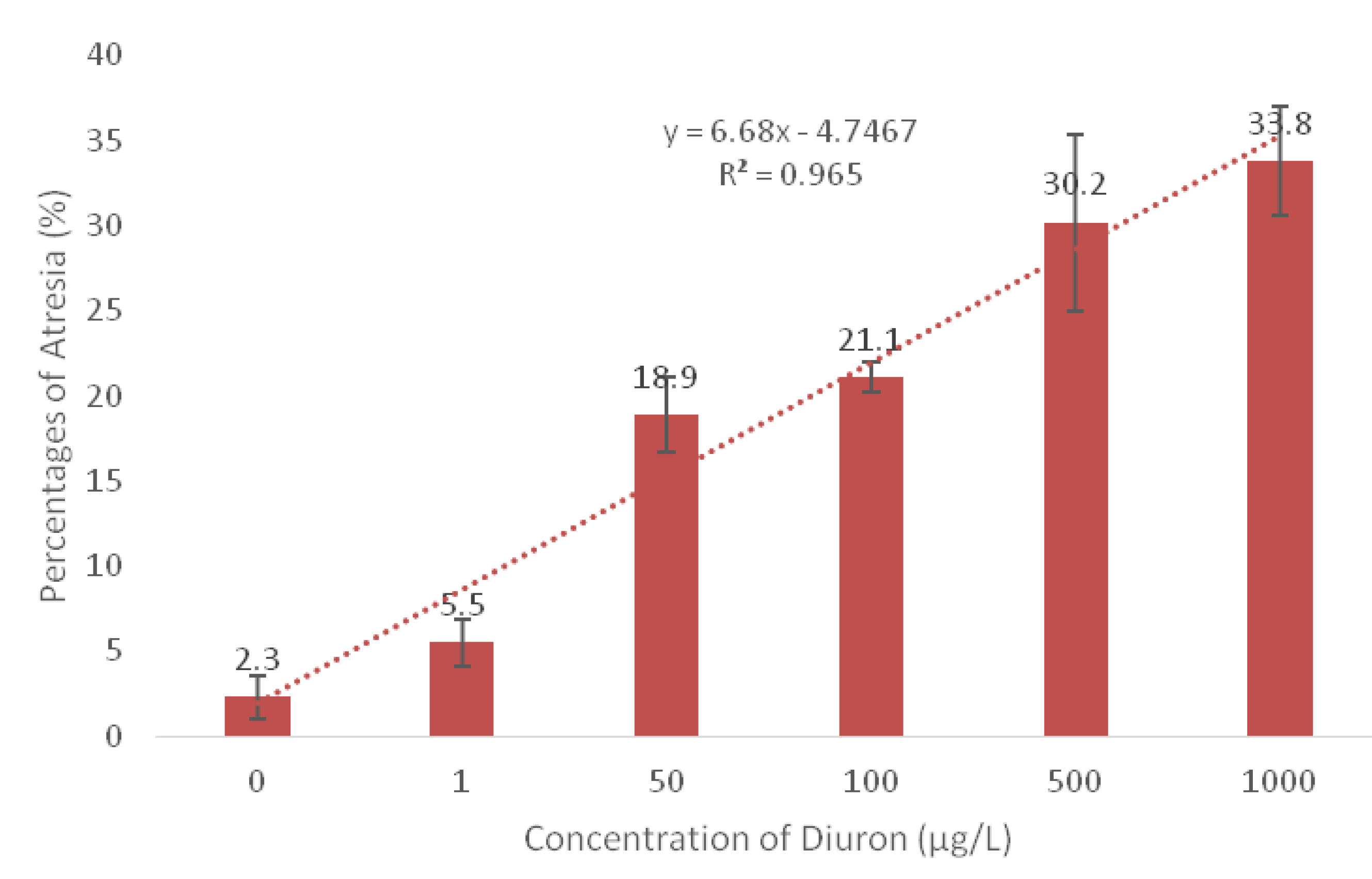
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Giacomazzi, S.; Cochet, N. Environmental impact of Diuron transformation: A review. Chemosphere 2004, 56, 1021–1032. [Google Scholar] [CrossRef]
- Moncada, A. Environmental Fate of Diuron; Environmental Monitoring Branch: Sacramento, CA, USA, 2013.
- Ali, H.R.; Arifin, M.M.; Sheikh, M.A.; Shazili, N.A.M.; Bakari, S.S.; Bachok, Z. Contamination of Diuron in coastal waters around Malaysian Peninsular. Mar. Poll. Bull. 2014, 85, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Ramli, M.I.; Zakaria, Z.; Sahid, I.; Ai, T.Y.; Muhamad, H. Determination of herbicide Diuron levels in palm oil matrices using HPLC-UV. Sains Malaysiana. 2012, 41, 1451–1459. [Google Scholar]
- Muhamad, H.; Ramli, M.I.; Zakaria, Z.; Sahid, I. The fate of Diuron in soil in a Malaysian oil palm plantation. J. Oil Palm Res. 2013, 25, 149–158. [Google Scholar]
- Hanapiah, M.; Zulkifli, S.Z.; Shahrun, M.S.; Mohamat-Yusuff, F.; Ismail, A.; Harino, H.; Inoe, K. Concentration of antifoulant herbicides, Diuron in the vicinity of Port Klang, Malaysia. Isu-isu Terkini Penyelidikan Saintifik Sains Marin di Malaysia 2017, 1, 35–38. [Google Scholar]
- Lamoree, M.H.; Swart, C.P.; Van der Horst, A.; Van Hattum, B. Determination of Diuron and the antifouling paint biocide Irgarol 1051 in Dutch marinas and coastal waters. J. Chromatogr. A. 2002, 970, 183–190. [Google Scholar] [CrossRef]
- Australian Pesticides and Veterinary Medicines Authority. Diuron—The Reconsideration of the Registrations of Selected Products Containing Diuron and Their Associated Labels; APVMA: Sydney, Australia, 2012.
- Lambert, S.J.; Thomas, K.V.; Davy, A.J. Assessment of the risk posed by the antifouling booster biocides Irgarol 1051 and Diuron to freshwater macrophytes. Chemosphere 2006, 63, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.V.; McHugh, M.; Hilton, M.; Waldock, M. Antifouling paint booster biocides in UK coastal waters: Inputs, occurrence and environmental fate. Sci. Total Environ. 2002, 293, 117–127. [Google Scholar] [CrossRef]
- Okamura, H. Photodegradation of the antifouling compounds Irgarol 1051 and Diuron released from a commercial antifouling paint. Chemosphere 2002, 48, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Doval, J.C.; Poquet, M.; Munoz, I. Sublethal effects of the herbicide Diuron on the freshwater snail Physella acuta. Limnetica 2014, 33, 205–216. [Google Scholar]
- Cardone, A.; Comitato, R.; Angelini, F. Spermatogenesis, epididymis morphology and plasma sex steroid secretion in the male lizard Podarcis sicula exposed to Diuron. Environ. Res. 2008, 108, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Saglio, P.; Trijasse, S. Behavioral responses to atrazine and Diuron in goldfish. Arch. Environ. Contam. Toxicol. 1998, 35, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Aziz, F.Z.A.; Zulkifli, S.Z.; Mohamat-Yusuff, F.; Azmai, M.N.A.; Ismail, A. A histological study on mercury-induced gonadal impairment in Javanese medaka (Oryzias javanicus). Turk. J. Fish. Aquat. Sci. 2017, 17, 621–627. [Google Scholar] [CrossRef]
- Kamarudin, N.A.; Zulkifli, S.Z.; Aziz, F.Z.A.; Ismail, A. Histological Alterations in Liver and Kidney of Javanese Medaka (Oryzias javanicus, Bleeker 1854) Exposed to Sublethal Concentration of Herbicide Diuron. Pertanika J. Sci. Technol. 2019, 27, 1041–1050. [Google Scholar]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Shaala, N.M.A.; Zulkifli, S.Z.; Ismail, A.; Azmai, M.N.A.; Mohamat-Yusuff, F. Lethal concentration 50 (LC50) and effects of Diuron on morphology of brine shrimp Artemia salina (Branchiopoda: Anostraca) Nauplii. Procedia Environ. Sci. 2015, 30, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Hasenbein, S.; Peralta, J.; Lawler, S.P.; Connon, R.E. Environmentally relevant concentrations of herbicides impact non-target species at multiple sublethal endpoints. Sci. Total Environ. 2017, 607, 733–743. [Google Scholar] [CrossRef]
- Johnson, R.; Wolf, J.; Braunbeck, T. OECD Guidance Document for the Diagnosis of Endocrine-Related Histopathology of Fish Gonads; Organization for Economic Co-operation and Development: Paris, France, 7 January 2009; pp. 1–96. [Google Scholar]
- Koç, N.D.; Yener, A.; Yüce, R. Ovary maturatıon stages and histological investigation of ovary of the Zebrafish (Danio rerio). Braz. Arch. Biol. Technol. 2008, 51, 513–522. [Google Scholar] [CrossRef]
- Korkmaz, N.; Cengiz, E.I.; Unlu, E.; Uysal, E.; Yanar, M. Cypermethrin-induced histopathological and biochemical changes in Nile tilapia (Oreochromis niloticus), and the protective and recuperative effect of ascorbic acid. Environ. Toxicol. Pharmacol. 2009, 28, 198–205. [Google Scholar] [CrossRef]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol. 2008, 59, 2–11. [Google Scholar] [CrossRef]
- Guraya, S.S. The Cell and Molecular Biology of Fish Oogenesis; Karger Medical and Scientific Publishers: Basel, Switzerland, 1986. [Google Scholar]
- Pereira, T.S.B.; Boscolo, C.N.P.; Felício, A.A.; Batlouni, S.R.; Schlenk, D.; de Almeida, E.A. Estrogenic activities of Diuron metabolites in female Nile tilapia (Oreochromis niloticus). Chemosphere 2016, 146, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscolo, C.N.P.; Pereira, T.S.B.; Batalhão, I.G.; Dourado, P.L.R.; Schlenk, D.; de Almeida, E.A. Diuron metabolites act as endocrine disruptors and alter aggressive behavior in Nile tilapia (Oreochromis niloticus). Chemosphere 2018, 191, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnevali, O.; Santangeli, S.; Forner-Piquer, I.; Basili, D.; Maradonna, F. Endocrine-disrupting chemicals in aquatic environment: What are the risks for fish gametes? Fish Physiol. Biochem. 2018, 44, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- McHugh, K.J.; Smit, N.J.; Van Vuren, J.H.J.; van Dyk, J.C.; Bervoets, L.; Covaci, A.; Wepener, V. A histology-based fish health assessment of the tigerfish, Hydrocynus vittatus from a DDT-affected area. Phys. Chem. Earth. 2011, 36, 895–904. [Google Scholar] [CrossRef]
- Papoulias, D.M.; Tillitt, D.E.; Talykina, M.G.; Whyte, J.J.; Richter, C.A. Atrazine reduces reproduction in Japanese medaka (Oryzias latipes). Aquat. Toxicol. 2014, 154, 230–239. [Google Scholar] [CrossRef]
- Boscolo, C.N.P.; Morais, R.N.; Gonçalves-de-Freitas, E. Same-sized fish groups increase aggressive interaction of sex-reversed males Nile tilapia GIFT strain. Appl. Anim. Behav. Sci. 2011, 135, 154–159. [Google Scholar] [CrossRef]
- Tillitt, D.E.; Papoulias, D.M.; Whyte, J.J.; Richter, C.A. Atrazine reduces reproduction in fathead minnow (Pimephales promelas). Aquat. Toxicol. 2010, 99, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Marlatt, V.L.; Lo, B.P.; Ornostay, A.; Hogan, N.S.; Kennedy, C.J.; Elphick, J.R.; Martyniuk, C.J. The effects of the urea-based herbicide linuron on reproductive endpoints in the fathead minnow (Pimephales promelas). Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2013, 157, 24–32. [Google Scholar] [CrossRef]
- Pereira, T.S.B.; Boscolo, C.N.P.; da Silva, D.G.H.; Batlouni, S.R.; Schlenk, D.; de Almeida, E.A. Anti-androgenic activities of Diuron and its metabolites in male Nile tilapia (Oreochromis niloticus). Aquat. Toxicol. 2015, 164, 10–15. [Google Scholar] [CrossRef]
- Bauer, E.R.; Meyer, H.H.; Sauerwein, H.; Stahlschmidt-Allner, P. Application of an androgen receptor assay for the characterisation of the androgenic or antiandrogenic activity of various phenylurea herbicides and their derivatives. Anal. 1998, 123, 2485–2487. [Google Scholar] [CrossRef]
- Felício, A.A.; Crago, J.; Maryoung, L.A.; Almeida, E.A.; Schlenk, D. Effects of alkylphenols on the biotransformation of Diuron and enzymes involved in the synthesis and clearance of sex steroids in juvenile male tilapia (Oreochromus mossambica). Aquat. Toxicol. 2016, 180, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Thibaut, R.; Porte, C. Effects of endocrine disrupters on sex steroid synthesis and metabolism pathways in fish. J. Steroid Biochem. Mol. Biol. 2004, 92, 485–494. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration Treated (µg/L) | Increases in Oocyte Atresia (%) | Gonadal Staging Median Stage |
|---|---|---|
| Control | 2.0 | 4 a |
| 1 | 5.5 | 4 a |
| 50 | 18.9 | 3 b |
| 100 | 21.1 | 2 ab |
| 500 | 30.2 | 2 ab |
| 1000 | 33.8 | 2 ab |
| Concentration Treated (µg/L) | Gonadal Staging Median Stage |
|---|---|
| Control | 5 a |
| 1 | 5 a |
| 50 | 3 b |
| 100 | 3 b |
| 500 | 1 c |
| 1000 | 1 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamarudin, N.A.; Zulkifli, S.Z.; Azmai, M.N.A.; Abdul Aziz, F.Z.; Ismail, A. Herbicide Diuron as Endocrine Disrupting Chemicals (EDCs) through Histopathalogical Analysis in Gonads of Javanese Medaka (Oryzias javanicus, Bleeker 1854). Animals 2020, 10, 525. https://doi.org/10.3390/ani10030525
Kamarudin NA, Zulkifli SZ, Azmai MNA, Abdul Aziz FZ, Ismail A. Herbicide Diuron as Endocrine Disrupting Chemicals (EDCs) through Histopathalogical Analysis in Gonads of Javanese Medaka (Oryzias javanicus, Bleeker 1854). Animals. 2020; 10(3):525. https://doi.org/10.3390/ani10030525
Chicago/Turabian StyleKamarudin, Nur Amiera, Syaizwan Zahmir Zulkifli, Mohammad Noor Amal Azmai, Fatin Zahidah Abdul Aziz, and Ahmad Ismail. 2020. "Herbicide Diuron as Endocrine Disrupting Chemicals (EDCs) through Histopathalogical Analysis in Gonads of Javanese Medaka (Oryzias javanicus, Bleeker 1854)" Animals 10, no. 3: 525. https://doi.org/10.3390/ani10030525