Identification of Candidate Genes and Pathways Associated with Obesity-Related Traits in Canines via Gene-Set Enrichment and Pathway-Based GWAS Analysis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Phenotype Assignment
2.2. Genomic DNA Extraction, SNP Genotyping and Quality Control
2.3. Genome-Wide Association Analysis
2.4. Gene-Set Enrichment and Pathway Analysis
3. Result and Discussion
3.1. Phenotypes
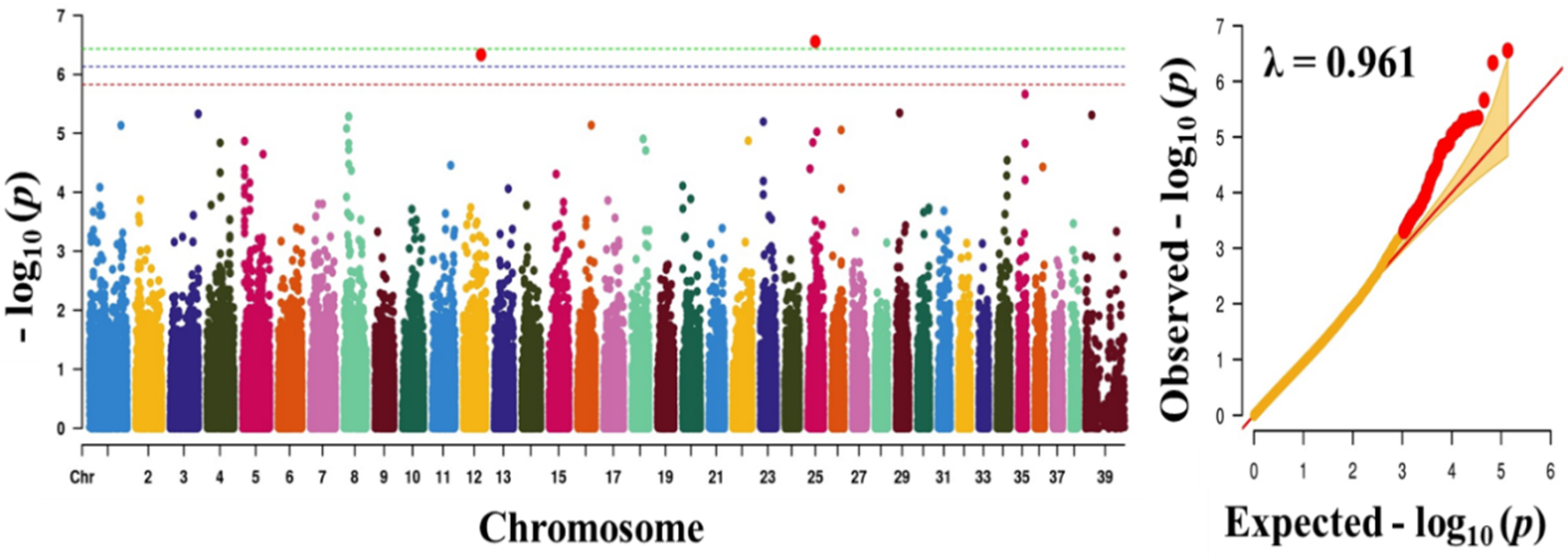
3.2. Genome-Wide Association Study
3.3. Gene-Set Enrichment and Pathway Analysis
3.3.1. Wnt Signaling, Adherens Junction, and Axon Guidance Pathways
3.3.2. Cancer-Related Pathways
3.3.3. Insulin Secretion Pathway
3.3.4. Other Enriched Gene Ontology Terms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- McGreevy, P.D.; Thomson, P.C.; Pride, C.; Fawcett, A.; Grassi, T.; Jones, B. Prevalence of obesity in dogs examined by Australian veterinary practices and the risk factors involved. Vet. Rec. 2005, 156, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Colliard, L.; Ancel, J.; Benet, J.-J.; Paragon, B.-M.; Blanchard, G. Risk Factors for Obesity in Dogs in France. J. Nutr. 2006, 136, 1951S–1954S. [Google Scholar] [CrossRef] [PubMed]
- Courcier, E.A.; Thomson, R.M.; Mellor, D.J.; Yam, P.S. An epidemiological study of environmental factors associated with canine obesity. J. Small Anim. Pr. 2010, 51, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Mankowska, M. Dog obesity—The need for identifying predisposing genetic markers. Res. Vet. Sci. 2013, 95, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Arslan, M.; Froguel, P. Genetics of Obesity in Consanguineous Populations: Toward Precision Medicine and the Discovery of Novel Obesity Genes. Obesity 2018, 26, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, W.-D.; Zhang, C.K.; Wang, Z.; Glessner, J.T.; Grant, S.F.A.; Zhao, H.; Hakonarson, H.; Price, R.A. A Genome-Wide Association Study on Obesity and Obesity-Related Traits. PLoS ONE 2011, 6, e18939. [Google Scholar] [CrossRef] [Green Version]
- Rudan, I.; Rudan, D.; Campbell, H.; Carothers, A.; Wright, A.; Smolej-Narancic, N.; Janicijevic, B.; Jin, L.; Chakraborty, R.; Deka, R.; et al. Inbreeding and risk of late onset complex disease. J. Med Genet. 2003, 40, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Xia, Z.; Chen, J.; Yu, J. Prevalence and risk factors for canine obesity surveyed in veterinary practices in Beijing, China. Prev. Vet. Med. 2013, 112, 438–442. [Google Scholar] [CrossRef]
- Yam, P.; Butowski, C.; Chitty, J.; Naughton, G.; Wiseman-Orr, M.; Parkin, T.; Reid, J. Impact of canine overweight and obesity on health-related quality of life. Prev. Vet. Med. 2016, 127, 64–69. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of Insulin Synthesis and Secretion and Pancreatic Beta-Cell Dysfunction in Diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- Lee, Y.; Berglund, E.D.; Yu, X.; Wang, M.-Y.; Evans, M.R.; Scherer, P.E.; Holland, W.L.; Charron, M.J.; Roth, M.G.; Unger, R.H. Hyperglycemia in rodent models of type 2 diabetes requires insulin-resistant alpha cells. Proc. Natl. Acad. Sci. USA 2014, 111, 13217–13222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Dominy, J.E.; Choi, Y.J.; Jurczak, M.J.; Tolliday, N.; Camporez, J.P.; Chim, H.; Lim, J.-H.; Ruan, H.-B.; Yang, X.; et al. Cyclin D1–Cdk4 controls glucose metabolism independently of cell cycle progression. Nat. Cell Biol. 2014, 510, 547–551. [Google Scholar] [CrossRef]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nat. Cell Biol. 2001, 409, 307–312. [Google Scholar] [CrossRef]
- Vittal, B.; Praveen, G.; Deepak, P. A study of body mass index in healthy individuals and its relationship with Fasting blood sugar. J. Clin. Diagn. Res. 2010, 4, 3421–3424. [Google Scholar]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nat. Cell Biol. 1998, 395, 763–770. [Google Scholar] [CrossRef]
- Grzemski, A.; Stachowiak, M.; Flisikowski, K.; Mankowska, M.; Krzeminska, P.; Gogulski, M.; Aleksiewicz, R.; Szydlowski, M.; Switonski, M.; Nowacka-Woszuk, J. FTO and IRX3 Genes are Not Promising Markers for Obesity in Labrador Retriever Dogs. Ann. Anim. Sci. 2019, 19, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Nishii, N.; Takasu, M.; Soe, O.K.; Maeda, S.; Ohba, Y.; Inoue-Murayama, M.; Kitagawa, H. Cloning, expression and investigation for polymorphisms of canine peroxisome proliferator-activated receptors. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 147, 690–697. [Google Scholar] [CrossRef]
- Scherag, A.; Jarick, I.; Grothe, J.; Biebermann, H.; Scherag, S.; Volckmar, A.-L.; Vogel, C.I.G.; Greene, B.; Hebebrand, J.; Hinney, A. Investigation of a Genome Wide Association Signal for Obesity: Synthetic Association and Haplotype Analyses at the Melanocortin 4 Receptor Gene Locus. PLoS ONE 2010, 5, e13967. [Google Scholar] [CrossRef] [Green Version]
- Raffan, E.; Dennis, R.J.; O’Donovan, C.J.; Becker, J.M.; Scott, R.A.; Smith, S.P.; Withers, D.J.; Wood, C.J.; Conci, E.; Clements, D.N.; et al. A Deletion in the Canine POMC Gene Is Associated with Weight and Appetite in Obesity-Prone Labrador Retriever Dogs. Cell Metab. 2016, 23, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Mankowska, M.; Krzeminska, P.; Graczyk, M.; Switonski, M. Confirmation that a deletion in the POMC gene is associated with body weight of Labrador Retriever dogs. Res. Vet. Sci. 2017, 112, 116–118. [Google Scholar] [CrossRef]
- Kao, P.Y.; Leung, K.H.; Chan, L.W.; Yip, S.P.; Yap, M.K. Pathway analysis of complex diseases for GWAS, extending to consider rare variants, multi-omics and interactions. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 335–353. [Google Scholar] [CrossRef] [Green Version]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Dadousis, C.; Pegolo, S.; Rosa, G.J.M.; Bittante, G.; Cecchinato, A. Genome-wide association and pathway-based analysis using latent variables related to milk protein composition and cheesemaking traits in dairy cattle. J. Dairy Sci. 2017, 100, 9085–9102. [Google Scholar] [CrossRef]
- Srikanth, K.; Lee, S.-H.; Chung, K.-Y.; Park, J.-E.; Jang, G.-W.; Park, M.-R.; Kim, N.Y.; Kim, T.-H.; Chai, H.-H.; Park, W.C.; et al. A Gene-Set Enrichment and Protein–Protein Interaction Network-Based GWAS with Regulatory SNPs Identifies Candidate Genes and Pathways Associated with Carcass Traits in Hanwoo Cattle. Genes 2020, 11, 316. [Google Scholar] [CrossRef] [Green Version]
- Dadousis, C.; Pegolo, S.; Rosa, G.J.M.; Gianola, D.; Bittante, G.; Cecchinato, A. Pathway-based genome-wide association analysis of milk coagulation properties, curd firmness, cheese yield, and curd nutrient recovery in dairy cattle. J. Dairy Sci. 2017, 100, 1223–1231. [Google Scholar] [CrossRef]
- Lin, P.-L.; Yu, Y.-W.; Chung, R.-H. Pathway Analysis Incorporating Protein-Protein Interaction Networks Identified Candidate Pathways for the Seven Common Diseases. PLoS ONE 2016, 11, e0162910. [Google Scholar] [CrossRef]
- Usui, S.; Yasuda, H.; Koketsu, Y. Characteristics of obese or overweight dogs visiting private Japanese veterinary clinics. Asian Pac. J. Trop. Biomed. 2016, 6, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Club, A.K. The Complete Dog Book; Howell Books: New York, NY, USA, 1998. [Google Scholar]
- Plassais, J.; Rimbault, M.; Williams, F.J.; Davis, B.W.; Schoenebeck, J.J.; Ostrander, E.A. Analysis of large versus small dogs reveals three genes on the canine X chromosome associated with body weight, muscling and back fat thickness. PLoS Genet. 2017, 13, e1006661. [Google Scholar] [CrossRef]
- Kang, J.M.; Seo, D.; Lee, S.H.; Lee, D.H.; Kim, Y.K.; Choi, B.H.; Lee, S.-H. Genome-wide association study to identify canine hip dysplasia loci in dogs. J. Anim. Sci. Technol. 2020, 62, 306–312. [Google Scholar] [CrossRef]
- Pegolo, S.; Mach, N.; Ramayo-Caldas, Y.; Schiavon, S.; Bittante, G.; Cecchinato, A. Integration of GWAS, pathway and network analyses reveals novel mechanistic insights into the synthesis of milk proteins in dairy cows. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Takita, C.; Wright, J.L.; Slifer, S.H.; Martin, E.R.; Urbanic, J.J.; Langefeld, C.D.; Lesser, G.J.; Shaw, E.G.; Hu, J.J. Genome-wide enriched pathway analysis of acute post-radiotherapy pain in breast cancer patients: A prospective cohort study. Hum. Genom. 2019, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.M.; Armstrong, P.J.; Kirk, C.A.; Klausner, J.S. Prevalence and risk factors for obesity in adult dogs from private US veterinary practices. Int. J. Appl. Res. Vet. Med. 2006, 4, 177–186. [Google Scholar]
- Antalis, T.M.; Bugge, T.H.; Wu, Q. Membrane-Anchored Serine Proteases in Health and Disease. Prog. Mol. Biol. Transl. Sci. 2011, 99, 1–50. [Google Scholar] [CrossRef]
- Hachem, J.-P.; Houben, E.; Crumrine, D.; Man, M.-Q.; Schurer, N.Y.; Roelandt, T.; Choi, E.H.; Uchida, Y.; Brown, B.E.; Feingold, K.R.; et al. Serine Protease Signaling of Epidermal Permeability Barrier Homeostasis. J. Investig. Dermatol. 2006, 126, 2074–2086. [Google Scholar] [CrossRef] [Green Version]
- Böttcher-Friebertshäuser, E. Membrane-Anchored Serine Proteases: Host Cell Factors in Proteolytic Activation of Viral Glycoproteins. In Activation of Viruses by Host Proteases; Springer: Cham, Switzerland, 2018; pp. 153–203. [Google Scholar]
- Marks, D.F. Homeostatic theory of obesity. Health Psychol. Open 2015, 2, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.K.; Fang, H.; Whistler, T.; Unger, E.R.; Rajeevan, M.S. Convergent Genomic Studies Identify Association of GRIK2 and NPAS2 with Chronic Fatigue Syndrome. Neuropsychobiology 2011, 64, 183–194. [Google Scholar] [CrossRef]
- Delgado, T.C. Glutamate and GABA in Appetite Regulation. Front. Endocrinol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Wang, W.; Zhang, S.; Wang, X.; Tang, Z.; Gu, J.; Li, J.; Huang, J. CACNA1B (Cav2. 2) overexpression and its association with clinicopathologic characteristics and unfavourable prognosis in non-small cell lung cancer. Dis. Markers 2017, 2017, 6136401. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Lai, M.-D.; Phan, N.N.; Sun, Z.; Lin, Y.-C. Meta-Analysis of Public Microarray Datasets Reveals Voltage-Gated Calcium Gene Signatures in Clinical Cancer Patients. PLoS ONE 2015, 10, e0125766. [Google Scholar] [CrossRef] [Green Version]
- Zhai, M.; Yang, D.; Yi, W.; Sun, W. Involvement of calcium channels in the regulation of adipogenesis. Adipocyte 2020, 9, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.-X. The Melanocortin-4 Receptor: Physiology, Pharmacology, and Pathophysiology. Endocr. Rev. 2010, 31, 506–543. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of Adipogenesis by Wnt Signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef]
- Fuster, J.J.; Zuriaga, M.A.; Ngo, D.T.-M.; Farb, M.G.; Aprahamian, T.R.; Yamaguchi, T.P.; Gokce, N.; Walsh, K. Noncanonical Wnt Signaling Promotes Obesity-Induced Adipose Tissue Inflammation and Metabolic Dysfunction Independent of Adipose Tissue Expansion. Diabetes 2015, 64, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Wang, J. Wnt/β-Catenin Signaling and Obesity. Front. Physiol. 2018, 9, 792. [Google Scholar] [CrossRef]
- Grant, S.F.A.; Thorleifsson, G.; Reynisdottir, I.; Benediktsson, R.; Manolescu, A.; Sainz, J.; Helgason, A.; Stefansson, H.; Emilsson, V.; Helgadottir, A.; et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 2006, 38, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, W.C.; Sorrenson, B.; Shepherd, P.R. The role of adherens junction proteins in the regulation of insulin secretion. Biosci. Rep. 2018, 38, 38. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Zhang, M.; Tong, M.; Yang, L.; Pang, L.; Chen, L.; Xu, G.; Chi, X.; Hong, Q.; Ni, Y.; et al. miR-148a is Associated with Obesity and Modulates Adipocyte Differentiation of Mesenchymal Stem Cells through Wnt Signaling. Sci. Rep. 2015, 5, srep09930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, T.W.; McPherson, M.; Darlington, L.G. Obesity and Cancer: Existing and New Hypotheses for a Causal Connection. EBioMedicine 2018, 30, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.-Y.; Im, K.-S.; Kim, N.-H.; Kim, H.-W.; Shin, J.-I.; Yhee, J.-Y.; Sur, J.-H. Effects of obesity and obesity-related molecules on canine mammary gland tumours. Vet. Pathol. 2015, 52, 1045–1051. [Google Scholar] [CrossRef] [Green Version]
- Thun, G.A.; Imboden, M.; Berger, W.; Rochat, T.; Probst-Hensch, N.M. The Association of a Variant in the Cell Cycle Control Gene CCND1 and Obesity on the Development of Asthma in the Swiss SAPALDIA Study. J. Asthma 2013, 50, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Zhang, L.; Biswas, S.; Schugar, R.C.; Brown, J.M.; Byzova, T.V.; Podrez, E.A. Akt3 inhibits adipogenesis and protects from diet-induced obesity via WNK1/SGK1 signalling. JCI Insight 2017, 2, e95687. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhu, H.; Qin, Q.; Yang, Y.; Yang, Y.; Cheng, H.; Sun, X. Genetic variants and risk of esophageal squamous cell carcinoma: A GWAS-based pathway analysis. Gene 2015, 556, 149–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-W.; Klemm, D.J.; Vinson, C.; Lane, M.D. Role of CREB in Transcriptional Regulation of CCAAT/Enhancer-binding Protein β Gene during Adipogenesis. J. Biol. Chem. 2004, 279, 4471–4478. [Google Scholar] [CrossRef] [Green Version]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Wang, Y.; Zhang, F.; Yao, F.; Sun, C. Calcium Signaling Pathways: Key Pathways in the Regulation of Obesity. Int. J. Mol. Sci. 2019, 20, 2768. [Google Scholar] [CrossRef] [Green Version]
- Kaser, S.; Sandhofer, A.; Föger, B.; Ebenbichler, C.F.; Igelseder, B.; Malaimare, L.; Paulweber, B.; Patsch, J.R. Influence of obesity and insulin sensitivity on phospholipid transfer protein activity. Diabetologia 2001, 44, 1111–1117. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Body Condition Score | 1 | 2 | 3 | 4 | 5 (Case) |
|---|---|---|---|---|---|
| Very thin | Underweight | Ideal body weight | Overweight | Obese | |
| Number of animal | 7 | 11 | 98 | 11 | 29 |
| Group | Number of Animals | Number of Control | Number of Case |
|---|---|---|---|
| Obesity | 153 | 124 (BCS 1–4) | 29 (BCS 5) |
| Body weight | 153 | 125 | 28 * |
| Blood sugar | 153 | 119 (≤120 mg/dL) | 34 (≥120 mg/dL) |
| No. | Breed Name | Number of Dogs Investigated | Obesity | Body Weight | Blood Sugar |
|---|---|---|---|---|---|
| 85 Females 68 Males | Case | Case | Case | ||
| 1 | Beagle | 3 | 1 | 1 | 1 |
| 2 | Bichon fris | 1 | 0 | 0 | 0 |
| 3 | Chihuahua | 6 | 0 | 0 | 2 |
| 4 | Cocker Spaniel | 8 | 1 | 1 | 1 |
| 5 | Dachshund | 3 | 0 | 0 | 0 |
| 6 | Doberman | 3 | 0 | 0 | 1 |
| 7 | German Shepherd | 1 | 0 | 0 | 1 |
| 8 | Golden Retriever | 1 | 0 | 0 | 0 |
| 9 | Maltese | 40 | 8 | 7 | 2 |
| 10 | Miniature Pinscher | 3 | 5 | 5 | 7 |
| 11 | Mixed | 19 | 1 | 1 | 1 |
| 12 | Parson Russell Terrier | 5 | 5 | 5 | 3 |
| 13 | Pomeranian | 7 | 2 | 2 | 6 |
| 14 | Poodle | 20 | 1 | 1 | 2 |
| 15 | Schnauzer | 6 | 1 | 1 | 1 |
| 16 | Shih tzu | 10 | 2 | 2 | 2 |
| 17 | Spitz | 5 | 0 | 0 | 0 |
| 18 | Yorkshire Terrier | 12 | 2 | 2 | 4 |
| Trait | SNP ID | Chr | Position | Freq | Gene | Type |
|---|---|---|---|---|---|---|
| Body weight | BICF2P1168261 | 9 | 47831552 | 6.38 × 10−9 | CACNA1B | Intron variant |
| G1314f25S201 | 26 | 29766195 | 4.09 × 10−8 | C22orf39 | Synonymous variant | |
| BICF2P940718 | 37 | 5443556 | 4.50 × 10−8 | U6 | Intergenic region | |
| BICF2P407675 | 1 | 1.06 × 108 | 1.77 × 10−7 | MYH14 | Intron variant | |
| BICF2P247463 | 39 | 39876923 | 2.86 × 10−7 | - | - | |
| BICF2P1124008 | 7 | 78278707 | 3.20 × 10−7 | PTPN2 | Intergenic region | |
| BICF2S23242598 | 7 | 78365053 | 3.20 × 10−7 | SEH1L | Intron variant | |
| Blood sugar | BICF2P1418953 | 25 | 27416887 | 2.78 × 10−7 | PRSS55 | Downstream_gene_variant |
| BICF2G630121162 | 12 | 60529545 | 4.64 × 10−7 | GRIK2 | Intergenic_region |
| Trait | Category | Term_ID | Term | Count | % | p-Value | Genes |
|---|---|---|---|---|---|---|---|
| Obesity | KEGG_PATHWAY | cfa04360 | Axon guidance | 10 | 0.015278 | 7.15 × 10−4 | DCC, MAPK1, NGEF, EPHA7, CXCR4, GNAI1, ROBO1, UNC5D, LRRC4C, EPHB1 |
| KEGG_PATHWAY | cfa04550 | Signaling pathways regulating pluripotency of stem cells | 10 | 0.015278 | 0.0015 | MAPK1, FGFR1, BMP2, ONECUT1, FZD1, FZD3, WNT11, FZD2, ZFHX3, KLF4 | |
| KEGG_PATHWAY | cfa05200 | Pathways in cancer | 18 | 0.0275 | 0.0016 | DCC, FGFR1, BMP2, COL4A1, BRAF, GNAI1, FGF9, RUNX1T1, FZD1, FZD3, FZD2, GLI3, CTNNA3, CTNNA2, LAMA2, MAPK1, CXCR4, WNT11 | |
| KEGG_PATHWAY | cfa04520 | Adherens junction | 7 | 0.010695 | 0.0026 | MAPK1, FGFR1, TJP1, PTPRM, SSX2IP, CTNNA3, CTNNA2 | |
| KEGG_PATHWAY | cfa05217 | Basal cell carcinoma | 6 | 0.009167 | 0.0040 | BMP2, FZD1, FZD3, WNT11, FZD2, GLI3 | |
| GOTERM_MF_DIRECT | GO:0016874 | Ligase activity | 7 | 1.682692 | 3.74 × 10−4 | HECW2, UBE3A, SUCLG2, HECTD3, SIAH1, SMURF1, NEDD4L | |
| GOTERM_BP_DIRECT | GO:0045892 | Negative regulation of transcription, DNA-templated | 13 | 3.125 | 0.0031 | RBFOX2, BCLAF1, RUNX1T1, FZD1, PAX2, CBFA2T3, ADIPOQ, GAS6, ZSCAN10, LHX1, ATP8B1, POU3F3, WNT11 | |
| GOTERM_BP_DIRECT | GO:0060022 | Hard palate development | 3 | 0.721154 | 0.0034 | FZD1, FZD2, MMP25 | |
| GOTERM_BP_DIRECT | GO:0034115 | Negative regulation of heterotypic cell-cell adhesion | 3 | 0.721154 | 0.0056 | APOA1, ADIPOQ, KLF4 | |
| GOTERM_BP_DIRECT | GO:0002062 | Chondrocyte differentiation | 5 | 1.201923 | 0.0057 | SNX19, FGFR1, BMP2, FGF9, NFIB | |
| Body weight | KEGG_PATHWAY | cfa05033 | Nicotine addiction | 8 | 0.900900 | 0.0021 | GABRG3, GRIA2, GRIA1, GABRB1, GABRA5, GRIN2A, GRIN3A, CACNA1B |
| KEGG_PATHWAY | cfa04080 | Neuroactive ligand-receptor interaction | 23 | 2.590090 | 0.0049 | GABRG3, GLRA1, GRIK2, GABRB1, OPRK1, GRIN3A, P2RY6, GRIA2, GRIA1, NMUR2, HRH4, ADRA1A, NMBR, PRL, CHRNE, PTAFR, GRID1, GHR | |
| KEGG_PATHWAY | cfa05206 | MicroRNAs in cancer | 13 | 1.463963 | 0.0268 | KIF23, PDGFA, SOCS1, MET, BMPR2, PIM1, TP63, ZEB1, IRS1, PDCD4, CCND1, CDKN2A, DNMT1 | |
| KEGG_PATHWAY | cfa04022 | cGMP-PKG signaling pathway | 14 | 1.576576 | 0.0293 | EDNRA, KCNU1, KCNMB4, PLCB4, TRPC6, ATP2A3, GTF2IRD1, ADRA1A, CREB5, NOS3, PLCB1, CACNA1D, IRS1, KCNMB2 | |
| KEGG_PATHWAY | cfa04310 | Wnt signaling pathway | 12 | 1.351351 | 0.0394 | DKK2, MAP3K7, CCND1, PLCB4, DKK1, VANGL1, PRICKLE1, MMP7, SIAH1 | |
| GOTERM_MF_DIRECT | GO:0004725 | Protein tyrosine phosphatase activity | 15 | 1.689189 | 5.27 × 10−5 | PTPRB, CDC14A, PTPN2, CDC14B, EPM2A, DUSP10, PTPN13, PTPRT, PTPRU, EYA3, EYA4, EYA1, DUSP26, UBASH3B, PTPN1 | |
| GOTERM_BP_DIRECT | GO:0045444 | Fat cell differentiation | 12 | 1.3513513 | 1.90 × 10−4 | BBS2, METTL8, CCND1, FAM120B, SMAD6, BBS9, FFAR2, SOCS1, OSBPL11, TTC8, PIAS1, PLCB1 | |
| GOTERM_CC_DIRECT | GO:0005794 | Golgi apparatus | 43 | 4.8423423 | 3.54 × 10−4 | GLIS3, ACHE, SYT4, RAB3GAP1, NOS3, CDK5RAP2, JAKMIP2, OLFM3, GOLM1, TERF2, CDK13, KLF5, NMNAT2, MSH6, CLN3, PLD1, MYO6, DNM1L, CCDC88A, LYN, ACO1, BEND5, GOLIM4, PKDCC, NMT2, ATF6, DUSP26, CPE, BACE2, SULF1, DYM, RAB14, SGCE, CWC22, EXT1 | |
| GOTERM_BP_DIRECT | GO:0007156 | Homophilic cell adhesion via plasma membrane adhesion molecules | 14 | 1.5765765 | 4.51 × 10−4 | CADM1, CLSTN2, SDK2, PCDH15, PCDH17, CDH8, CDH13, DSG2, CDH18, FAT1, CDH19, FAT2, CDH26, KIRREL3 | |
| GOTERM_CC_DIRECT | GO:0043025 | Neuronal cell body | 16 | 1.8018018 | 7.20 × 10−4 | GLRA1, GRIK2, DENND1A, GDPD5, GRIN3A, KLHL1, ALCAM, SEZ6L2, APOB, BRINP1, CPE, GRIA1, PSEN2, RAPGEF2, BRINP3, CACNA1B | |
| Blood sugar | KEGG_PATHWAY | cfa04919 | Thyroid hormone signaling pathway | 14 | 1.837270341 | 8.07 × 10−4 | ACTB, THRB, ATP1A1, RCAN2, PLCB3, CCND1, PLCB4, DIO2, GSK3B, PLCG2, PRKACB, PLCB1, AKT3, PIK3R1 |
| KEGG_PATHWAY | cfa05206 | MicroRNAs in cancer | 15 | 1.968503937 | 0.0019 | IRS2, E2F3, MCL1, MMP16, CDK6, ZEB2, ZEB1, PRKCE, TIMP3, RPS6KA5, CCNE1, CCND1, PLCG2, DNMT1, ZFPM2 | |
| KEGG_PATHWAY | cfa05223 | Non-small cell lung cancer | 9 | 1.181102362 | 0.0023 | FHIT, CCND1, E2F3, PLCG2, CDK6, EGF, AKT3, PIK3R1, EML4 | |
| KEGG_PATHWAY | cfa04916 | Melanogenesis | 12 | 1.57480315 | 0.0026 | WNT1, PLCB3, PLCB4, GSK3B, MITF, EDN1, FZD1, KITLG, PRKACB, PLCB1, WNT7A, CALM1 | |
| KEGG_PATHWAY | cfa04750 | Inflammatory mediator regulation of TRP channels | 12 | 1.57480315 | 0.0031 | PLCB3, IL1R1, PLCB4, PLCG2, IL1RAP, PLA2G6, PRKCH, PRKACB, PRKCE, PLCB1, PIK3R1, CALM1 | |
| GOTERM_MF_DIRECT | GO:0004222 | Metalloendopeptidase activity | 15 | 1.968504 | 0.0011 | ELP3, IRS2, BMP2, LYN, EDN1, HGF, DAB2, SEMA6D, FOXF1, SEMA3C, COL1A1, PDGFD, PAK1, PIK3R1, CSF1R | |
| GOTERM_BP_DIRECT | GO:0030335 | positive regulation of cell migration | 31 | 4.068241 | 0.0017 | FRK, THRB, MITF, EDN1, ZEB2, PRDM16, EPC1, REL, FOXF1, PRMT6, ETV6, DLG1, SIM2, ALX1, TBL1XR1, BMP2, ASXL1, ZHX2, LMCD1, SMYD2, SHOX2, HDAC4, CCND1, DKK1, DUSP26, PDE2A, HOPX, TFAP2B, DNMT1, RIPPLY2, BMP6 | |
| GOTERM_BP_DIRECT | GO:0000122 | negative regulation of transcription from RNA polymerase II promoter | 27 | 3.543307 | 0.0017 | CYB5R4, CAV2, GALNT1, RAB3C, PKHD1, SLC39A12, PINK1, ARFGEF1, SLC11A2, APP, BDNF, ECE1, PTK2B, FAT1, TMEM192, CDK5RAP2, DLG1, PTPRM, LYN, STC2, PRKCE, PDE2A, VAMP8, GSK3B, CYFIP2, AKAP6, SPAST | |
| GOTERM_CC_DIRECT | GO:0048471 | perinuclear region of cytoplasm | 11 | 1.44357 | 0.0039 | EPHA5, SEMA6A, ZNF280D, KIF5B, ANK3, ROBO1, SEMA3C, ETV1, RELN, UNC5D, CSF1R | |
| GOTERM_BP_DIRECT | GO:0007411 | axon guidance | 8 | 1.049869 | 0.0041 | SOX10, EDN3, SEMA6A, SEMA6D, SEMA3C, KITLG, ZEB2, ALX1 |
| Category | Term_ID | Term | Genes a |
|---|---|---|---|
| GOTERM_BP_DIRECT | GO:0045444 | Fat cell differentiation | AQP1 |
| GOTERM_MF_DIRECT | GO:0005509 | Calcium ion binding | CACNA1B, GRIK2 |
| GOTERM_CC_DIRECT | GO:0005737 | Cytoplasm | ROBO1, LDHB |
| GOTERM_CC_DIRECT | GO:0005634 | Nucleus | ATP2B1, CSRNP3, CSRNP2 |
| GOTERM_BP_DIRECT | GO:0015914 | Phospholipid transport | PCTP |
| GOTERM_BP_DIRECT | GO:0007417 | Central nervous system development | CHD7 |
| GOTERM_CC_DIRECT | GO:0009986 | Cell surface | IL1R1, ROBO1, WNT1 |
| KEGG_PATHWAY | cfa04310 | Wnt signaling pathway | WNT11, MAP3K7, WNT7A |
| KEGG_PATHWAY | cfa04520 | Adherens junction | MAPK1, CTNNA2, MAP3K7 |
| KEGG_PATHWAY | cfa05200 | Pathways in cancer | CCND1, FZD1, FZD3, FZD2, CTNNA1 |
| KEGG_PATHWAY | cfa04360 | Axon guidance | MAPK1, ROBO1 |
| KEGG_PATHWAY | cfa04911 | Insulin secretion | CACNA1B, CACNA1D, PLCB4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheet, S.; Krishnamoorthy, S.; Cha, J.; Choi, S.; Choi, B.-H. Identification of Candidate Genes and Pathways Associated with Obesity-Related Traits in Canines via Gene-Set Enrichment and Pathway-Based GWAS Analysis. Animals 2020, 10, 2071. https://doi.org/10.3390/ani10112071
Sheet S, Krishnamoorthy S, Cha J, Choi S, Choi B-H. Identification of Candidate Genes and Pathways Associated with Obesity-Related Traits in Canines via Gene-Set Enrichment and Pathway-Based GWAS Analysis. Animals. 2020; 10(11):2071. https://doi.org/10.3390/ani10112071
Chicago/Turabian StyleSheet, Sunirmal, Srikanth Krishnamoorthy, Jihye Cha, Soyoung Choi, and Bong-Hwan Choi. 2020. "Identification of Candidate Genes and Pathways Associated with Obesity-Related Traits in Canines via Gene-Set Enrichment and Pathway-Based GWAS Analysis" Animals 10, no. 11: 2071. https://doi.org/10.3390/ani10112071