Nutritional Properties and Oxidative Indices of Broiler Breast Meat Affected by Wooden Breast Abnormality


 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Characterization of Muscle Abnormalities
2.2. Properties of 20-min Postmortem Skeletal Muscle
2.3. Chemical Composition, Oxidation of Lipid and Protein and Technological Properties of 24-h Postmortem Breast Meat
2.4. Absolute Gene Expression
2.5. Statistical Analysis
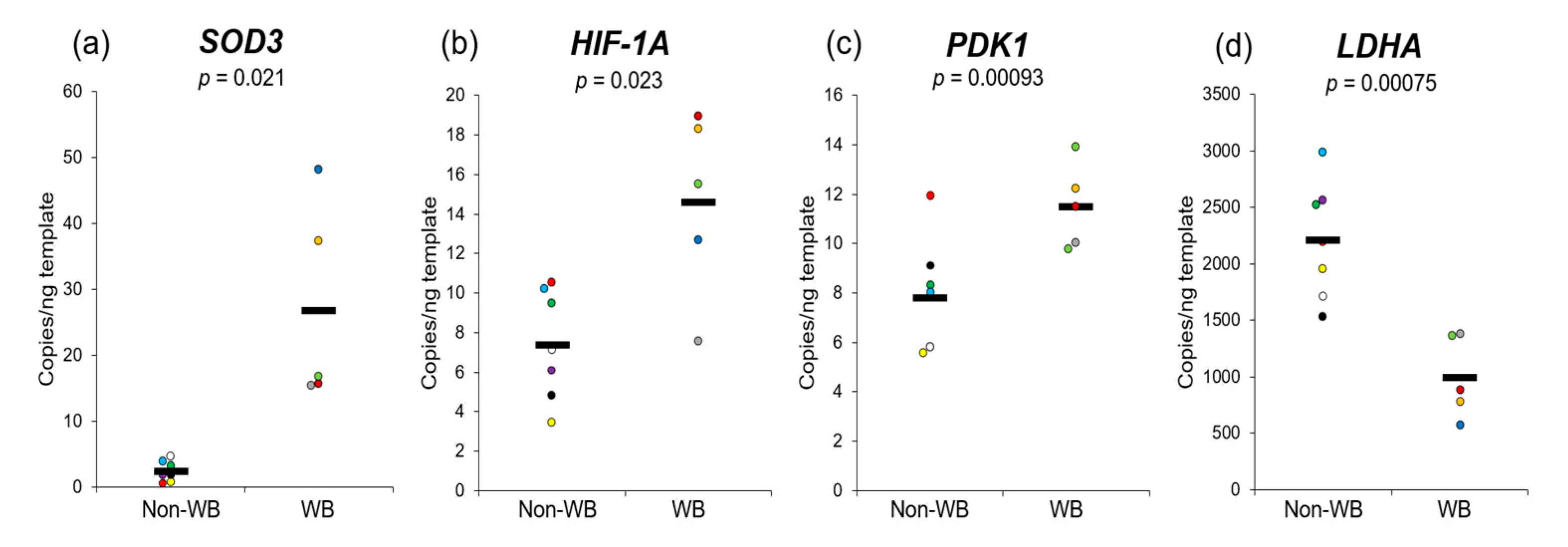
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boye, J.; Wijesinha-Bettoni, R.; Burlingame, B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br. J. Nutr. 2012, 108, S183–S211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrón-Hoyos, J.M.; Archuleta, A.R.; del Refugio Falcón-Villa, M.; Canett-Romero, R.; Cinco-Moroyoqui, F.J.; Romero-Barancini, A.L.; Rueda-Puente, E.O. Protein quality evaluation of animal food proteins by in-vitro methodologies. Food Nutr. Sci. 2013, 04, 376–384. [Google Scholar] [CrossRef] [Green Version]
- Marangoni, F.; Corsello, G.; Cricelli, C.; Ferrara, N.; Ghiselli, A.; Lucchin, L.; Poli, A. Role of poultry meat in a balanced diet aimed at maintaining health and wellbeing: An Italian consensus document. Food Nutr. Res. 2015, 59, 27606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbut, S. Recent myopathies in broiler’s breast meat fillets. Worlds. Poult. Sci. J. 2019, 75, 559–582. [Google Scholar] [CrossRef]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-breast, white striping, and spaghetti meat: Causes, consequences and consumer perception of emerging broiler meat abnormalities. Compr. Rev. Food Sci. Food Saf. 2019, 18, 565–583. [Google Scholar] [CrossRef] [Green Version]
- Sihvo, H.-K.; Immonen, K.; Puolanne, E. Myodegeneration with fibrosis and regeneration in the pectoralis major muscle of broilers. Vet. Pathol. 2014, 51, 619–623. [Google Scholar] [CrossRef]
- Velleman, S.G.; Clark, D.L. Histopathologic and myogenic gene expression changes associated with wooden breast in broiler breast muscles. Avian Dis. 2015, 59, 410–418. [Google Scholar] [CrossRef]
- Soglia, F.; Mudalal, S.; Babini, E.; Di Nunzio, M.; Mazzoni, M.; Sirri, F.; Cavani, C.; Petracci, M. Histology, composition, and quality traits of chicken pectoralis major muscle affected by wooden breast abnormality. Poult. Sci. 2016, 95, 651–659. [Google Scholar] [CrossRef]
- Chatterjee, D.; Zhuang, H.; Bowker, B.C.; Sanchez-Brambila, G.; Rincon, A.M. Instrumental texture characteristics of broiler pectoralis major with the wooden breast condition. Poult. Sci. 2016, 95, 2449–2454. [Google Scholar] [CrossRef]
- Tijare, V.V.; Yang, F.L.; Kuttappan, V.A.; Alvarado, C.Z.; Coon, C.N.; Owens, C.M. Meat quality of broiler breast fillets with white striping and woody breast muscle myopathies. Poult. Sci. 2016, 95, 2167–2173. [Google Scholar] [CrossRef]
- Malila, Y.; U-Chupaj, J.; Srimarut, Y.; Chaiwiwattrakul, P.; Uengwetwanit, T.; Arayamethakorn, S.; Punyapornwithaya, V.; Sansamur, C.; Kirschke, C.P.; Huang, L.; et al. Monitoring of white striping and wooden breast cases and impacts on quality of breast meat collected from commercial broilers (Gallus gallus). Asian-Australasian J. Anim. Sci. 2018, 31, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Dalle Zotte, A.; Ricci, R.; Cullere, M.; Serva, L.; Tenti, S.; Marchesini, G. Research Note: Effect of chicken genotype and white striping–wooden breast condition on breast meat proximate composition and amino acid profile. Poult. Sci. 2020, 99, 1797–1803. [Google Scholar] [CrossRef] [PubMed]
- Madruga, M.S.; da Rocha, T.C.; de Carvalho, L.M.; Sousa, A.M.B.L.; de Sousa Neto, A.C.; Coutinho, D.G.; de Carvalho Ferreira, A.S.; Soares, A.J.; de Sousa Galvão, M.; Ida, E.I.; et al. The impaired quality of chicken affected by the wooden breast myopathy is counteracted in emulsion-type sausages. J. Food Sci. Technol. 2019, 56, 1380–1388. [Google Scholar] [CrossRef]
- Hoving-Bolink, A.H.; Kranen, R.W.; Klont, R.E.; Gerritsen, C.L.M.; de Greef, K.H. Fibre area and capillary supply in broiler breast muscle in relation to productivity and ascites. Meat Sci. 2000, 56, 397–402. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Airas, N.; Lindén, J.; Puolanne, E. Pectoral vessel density and early ultrastructural changes in broiler chicken wooden breast myopathy. J. Comp. Pathol. 2018, 161, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Clanton, T.L. Hypoxia-induced reactive oxygen species formation in skeletal muscle. J. Appl. Physiol. 2007, 102, 2379–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutryn, M.F.; Brannick, E.M.; Fu, W.; Lee, W.R.; Abasht, B. Characterization of a novel chicken muscle disorder through differential gene expression and pathway analysis using RNA-sequencing. BMC Genom. 2015, 16, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambonelli, P.; Zappaterra, M.; Soglia, F.; Petracci, M.; Sirri, F.; Cavani, C.; Davoli, R. Detection of differentially expressed genes in broiler pectoralis major muscle affected by white striping–wooden breast myopathies. Poult. Sci. 2016, 95, 2771–2785. [Google Scholar] [CrossRef] [PubMed]
- Papah, M.B.; Brannick, E.M.; Schmidt, C.J.; Abasht, B. Evidence and role of phlebitis and lipid infiltration in the onset and pathogenesis of wooden breast disease in modern broiler chickens. Avian Pathol. 2017, 46, 623–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malila, Y.; Thanatsang, K.; Arayamethakorn, S.; Uengwetwanit, T.; Srimarut, Y.; Petracci, M.; Strasburg, G.M.; Rungrassamee, W.; Visessanguan, W. Absolute expressions of hypoxia-inducible factor-1 alpha (HIF1A) transcript and the associated genes in chicken skeletal muscle with white striping and wooden breast myopathies. PLoS ONE 2019, 14, e0220904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative stress and metabolic perturbations in wooden breast disorder in chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, Y.; Pan, D.; He, J.; Cao, J.; Wang, H.; Ertbjerg, P. Metabolite profile based on 1H NMR of broiler chicken breasts affected by wooden breast myodegeneration. Food Chem. 2020, 310, 125852. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Ahn, D. Mechanism of lipid peroxidation in meat and meat products -a review. Food Sci. Biotechnol. 2005, 14, 152–163. [Google Scholar]
- Møller, J.K.S.; Skibsted, L.H. Myoglobins—The link between discoloration and lipid oxidation in muscle and meat. Quim. Nova 2006, 29, 1270–1278. [Google Scholar] [CrossRef]
- Ahmed, M.; Pickova, J.; Ahmad, T.; Liaquat, M.; Farid, A.; Jahangir, M. Oxidation of lipids in foods. Sarhad J. Agric. 2016, 32, 230–238. [Google Scholar] [CrossRef]
- Keller, J.; Camaré, C.; Bernis, C.; Astello-García, M.; de la Rosa, A.-P.B.; Rossignol, M.; del Socorro Santos Díaz, M.; Salvayre, R.; Negre-Salvayre, A.; Guéraud, F. Antiatherogenic and antitumoral properties of Opuntia cladodes: Inhibition of low density lipoprotein oxidation by vascular cells, and protection against the cytotoxicity of lipid oxidation product 4-hydroxynonenal in a colorectal cancer cellular model. J. Physiol. Biochem. 2015, 71, 577–587. [Google Scholar] [CrossRef]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estévez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef]
- Bowker, B.C.; Zhuang, H. Relationship between muscle exudate protein composition and broiler breast meat quality. Poult. Sci. 2013, 92, 1385–1392. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Lindén, J.; Airas, N.; Immonen, K.; Valaja, J.; Puolanne, E. Wooden breast myodegeneration of pectoralis major muscle over the growth period in broilers. Vet. Pathol. 2017, 54, 119–128. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Jiménez-Martín, E.; Ruiz, J.; Pérez-Palacios, T.; Silva, A.; Antequera, T. Gas chromatography-mass spectrometry method for the determination of free amino acids as their dimethyl-tert-butylsilyl (TBDMS) derivatives in animal source food. J. Agric. Food Chem. 2012, 60, 2456–2463. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Biomembranes-Part C: Biological Oxidations; Fleischer, S., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 1978; Volume 52, pp. 302–310. [Google Scholar]
- Soglia, F.; Petracci, M.; Ertbjerg, P. Novel DNPH-based method for determination of protein carbonylation in muscle and meat. Food Chem. 2016, 197, 670–675. [Google Scholar] [CrossRef] [PubMed]
- U-chupaj, J.; Malila, Y.; Gamonpilas, C.; Kijroongrojana, K.; Petracci, M.; Benjakul, S.; Visessanguan, W. Differences in textural properties of cooked caponized and broiler chicken breast meat. Poult. Sci. 2017, 96, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Mahy, M.; Van Eycken, L.; Oosterlinck, A. Evaluation of Uniform Color Spaces Developed after the Adoption of CIELAB and CIELUV. Color Res. Appl. 1994, 19, 105–121. [Google Scholar] [CrossRef]
- Cai, K.; Shao, W.; Chen, X.; Campbell, Y.L.; Nair, M.N.; Suman, S.P.; Beach, C.M.; Guyton, M.C.; Schilling, M.W. Meat quality traits and proteome profile of woody broiler breast (pectoralis major) meat. Poult. Sci. 2018, 97, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, P.; Hilton, K.; Weil, J.; Suesuttajit, N.; Beitia, A.; Owens, C.M.; Coon, C. Characterizing woody breast myopathy in a meat broiler line by heat production, microbiota, and plasma metabolites. Front. Vet. Sci. 2020, 6, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velleman, S.G. Recent developments in breast muscle myopathies associated with growth in poultry. Annu. Rev. Anim. Biosci. 2019, 7, 289–308. [Google Scholar] [CrossRef]
- Mudalal, S.; Lorenzi, M.; Soglia, F.; Cavani, C.; Petracci, M. Implications of white striping and wooden breast abnormalities on quality traits of raw and marinated chicken meat. Animal 2015, 9, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Baldi, G.; Soglia, F.; Laghi, L.; Tappi, S.; Rocculi, P.; Tavaniello, S.; Prioriello, D.; Mucci, R.; Maiorano, G.; Petracci, M. Comparison of quality traits among breast meat affected by current muscle abnormalities. Food Res. Int. 2019, 115, 369–376. [Google Scholar] [CrossRef]
- Livingston, M.L.; Ferket, P.R.; Brake, J.; Livingston, K.A. Dietary amino acids under hypoxic conditions exacerbates muscle myopathies including wooden breast and white stripping. Poult. Sci. 2019, 98, 1517–1527. [Google Scholar] [CrossRef]
- Tasoniero, G.; Bowker, B.; Stelzleni, A.; Zhuang, H.; Rigdon, M.; Thippareddi, H. Use of blade tenderization to improve wooden breast meat texture. Poult. Sci. 2019, 98, 4204–4211. [Google Scholar] [CrossRef]
- Berri, C.; Debut, M.; Santé-Lhoutellier, V.; Arnould, C.; Boutten, B.; Sellier, N.; Baéza, E.; Jehl, N.; Jégo, Y.; Duclos, M.J.; et al. Variations in chicken breast meat quality: Implications of struggle and muscle glycogen content at death. Br. Poult. Sci. 2005, 46, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Yen, C.-N.; Daughtry, M.R.; Bodmer, J.; Bowker, B.C.; Zhuang, H.; Petracci, M.; Gerrard, D.E. Exploring the factors contributing to the high ultimate ph of broiler pectoralis major muscles affected by wooden breast condition. Front. Physiol. 2020, 11, 343. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; Gao, F.; Tume, R.K.; Zhou, G.; Xu, X. Stress Effects on meat quality: A mechanistic perspective. Compr. Rev. Food Sci. Food Saf. 2019, 18, 380–401. [Google Scholar] [CrossRef] [Green Version]
- Tasoniero, G.; Cullere, M.; Cecchinato, M.; Puolanne, E.; Dalle Zotte, A. Technological quality, mineral profile, and sensory attributes of broiler chicken breasts affected by white striping and wooden breast myopathies. Poult. Sci. 2016, 95, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Dalle Zotte, A.; Tasoniero, G.; Puolanne, E.; Remignon, H.; Cecchinato, M.; Catelli, E.; Cullere, M. Effect of “wooden breast” appearance on poultry meat quality, histological traits, and lesions characterization. Czech J. Anim. Sci. 2017, 62, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, H.; Bowker, B. The wooden breast condition results in surface discoloration of cooked broiler pectoralis major. Poult. Sci. 2018, 97, 4458–4461. [Google Scholar] [CrossRef]
- Soglia, F.; Silva, A.K.; Lião, L.M.; Laghi, L.; Petracci, M. Effect of broiler breast abnormality and freezing on meat quality and metabolites assessed by 1 H-NMR spectroscopy. Poult. Sci. 2019, 98, 7139–7150. [Google Scholar] [CrossRef]
- Dalgaard, L.B.; Rasmussen, M.K.; Bertram, H.C.; Jensen, J.A.; Møller, H.S.; Aaslyng, M.D.; Hejbøl, E.K.; Pedersen, J.R.; Elsser-Gravesen, D.; Young, J.F. Classification of wooden breast myopathy in chicken pectoralis major by a standardised method and association with conventional quality assessments. Int. J. Food Sci. Technol. 2018, 53, 1744–1752. [Google Scholar] [CrossRef]
- Xing, T.; Zhao, X.; Zhang, L.; Li, J.L.; Zhou, G.H.; Xu, X.L.; Gao, F. Characteristics and incidence of broiler chicken wooden breast meat under commercial conditions in China. Poult. Sci. 2020, 99, 620–628. [Google Scholar] [CrossRef]
- Sanchez-Brambila, G.; Bowker, B.C.; Chatterjee, D.; Zhuang, H. Descriptive texture analyses of broiler breast fillets with the wooden breast condition stored at 4 °C and –20 °C. Poult. Sci. 2018, 97, 1762–1767. [Google Scholar] [CrossRef]
- Dransfield, E.; Sosnicki, A.A. Relationship between muscle growth and poultry meat quality. Poult. Sci. 1999, 78, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Limpisophon, K.; E-tun, S.; Koeipudsa, C.; Charoensuk, D.; Malila, Y. Characterization of breast meat collected from spent lohmann brown layers in comparison to commercial ross broilers. Braz. J. Poult. Sci. 2019, 21, eRBCA-2018-0941. [Google Scholar] [CrossRef]
- Soglia, F.; Gao, J.; Mazzoni, M.; Puolanne, E.; Cavani, C.; Petracci, M.; Ertbjerg, P. Superficial and deep changes of histology, texture and particle size distribution in broiler wooden breast muscle during refrigerated storage. Poult. Sci. 2017, 96, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T. The role of intramuscular connective tissue in meat texture. Anim. Sci. J. 2010, 81, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Iwasaki, T.; Yamada, M.; Yoshida, T.; Watanabe, T. Rapid growth rate results in remarkably hardened breast in broilers during the middle stage of rearing: A biochemical and histopathological study. PLoS ONE 2018, 13, e0193307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papah, M.B.; Brannick, E.M.; Schmidt, C.J.; Abasht, B. Gene expression profiling of the early pathogenesis of wooden breast disease in commercial broiler chickens using RNA-sequencing. PLoS ONE 2018, 13, e0207346. [Google Scholar] [CrossRef]
- Papah, M.B.; Abasht, B. Dysregulation of lipid metabolism and appearance of slow myofiber-specific isoforms accompany the development of wooden breast myopathy in modern broiler chickens. Sci. Rep. 2019, 9, 17170. [Google Scholar] [CrossRef]
- Soglia, F.; Laghi, L.; Canonico, L.; Cavani, C.; Petracci, M. Functional property issues in broiler breast meat related to emerging muscle abnormalities. Food Res. Int. 2016, 89, 1071–1076. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [Green Version]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Cecarini, V.; Gee, J.; Fioretti, E.; Amici, M.; Angeletti, M.; Eleuteri, A.M.; Keller, J.N. Protein oxidation and cellular homeostasis: Emphasis on metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Utrera, M.; Estévez, M. Oxidation of myofibrillar proteins and impaired functionality: Underlying mechanisms of the carbonylation pathway. J. Agric. Food Chem. 2012, 60, 8002–8011. [Google Scholar] [CrossRef] [PubMed]
- Soyer, A.; Özalp, B.; Dalmış, Ü.; Bilgin, V. Effects of freezing temperature and duration of frozen storage on lipid and protein oxidation in chicken meat. Food Chem. 2010, 120, 1025–1030. [Google Scholar] [CrossRef]
- Roldan, M.; Antequera, T.; Armenteros, M.; Ruiz, J. Effect of different temperature–time combinations on lipid and protein oxidation of sous-vide cooked lamb loins. Food Chem. 2014, 149, 129–136. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, M.A.; Schlater, A.E.; Green, T.L.; Kanatous, S.B. In the face of hypoxia: Myoglobin increases in response to hypoxic conditions and lipid supplementation in cultured Weddell seal skeletal muscle cells. J. Exp. Biol. 2012, 215, 806–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, M.; Petracci, M. Benefits of magnesium supplementation to broiler subjected to dietary and heat stress: Improved redox status, breast quality and decreased myopathy incidence. Antioxidants 2019, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, W.B.; Edwards, D.G.; Jurkovitz, C.T.; Weintraub, W.S. Dietary sodium and health: More than just blood pressure. J. Am. Coll. Cardiol. 2015, 65, 1042–1050. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.L.; Santiago, S.; Nunes, M.L. Assessment of the essential element and heavy metal content of edible fish muscle. Anal. Bioanal. Chem. 2005, 382, 426–432. [Google Scholar] [CrossRef]
- Jastrzębska, A.; Cichosz, M.; Szłyk, E. Simple and rapid determination of phosphorus in meat samples by WD-XRF method. J. Anal. Chem. 2010, 65, 376–381. [Google Scholar] [CrossRef]
- Ali, M.; Choudhury, T.R.; Hossain, B.; Ali, M.P. Determination of traces of molybdenum and lead in foods by x-ray fluorescence spectrometry. Springerplus 2014, 3, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turhan, S. Aluminium contents in baked meats wrapped in aluminium foil. Meat Sci. 2006, 74, 644–647. [Google Scholar] [CrossRef]
- Griffin, J.R.; Moraes, L.; Wick, M.; Lilburn, M.S. Onset of white striping and progression into wooden breast as defined by myopathic changes underlying pectoralis major growth. Estimation of growth parameters as predictors for stage of myopathy progression. Avian Pathol. 2018, 47, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Tchernyshyov, I.; Semenza, G.L.; Dang, C. V HIF-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuttappan, V.A.; Bottje, W.; Ramnathan, R.; Hartson, S.D.; Coon, C.N.; Kong, B.-W.; Owens, C.M.; Vazquez-Añon, M.; Hargis, B.M. Proteomic analysis reveals changes in carbohydrate and protein metabolism associated with broiler breast myopathy. Poult. Sci. 2017, 96, 2992–2999. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Kogut, M.H.; Genovese, K.J.; Hsu, C.-Y.; Lee, J.T.; Farnell, Y.Z. Altered expression of lactate dehydrogenase and monocarboxylate transporter involved in lactate metabolism in broiler wooden breast. Poult. Sci. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Lake, J.A.; Abasht, B. Glucolipotoxicity: A proposed etiology for wooden breast and related myopathies in commercial broiler chickens. Front. Physiol. 2020, 11, 169. [Google Scholar] [CrossRef]
- Salles, G.B.C.; Boiago, M.M.; Silva, A.D.; Morsch, V.M.; Gris, A.; Mendes, R.E.; Baldissera, M.D.; da Silva, A.S. Lipid peroxidation and protein oxidation in broiler breast fillets with white striping myopathy. J. Food Biochem. 2019, 43, e12792. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Delgado, J.; Madruga, M.S.; Estévez, M. Pinpointing oxidative stress behind the white striping myopathy: Depletion of antioxidant defenses, accretion of oxidized proteins and impaired proteostasis. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef]


{kind=link}
| NCBI Accession | Gene ID | Sequence (5’→3’) | Amplicon Size (bp) | Template 1 (ng) |
|---|---|---|---|---|
| XR_001466725.2 | HIF-1A | F: ATCAGAGTGGTTGTCCAGCAG R: CAGTCCAAGCCCACCTTACT | 111 | 25 |
| NM_205284.1 | LDHA | F: TTCTCTGCCAGCTGAATAGCTT R: CGGGTCATTGTCTTGTTGCAT | 200 | 1 |
| NM_001031352.3 | PDK1 | F: TGCCAAGCAGTGAGCCAAAG R: AACTCCCTTCACATGACACACAT | 97 | 10 |
| XM_015285700.1 | SOD3 | F: TACAAACCCAACCTCTTCGC R: GTTATTGCCCTTGCCCATGT | 102 | 10 |
| Composition | Non-WB | WB | p-Value |
|---|---|---|---|
| pH20 | 6.86 ± 0.04 | 6.71 ± 0.03 | 0.012 |
| Glycogen (mg/g muscle) | 2.41 ± 0.40 | 1.90 ± 0.39 | 0.332 |
| Lactate (mg/g muscle) | 2.55 ± 0.14 | 2.81 ± 0.21 | 0.217 |
| Composition | Non-WB | WB | p-Value |
|---|---|---|---|
| Moisture (%) | 74.76 ± 0.55 | 77.86 ± 0.60 | 0.004 |
| Protein (% wet basis) | 22.12 ± 0.22 | 18.67 ± 0.57 | 0.002 |
| Fat (% wet basis) | 1.62 ± 0.05 | 1.52 ± 0.19 | 0.667 |
| Protein (% dry basis) | 87.92 ± 2.27 | 84.38 ± 1.49 | 0.222 |
| Fat (% dry basis) | 6.41 ± 0.21 | 6.81 ± 0.66 | 0.583 |
| Ash (% wet basis) | 1.16 ± 0.03 | 1.05 ± 0.02 | 0.012 |
| Lactate (mg/g meat) | 7.87 ± 0.51 | 6.18 ± 0.35 | 0.022 |
| TBARS (μmol malondialdehyde/kg meat) | 1.15 ± 0.11 | 1.23 ± 0.06 | 0.593 |
| Protein carbonyls (nmol/mg protein) | 0.79 ± 0.05 | 1.09 ± 0.10 | 0.029 |
| Amino Acid | Non-WB | WB | p-Value | |
|---|---|---|---|---|
| Essential amino acids | ||||
| Arginine | Arg | 4.77 ± 0.53 | 4.08 ± 0.23 | 0.098 |
| Histidine | His | 3.14 ± 0.18 | 3.08 ± 0.17 | 0.721 |
| Isoleucine | Ile | 4.93 ± 0.16 | 4.28 ± 0.31 | 0.020 |
| Leucine | Leu | 7.92 ± 0.20 | 7.33 ± 0.25 | 0.015 |
| Lysine | Lys | 7.70 ± 0.37 | 9.30 ± 0.81 | 0.025 |
| Methionine | Met | 3.73 ± 0.12 | 3.58 ± 0.14 | 0.229 |
| Phenylalanine | Phe | 4.32 ± 0.10 | 4.11 ± 0.12 | 0.055 |
| Threonine | Thr | 5.78 ± 0.12 | 5.71 ± 0.07 | 0.495 |
| Tryptophan | Trp | 0.80 ± 0.04 | 0.71 ± 0.09 | 0.186 |
| Valine | Val | 5.08 ± 0.15 | 4.72 ± 0.09 | 0.008 |
| Non-essential amino acids | ||||
| Alanine | Ala | 6.28 ± 0.08 | 6.09 ± 0.07 | 0.019 |
| Aspartic acid | Asp | 8.31 ± 0.33 | 7.53 ± 0.16 | 0.008 |
| Cysteine | Cys | 2.41 ± 0.08 | 2.75 ± 0.14 | 0.010 |
| Glutamic acid | Glu | 14.12 ± 0.39 | 13.09 ± 0.15 | 0.003 |
| Glycine | Gly | 4.14 ± 0.22 | 4.26 ± 0.06 | 0.418 |
| Hydroxyproline | Hyp | 1.33 ± 0.13 | 1.17 ± 0.12 | 0.181 |
| Proline | Pro | 5.13 ± 0.40 | 4.82 ± 0.19 | 0.298 |
| Serine | Ser | 4.01 ± 0.10 | 4.05 ± 0.03 | 0.548 |
| Tyrosine | Tyr | 3.07 ± 0.12 | 3.09 ± 0.07 | 0.761 |
| Cystine | C-C | 3.05 ± 1.25 | 6.23 ± 0.65 | 0.005 |
| Mineral | Non-WB | WB | p-Value |
|---|---|---|---|
| Aluminum (Al) | 0.0 ± 0.0 | 50.8 ± 20.8 | <0.001 |
| Calcium (Ca) | 55.9 ± 13.6 | 86.1 ± 39.9 | 0.01 |
| Chromium (Cr) | 5.7 ± 0.6 | 5.0 ± 0.6 | 0.69 |
| Copper (Cu) | 3.6 ± 3.7 | 2.2 ± 0.4 | 0.46 |
| Iron (Fe) | 8.6 ± 0.9 | 9.0 ± 1.2 | 0.01 |
| Magnesium (Mg) | 351.2 ± 68.9 | 284.9 ± 42.5 | 0.24 |
| Phosphorus (P) | 2219.2 ± 78.0 | 1958.2 ± 100.2 | 0.02 |
| Potassium (K) | 3153.9 ± 114.2 | 2758.4 ± 278.6 | 0.04 |
| Selenium (Se) | 0.3 ± 1.0 | 0.0 ± 0.0 | 0.35 |
| Sodium (Na) | 392.1 ± 70.1 | 545.4 ± 160.3 | 0.001 |
| Sulfur (S) | 11.1 ± 11.2 | 57.3 ± 30.1 | <0.001 |
| Zinc (Zn) | 14.3 ± 2.0 | 15.0 ± 2.7 | 0.06 |
| Property | Non-WB | WB | p-Value |
|---|---|---|---|
| pHu | 6.01 ± 0.06 | 6.04 ± 0.06 | 0.717 |
| L*-value | 52.21 ± 0.75 | 54.82 ± 0.54 | 0.018 |
| a*-value | 2.60 ± 0.23 | 2.66 ± 0.17 | 0.821 |
| b*-value | 0.53 ± 0.24 | 2.99 ± 0.99 | 0.065 |
| Drip loss (%) | 0.89 ± 0.14 | 2.04 ± 0.23 | 0.004 |
| Cook loss (%) | 19.54 ± 1.95 | 22.81 ± 3.04 | 0.396 |
| Shear force (N) | 39.79 ± 2.68 | 26.87 ± 4.09 | 0.032 |
| Hardness (N) | 27.48 ± 0.86 | 27.02 ± 3.17 | 0.894 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thanatsang, K.V.; Malila, Y.; Arayamethakorn, S.; Srimarut, Y.; Tatiyaborworntham, N.; Uengwetwanit, T.; Panya, A.; Rungrassamee, W.; Visessanguan, W. Nutritional Properties and Oxidative Indices of Broiler Breast Meat Affected by Wooden Breast Abnormality. Animals 2020, 10, 2272. https://doi.org/10.3390/ani10122272
Thanatsang KV, Malila Y, Arayamethakorn S, Srimarut Y, Tatiyaborworntham N, Uengwetwanit T, Panya A, Rungrassamee W, Visessanguan W. Nutritional Properties and Oxidative Indices of Broiler Breast Meat Affected by Wooden Breast Abnormality. Animals. 2020; 10(12):2272. https://doi.org/10.3390/ani10122272
Chicago/Turabian StyleThanatsang, Krittaporn V., Yuwares Malila, Sopacha Arayamethakorn, Yanee Srimarut, Nantawat Tatiyaborworntham, Tanaporn Uengwetwanit, Atikorn Panya, Wanilada Rungrassamee, and Wonnop Visessanguan. 2020. "Nutritional Properties and Oxidative Indices of Broiler Breast Meat Affected by Wooden Breast Abnormality" Animals 10, no. 12: 2272. https://doi.org/10.3390/ani10122272