The Regulatory Role of Apelin on the Appetite and Growth of Common Carp (Cyprinus Carpio L.)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Reagents
2.3. Gavage Treatment of Pyr-Apelin-13
2.4. Hypothalamus Preparation and Treatments
2.5. Isolation, Culture and Treatments of Primary Hepatocytes
2.6. RNA Extraction, Reverse-Transcription, and Real-Time Quantitative PCR
2.7. Statistical Analysis
3. Results
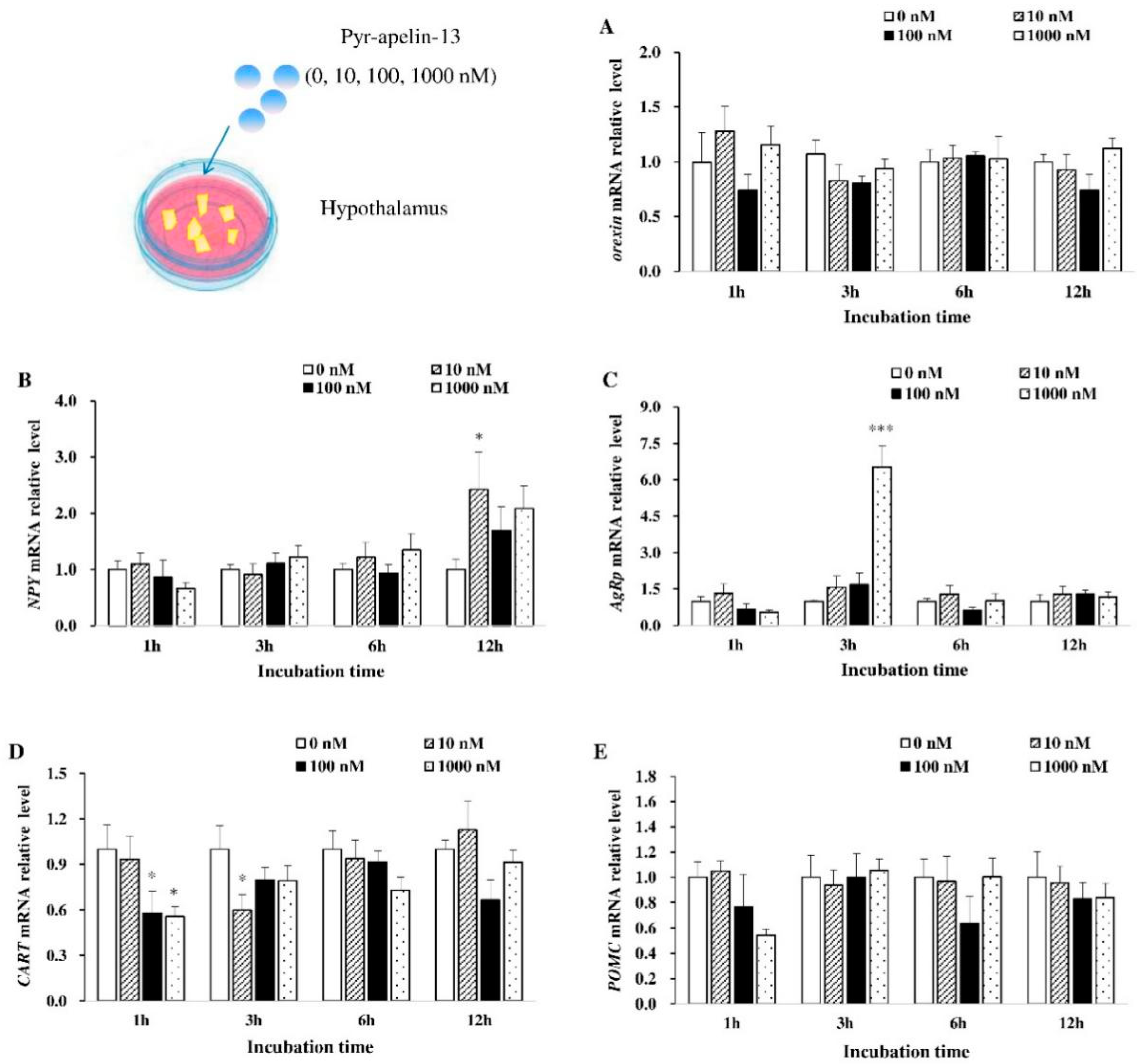
3.1. Effects of Pyr-Apelin-13 Incubation on the Expression of Appetite-Related Genes in the Hypothalamic Debris
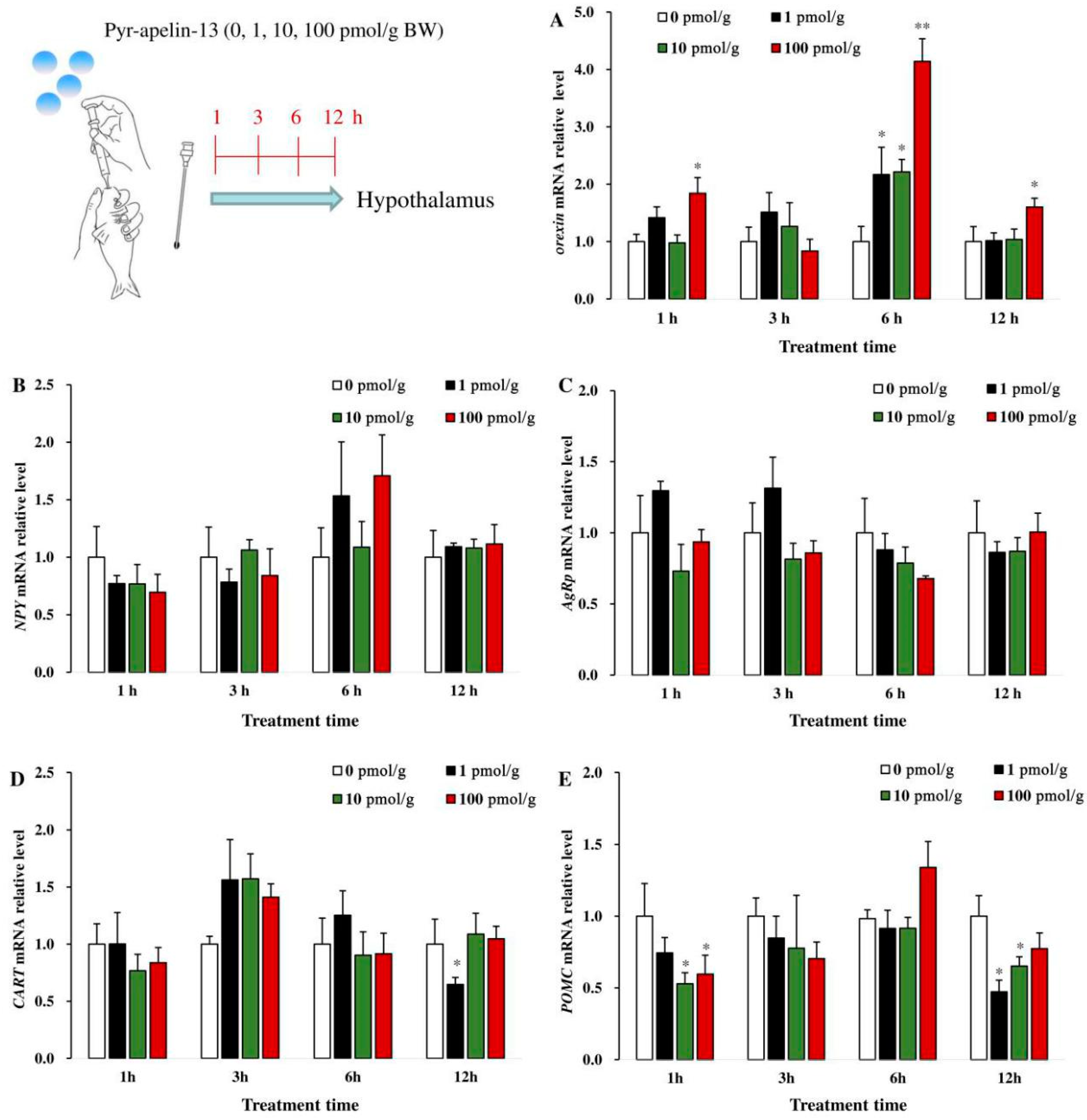
3.2. Effects of Perfusing Pyr-Apelin-13 on the Expression of Appetite-Related Genes in the Hypothalamus
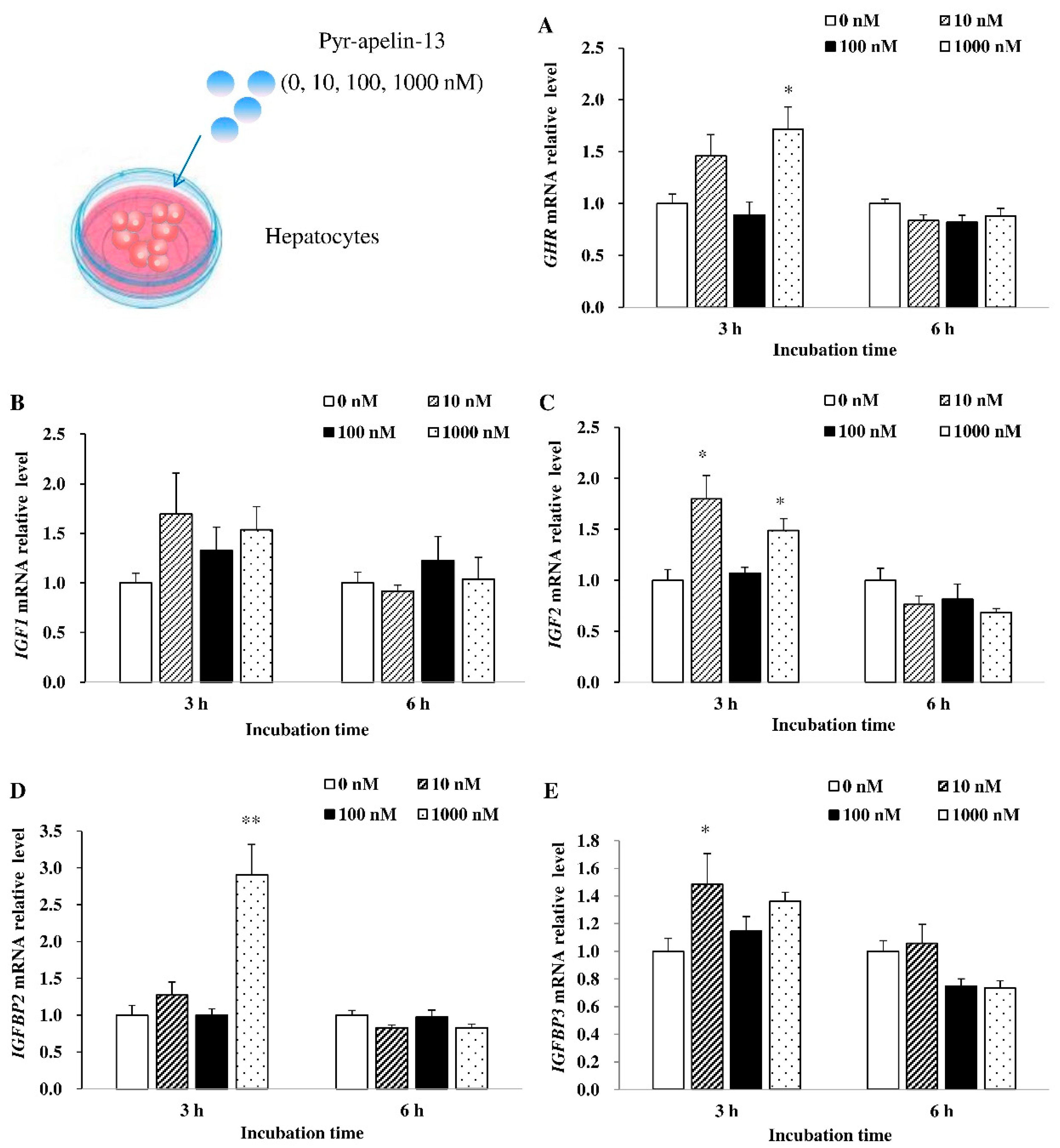
3.3. Effects of Pyr-Apelin-13 Incubation on the Expression of Growth-Related Genes in the Primary Hepatocytes
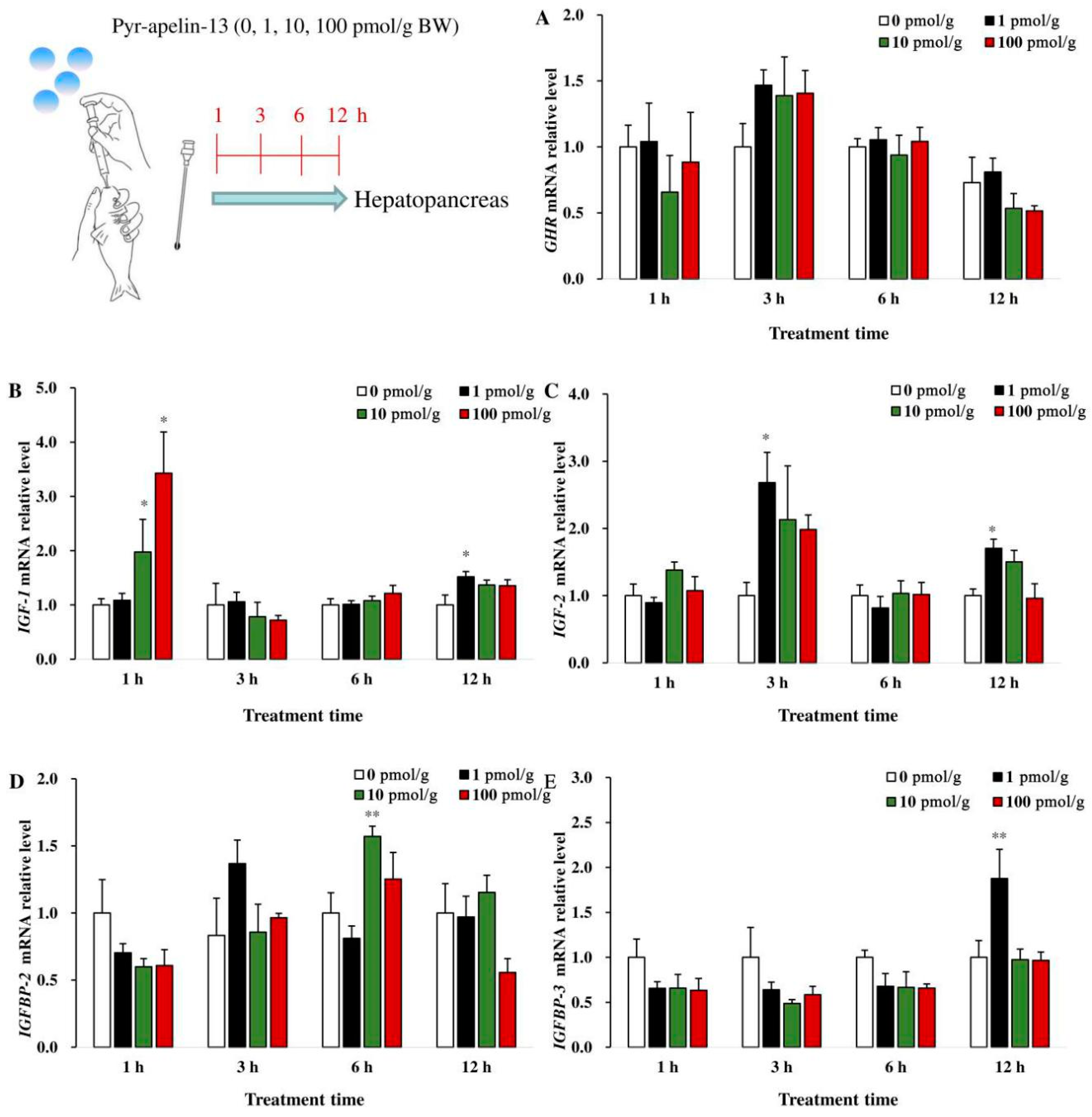
3.4. Effects of Perfusing Pyr-Apelin-13 on the Expression of Growth-Related Genes in the Hepatopancreas
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Dowd, B.F.; Heiber, M.; Chan, A.; Heng, H.H.Q.; Tsui, L.C.; Kennedy, J.L.; Shi, X.; Petronis, A.; George, S.R.; Nguyen, T. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene 1993, 136, 355–360. [Google Scholar] [CrossRef]
- Maguire, J.J.; Kleinz, M.J.; Pitkin, S.L.; Davenport, A.P. [Pyr(1)]Apelin-13 Identified as the Predominant Apelin Isoform in the Human Heart Vasoactive Mechanisms and Inotropic Action in Disease. Hypertension 2009, 54, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Messari, S.; Iturrioz, X.; Fassot, C.; De Mota, N.; Roesch, D.; Llorens-Cortes, C. Functional dissociation of apelin receptor signaling and endocytosis: Implications for the effects of apelin on arterial blood pressure. J. Neurochem. 2004, 90, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Wyatt, J.L. Apelin in goldfish (Carassius auratus): Cloning, distribution and role in appetite regulation. Peptides 2009, 30, 1434–1440. [Google Scholar] [CrossRef]
- Yan, X.; Qin, C.; Deng, D.; Yang, G.; Feng, J.; Lu, R.; Yang, L.; Meng, X.; Cao, X.; Wang, G.; et al. Apelin and apj in common carp (Cyprinus carpio L.): Molecular characterization, tissue expression and responses to periprandial changes and fasting–refeeding. Aquacult. Res. 2020, 51, 1012–1025. [Google Scholar] [CrossRef]
- Lin, F.J.; Wu, H.W.; Chen, H.; Xin, Z.M.; Yuan, D.Y.; Wang, T.; Liu, J.; Gao, Y.D.; Zhang, X.; Zhou, C.W.; et al. Molecular and physiological evidences for the role in appetite regulation of apelin and its receptor APJ in Ya-fish (Schizothorax prenanti). Mol. Cell. Endocrinol. 2014, 396, 46–57. [Google Scholar] [CrossRef]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Bertrand, C.; Valet, P.; Castan-Laurell, I. Apelin and energy metabolism. Front. Physiol. 2015, 6, 115–119. [Google Scholar] [CrossRef]
- O’Carroll, A.M.; Selby, T.L.; Palkovits, M.; Lolait, S.J. Distribution of mRNA encoding B78/apj, the rat homologue of the human APJ receptor, and its endogenous ligand apelin in brain and peripheral tissues. Biochim. Biophys. Acta. 2000, 1492, 72–80. [Google Scholar] [CrossRef]
- Hao, J.; Liu, Q.; Zhang, X.; Wu, Y.B.; Zhu, J.Y.; Qi, J.W.; Tang, N.; Wang, S.Y.; Wang, H.; Chen, D.F.; et al. The evidence of apelin has the bidirectional effects on feeding regulation in Siberian sturgeon (Acipenser baerii). Peptides 2017, 94, 78–85. [Google Scholar] [CrossRef]
- Volkoff, H. Appetite regulating peptides in red-bellied piranha, Pygocentrus nattereri: Cloning, tissue distribution and effect of fasting on mRNA expression levels. Peptides 2014, 56, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. Cloning and tissue distribution of appetite-regulating peptides in pirapitinga (Piaractus brachypomus). J. Anim. Physiol. Anim. Nutr. 2015, 99, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Saral, S.; Alkanat, M.; Sumer, A.; Canpolat, S. Apelin-13 increased food intake with serum ghrelin and leptin levels in male rats. Bratisl. Lek. Listy 2018, 119, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri, S.; Murphy, K.; Cohen, M.; Sujkovic, E.; Kennedy, A.; Dhillo, W.; Dakin, C.; Sajedi, A.; Ghatei, M.; Bloom, S. The effects of centrally administered apelin-13 on food intake, water intake and pituitary hormone release in rats. Biochem. Biophys. Res. Commun. 2002, 291, 1208–1212. [Google Scholar] [CrossRef]
- Volkoff, H. The Neuroendocrine Regulation of Food Intake in Fish: A Review of Current Knowledge. Front. Neurosci. 2016, 10, 540–570. [Google Scholar] [CrossRef] [Green Version]
- Penney, C.C.; Volkoff, H. Peripheral injections of cholecystokinin, apelin, ghrelin and orexin in cavefish (Astyanax fasciatus mexicanus): Effects on feeding and on the brain expression levels of tyrosine hydroxylase, mechanistic target of rapamycin and appetite-related hormones. Gen. Comp. Endocrinol. 2014, 196, 34–40. [Google Scholar] [CrossRef]
- Kim, K.S.; Seeley, R.J.; Sandoval, D.A. Signalling from the periphery to the brain that regulates energy homeostasis. Nat. Rev. Neurosci. 2018, 19, 185–196. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Jayakumar, J.; Rajeswar, J.J.; Velasco, C.; Unniappan, S. Nutrient regulation of endocrine factors influencing feeding and growth in gish. Front. Endocrinol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.L.; Piper, S.J.; Debra, Y.; Stephen, O.R.; Coll, A.P. A comparative Study of the central effects of specific proopiomelancortin (POMC)-derived melanocortin peptides on food intake and body weight in Pomc null mice. Endocrinology 2006, 12, 5940–5947. [Google Scholar]
- Yuan, D.; Gao, Y.; Zhang, X.; Wang, B.; Li, Z. NPY and NPY receptors in the central control of feeding and interactions with CART and MC4R in Siberian sturgeon. Gen. Comp. Endocrinol. 2019, 284, 113239–113246. [Google Scholar] [CrossRef]
- Kiris, G.A.; Kumlu, M.; Dikel, S. Stimulatory effects of neuropeptide Y on food intake and growth of Oreochromis niloticus. Aquaculture 2007, 264, 383–389. [Google Scholar] [CrossRef]
- Han, S.; Englander, E.W.; Gomez, G.A.; Greeley, G.H. Apelin regulates nuclear Factor-kappa B’s involvement in the inflammatory response of pancreatitis. Pancreas 2017, 46, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jia, J.R.; Yang, G.K.; Qin, J.K.; Zhang, C.; Zhang, Q.P.; Sun, C.Y.; Li, W.S. In vitro effects of somatostatin on the growth hormone-insulin-like growth factor axis in orange-spotted grouper (Epinephelus coioides). Gen. Comp. Endocrinol. 2016, 237, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.Y.; Yang, Y.J.; Chen, Q. Regulation of feeding behavior, gastrointestinal function and fluid homeostasis by apelin. Peptides 2013, 44, 87–92. [Google Scholar] [CrossRef]
- Volkoff, H. In vitro assessment of interactions between appetite-regulating peptides in brain of goldfish (Carassius auratus). Peptides 2014, 61, 61–68. [Google Scholar] [CrossRef]
- Ferrante, C.; Orlando, G.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di, N.C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; Vacca, M. Central apelin-13 administration modulates hypothalamic control of feeding. J. Biol. Regul. Homeost. Agents 2016, 30, 883. [Google Scholar]
- Hoskins, L.J.; Volkoff, H. Daily patterns of mRNA expression of two core circadian regulatory proteins, Clock2 and Per1, and two appetite-regulating peptides, OX and NPY, in goldfish (Carassius auratus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163, 127–136. [Google Scholar] [CrossRef]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S. AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; AVMA: Schaumburg, IL, USA, 2013. [Google Scholar]
- Pedro, N.D.; López-Patiño, M.A.; Guijarro, A.I.; Pinillos, M.L.; Delgado, M.J.; Alonso-Bedate, M. NPY receptors and opioidergic system are involved in NPY-induced feeding in goldfish. Peptides 2000, 21, 1495–1502. [Google Scholar] [CrossRef]
- Clarke, K.J.; Whitaker, K.W.; Reyes, T.M. Diminished metabolic responses to centrally-administered Apelin-13 in diet-induced obese rats fed a high-fat diet. J. Neuroendocrinol. 2009, 21, 83–89. [Google Scholar] [CrossRef]
- Escobar, S.; Fuentes, E.N.; Poblete, E.; Valdés, J.A.; Safian, D.; Reyes, A.E.; Álvarez, M.; Molina, A. Molecular cloning of IGF-1 and IGF-1 receptor and their expression pattern in the Chilean flounder (Paralichthys adspersus). Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2011, 159, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Tse, M.C.L.; Cheng, C.H.K.; Chan, K.M. Effects of chronic cysteamine treatment on growth enhancement and insulin-like growth factor I and II mRNA levels in common carp tissues. Br. J. Nutr. 2006, 96, 650–659. [Google Scholar] [PubMed]
- Douros, J.D.; Baltzegar, D.A.; Mankiewicz, J.; Taylor, J.; Yamaguchi, Y.; Lerner, D.T.; Seale, A.P.; Grau, E.G.; Breves, J.P.; Borski, R.J. Control of leptin by metabolic state and its regulatory interactions with pituitary growth hormone and hepatic growth hormone receptors and insulin like growth factors in the tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 2016, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Kenichi, M.; Cobb, L.J.; Malik, F.; Alden, H.; Bonnie, P.; Pintar, J.E.; Hiroyoshi, A.; Pinchas, C. PAPA-1 Is a nuclear binding partner of IGFBP-2 and modulates its growth-promoting actions. Mol. Endocrinol. 2009, 2, 169–175. [Google Scholar]
- Breves, J.P.; Springer-Miller, R.H.; Chenoweth, D.A.; Paskavitz, A.L.; Chang, A.Y.H.; Regish, A.M.; Einarsdottir, I.E.; Bjornsson, B.T.; McCormick, S.D. Cortisol regulates insulin-like growth-factor binding protein (igfbp) gene expression in Atlantic salmon parr. Mol. Cell. Endocrinol. 2020, 518, 110989. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Contents (%) | Proximate Composition | Contents (%) |
|---|---|---|---|
| Fish meal | 2.0 | Moisture | 9.05 |
| Soybean meal | 25.0 | Crude lipid | 9.56 |
| Rapeseed meal | 20.0 | Crude protein | 34.93 |
| Rice bran meal | 10.0 | Ash | 9.00 |
| Rice bran | 10.0 | Nitrogen-free extract | 29.26 |
| Fish oil | 2.0 | Note: 1 Vitamin premix supplied the following vitamins (kg): VA 800,000 IU, VB1 1500 mg, VB2 1250 mg, VC 2.5 g, VD3 160,000 IU, VE 15 g, VB12 4 mg, VK3 325 mg, VB6 1100 mg, Creatine 5.5 g, Folic acid 70 mg, Biotin 125 mg, Niacin 4 g, Pantothenic acid 4.5 g. 2 Mineral premix supplied the following minerals (kg): P 105 g, Ca 330 g, Mg 45 g, Fe 15 g, I 50 mg, Se 9 mg, Cu 0.35 g, Zn 3 g, Mn 1.5 g, Co 11 mg. | |
| Soybean oil | 3.0 | ||
| Sesame meal | 5.0 | ||
| Flour | 10.0 | ||
| Wheat bran | 10.0 | ||
| Ca(H2PO4)2 | 2.0 | ||
| Vitamin premix 1 | 0.2 | ||
| Mineral premix 2 | 0.6 | ||
| Choline chloride | 0.2 | ||
| Total | 100.0 | ||
| Gene | Accession No. | Forward (5′→3′) | Reverse (5′→3′) | Product Length (bp) |
|---|---|---|---|---|
| 18S | FJ710826.1 | GAGACTCCGGCTTGCTAAAT | CAGACCTGTTATTGCTCCATCT | 107 |
| β-actin | M24113.1 | TGCAAAGCCGGATTCGCTGG | AGTTGGTGACAATACCGTGC | 293 |
| orexin | XM_019086797.1 | CGTCAAGGTCCTGCAAATTATAC | CGATAGCCGCGTCGTTATTA | 105 |
| NPY | XM_019063564.1 | GCACTAAGACACTACATCAACCT | TGGGACTCTGTTTCACCAATC | 104 |
| AgRP | FR726954.1 | GCACCACAACTCTGCATTAAC | GGTCTCACCACATGATGTCTC | 101 |
| CART | AM498379.1 | TTCAGGGTGCCGAAATGG | CTGCTTCTCGTTGGTCAGATT | 101 |
| POMC | XM_019064968.1 | GCTTCTACCACGCAGACTTTA | AAGGGCACATAGGTGCTAATC | 101 |
| GHR | AY741100.1 | GAGTGATTGGAGTGGTGATTCT | GGTGCAGGAATAGGTGGTAAA | 91 |
| IGF1 | KP661168.1 | GGATATGGGCCTAGTTCAAGAC | TACGGGTGCACAATACATCTC | 105 |
| IGF2 | AF402958.1 | CCAGTTTCTATTCTTGCGGTTTC | CTGCGGCTTCTTTGTTCTTTC | 105 |
| IGFBP2 | FJ009001.1 | CCCACCTCTCCAATGATAAGG | GGGAAGGGATAGGAAGGTTTAG | 113 |
| IGFBP3 | FJ424519.1 | GCTGACCTCCCACTTGTATG | CACTCTCTCTGTCTCCTCTTCT | 103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Qin, C.; Yang, G.; Deng, D.; Yang, L.; Feng, J.; Mi, J.; Nie, G. The Regulatory Role of Apelin on the Appetite and Growth of Common Carp (Cyprinus Carpio L.). Animals 2020, 10, 2163. https://doi.org/10.3390/ani10112163
Yan X, Qin C, Yang G, Deng D, Yang L, Feng J, Mi J, Nie G. The Regulatory Role of Apelin on the Appetite and Growth of Common Carp (Cyprinus Carpio L.). Animals. 2020; 10(11):2163. https://doi.org/10.3390/ani10112163
Chicago/Turabian StyleYan, Xiao, Chaobin Qin, Guokun Yang, Dapeng Deng, Liping Yang, Junchang Feng, Jiali Mi, and Guoxing Nie. 2020. "The Regulatory Role of Apelin on the Appetite and Growth of Common Carp (Cyprinus Carpio L.)" Animals 10, no. 11: 2163. https://doi.org/10.3390/ani10112163