Polymorphism of the Myostatin (MSTN) Gene in Landes and Kielecka Geese Breeds

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Measurements of Body Weight
2.3. Molecular Analysis
2.4. Polymerase Chain Reaction (PCR)
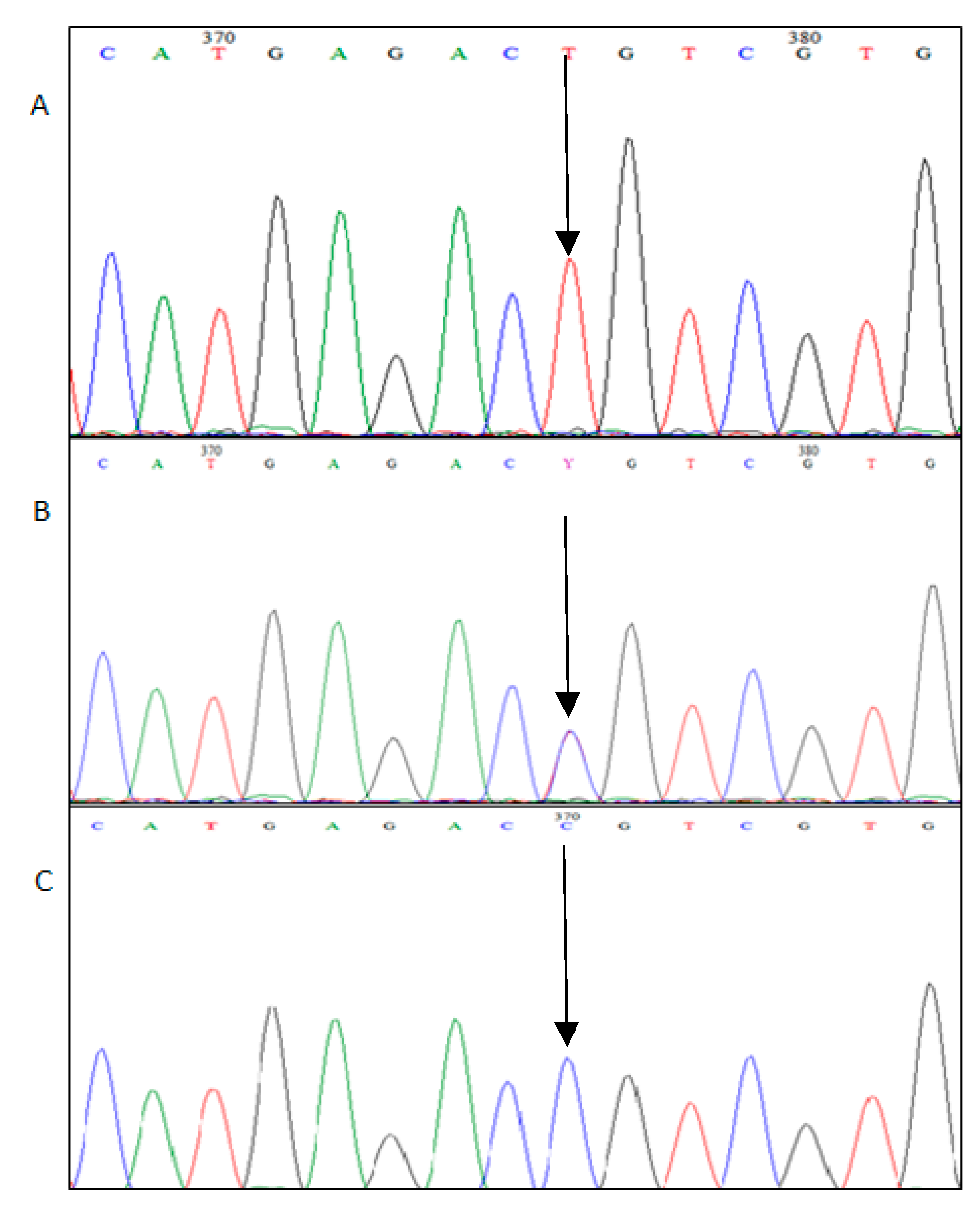
2.5. Sequencing Method
2.6. Alignment
2.7. Statistical Analyses
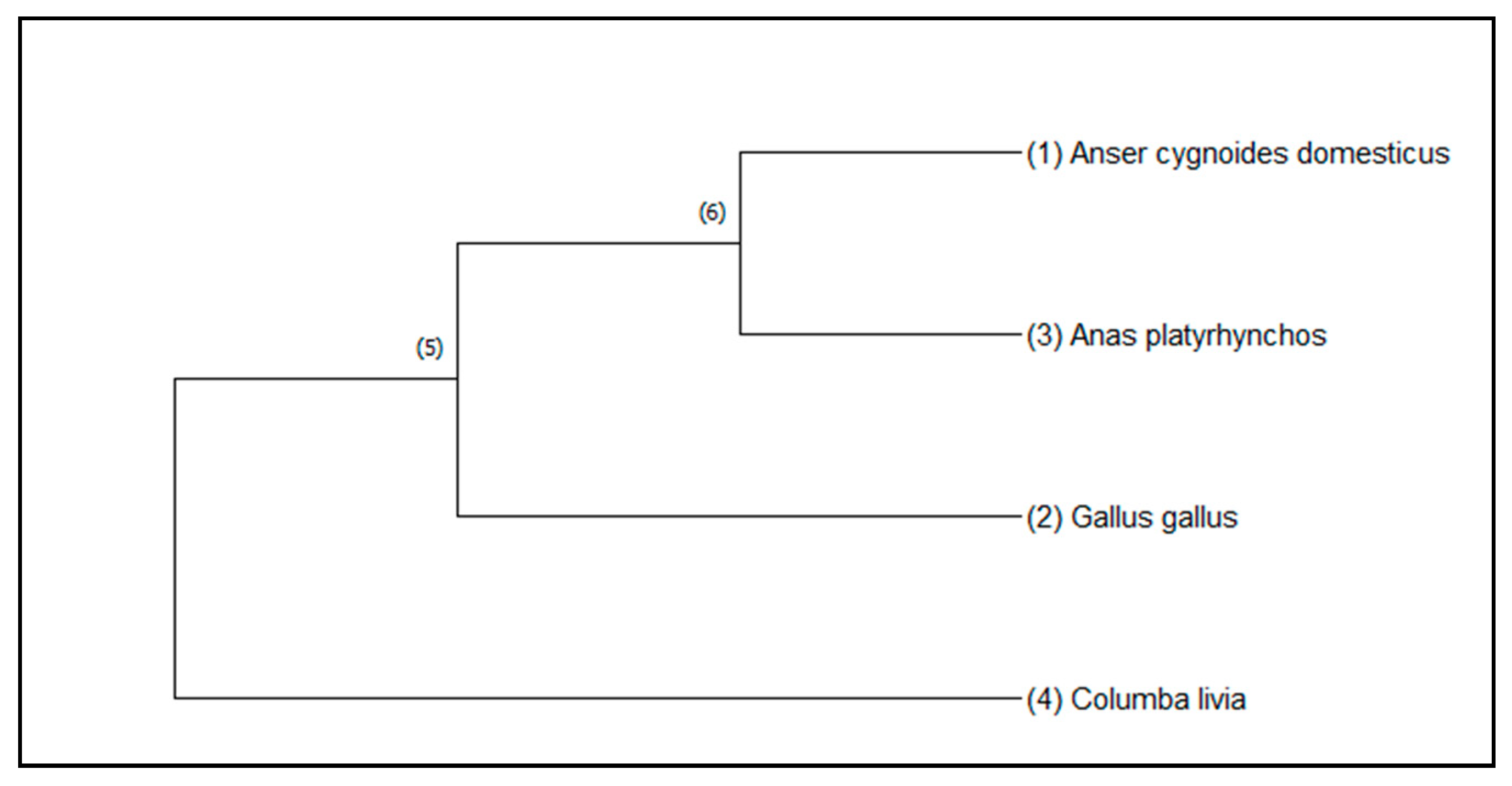
2.8. Phylogenetic Analyses
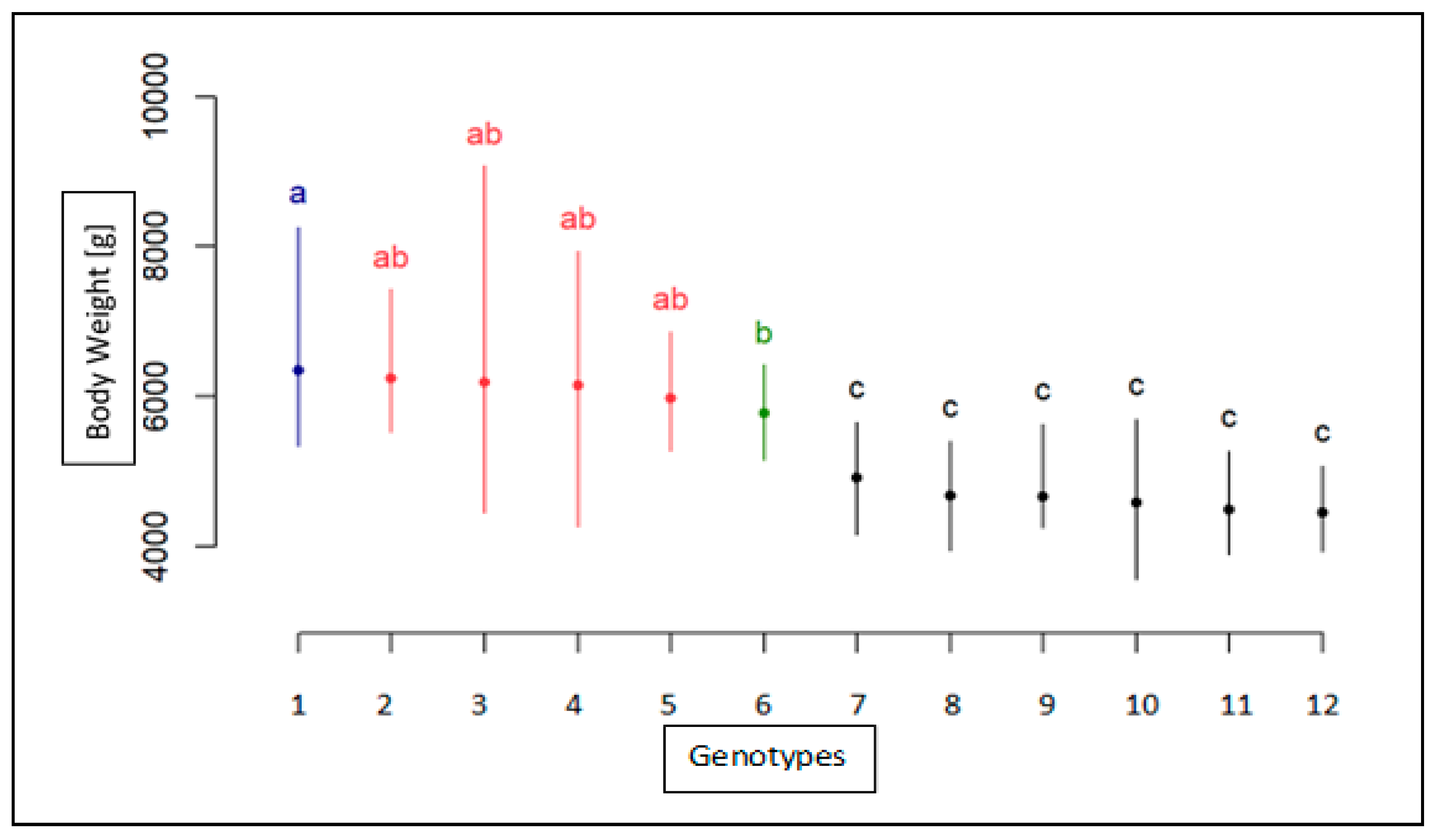
3. Results
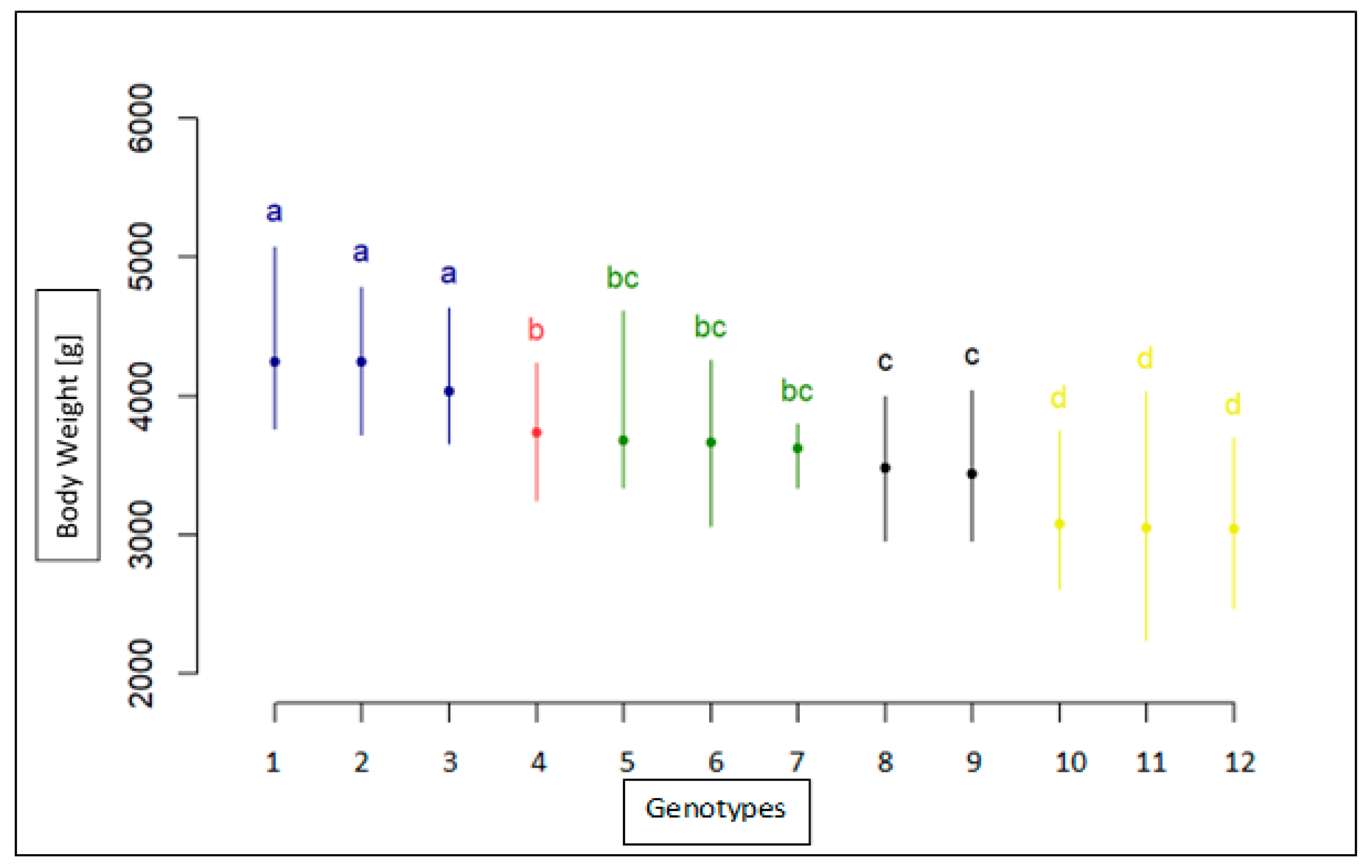
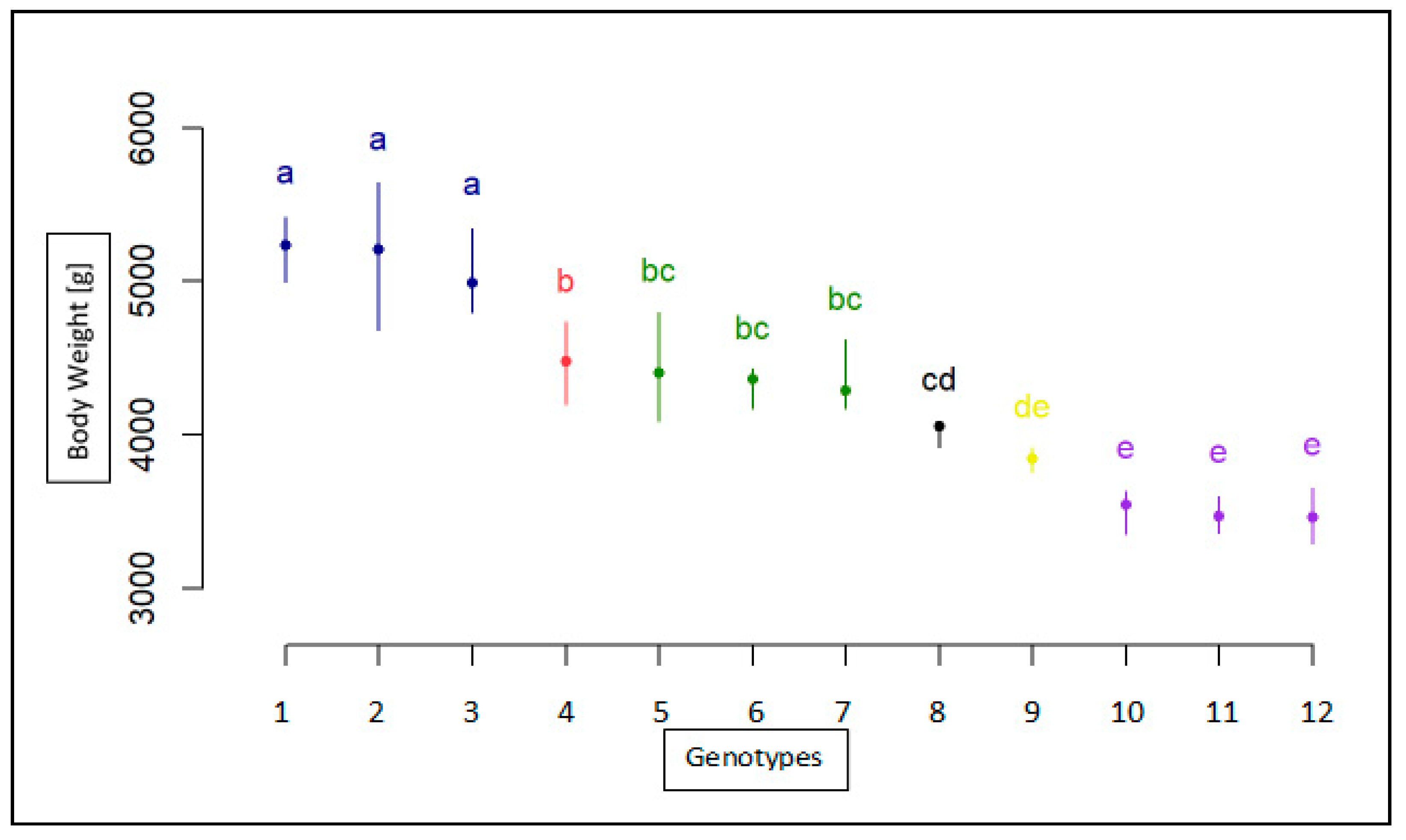
Association Analysis of SNP with Body Weight
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone Morphogenetic Protein-4 is Required for Mesoderm Formation and Patterning in the Mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef] [Green Version]
- Hogan, B.L. Bone Morphogenetic Proteins: Multifunctional Regulators of Vertebrate Development. Genes Dev. 1996, 10, 1580–1594. [Google Scholar] [CrossRef] [Green Version]
- Letterio, J.J.; Roberts, A.B. Regulation of Immune Responses by TGF-β. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef] [Green Version]
- Massagué, J. TGF-β Signal Transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef]
- Laitinen, M.; Vuojolainen, K.; Jaatinen, R.; Ketola, I.; Aaltonen, J.; Lehtonen, E.; Heikinheimo, M.; Ritvos, O. A Novel Growth Differentiation Factor-9 (GDF-9) Related Factor Is Co-Expressed with GDF-9 in Mouse Oocytes during Folliculogenesis. Mech. Dev. 1998, 78, 135–140. [Google Scholar] [CrossRef]
- Aaltonen, J.; Laitinen, M.P.; Vuojolainen, K.; Jaatinen, R.; Horelli-Kuitunen, N.; Seppä, L.; Louhio, H.; Tuuri, T.; Sjöberg, J.; Bützow, R.; et al. Human Growth Differentiation Factor 9 (GDF-9) and Its Novel Homolog GDF-9B Are Expressed in Oocytes during Early Folliculogenesis. J. Clin. Endocrinol. Metab. 1999, 84, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Elvin, J.A.; Yan, C.; Matzuk, M.M. Oocyte-Expressed TGF-β Superfamily Members in Female Fertility. Mol. Cell. Endocrinol. 2000, 159, 1–5. [Google Scholar] [CrossRef]
- Smołucha, G.; Piestrzy, A. Genetyczny aspekt wysokiej plenności u owiec. Cz. II. Wiadomości Zootechniczne 2012, 2, 5–10. [Google Scholar]
- Bellinge, R.H.S.; Liberles, D.A.; Iaschi, S.P.A.; O’Brien, P.A.; Tay, G.K. Myostatin and Its Implications on Animal Breeding: A Review. Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/j.1365-2052.2004.01229.x (accessed on 26 September 2019).
- Ahad, W.A.; Andrabi, M.; Beigh, S.A.; Bhat, R.A.; Shah, R.A. Applications of Myostatin (MSTN) Gene in the Livestock Animals and Humans: A Review. Int. J. Curr. Microbiol. App. Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Kambadur, R.; Sharma, M.; Smith, T.P.; Bass, J.J. Mutations in Myostatin (GDF8) in Double-Muscled Belgian Blue and Piedmontese Cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.-M.; Eychenne, F.; et al. A Mutation Creating a Potential Illegitimate MicroRNA Target Site in the Myostatin Gene Affects Muscularity in Sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Hadjipavlou, G.; Matika, O.; Clop, A.; Bishop, S.C. Two Single Nucleotide Polymorphisms in the Myostatin (GDF8) Gene Have Significant Association with Muscle Depth of Commercial Charollais Sheep. Anim. Genet. 2008, 39, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Forrest, R.H.; Hickford, J.G.H. Genetic Variations in the Myostatin Gene (MSTN) in New Zealand Sheep Breeds. Mol. Biol. Rep. 2013, 40, 6379–6384. [Google Scholar] [CrossRef]
- Mosher, D.S.; Quignon, P.; Bustamante, C.D.; Sutter, N.B.; Mellersh, C.S.; Parker, H.G.; Ostrander, E.A. A Mutation in the Myostatin Gene Increases Muscle Mass and Enhances Racing Performance in Heterozygote Dogs. PLoS Genet. 2007, 3, e79. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lee, S.-J. Double Muscling in Cattle Due to Mutations in the Myostatin Gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [Green Version]
- Hill, E.W.; Gu, J.; Eivers, S.S.; Fonseca, R.G.; McGivney, B.A.; Govindarajan, P.; Orr, N.; Katz, L.M.; MacHugh, D. A Sequence Polymorphism in MSTN Predicts Sprinting Ability and Racing Stamina in Thoroughbred Horses. PLoS ONE 2010, 5, e8645. [Google Scholar] [CrossRef]
- Xu, T.S.; Gu, L.H.; Zhang, X.H.; Ye, B.G.; Liu, X.L.; Hou, S.S. Characterization of Myostatin Gene (MSTN) of Pekin Duck and the Association of Its Polymorphism with Breast Muscle Traits. Genet. Mol. Res. 2013, 12. [Google Scholar] [CrossRef]
- Zhao, Z.-H.; Li, H.; Yi, H.-J.; Peng, B.-X. The Correlation Between Polymorphisms of the MSTN Gene and Slaughter Traits in Sansui Ducks. Pak. J. Zool. 2016, 48, 1283–1290. [Google Scholar]
- Haraf, G.; Wołoszyn, J.; Okruszek, A.; Orkusz, A.; Wereńska, M. Nutritional Value of Proteins and Lipids in Breast Muscle of Geese from Four Different Polish Genotypes. Eur. Poult. Sci. 2018, 82. [Google Scholar] [CrossRef]
- Kapkowska, E.; Gumulka, M.; Rabsztyn, A.; Poltowicz, K.; Andres, K. Comparative Study on Fattening Results of Zatorska and White Koluda (R) Geese. Ann. Anim. Sci. 2011, 11, 207–217. [Google Scholar]
- Okruszek, A.; Wołoszyn, J.; Haraf, G.; Orkusz, A.; Wereńska, M. Chemical Composition and Amino Acid Profiles of Goose Muscles from Native Polish Breeds. Poult. Sci. 2013, 92, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Smolucha, G.; Kozubska-Sobocinska, A.; Koseniuk, A.; Lisowski, M.; Grajewski, B.; Zubrzycki, I. Identification of SNP Polymorphism in the Growth Hormone and Myostatin Genes of the Two Polish Geese Breeds. In Comparative Cytogenetics; Pensoft Publishers: Sofia, Bulgaria, 2018; Volume 12, p. 349. [Google Scholar]
- National Research Institute of Animal Production (NRIAP): Geese Landes Biodiversity, Genetic Resources Programme. Available online: http://www.bioroznorodnosc.izoo.krakow.pl/drob/gesi/charakterystyka_ras/landes (accessed on 26 September 2019).
- National Research Institute of Animal Production (NRIAP): Geese Kielecka Biodiversity, Genetic Resources Programme. Available online: http://www.bioroznorodnosc.izoo.krakow.pl/drob/gesi/charakterystyka_ras/kielecka (accessed on 26 September 2019).
- National Research Institute of Animal Production (NRIAP): Geese Biodiversity, Genetic Resources Programme. Available online: http://www.bioroznorodnosc.izoo.krakow.pl/drob/gesi (accessed on 26 September 2019).
- Hall, T.A. BioEdit A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95–98—References—Scientific Research Publishing. Available online: https://www.scirp.org/(S(lz5mqp453edsnp55rrgjct55))/reference/ReferencesPapers.aspx?ReferenceID=1383440 (accessed on 26 September 2019).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornowicz, E.; Węglarzy, K.; Pietrzak, M.; Bereza, M. Slaughter and Meat Traits of Domestic Geese of the Southern Breeds. Wiad. Zootech. 2012, 4, 5–16. [Google Scholar]
- Stinckens, A.; Luyten, T.; Bijttebier, J.; Van den Maagdenberg, K.; Dieltiens, D.; Janssens, S.; De Smet, S.; Georges, M.; Buys, N. Characterization of the Complete Porcine MSTN Gene and Expression Levels in Pig Breeds Differing in Muscularity. Anim. Genet. 2008, 39, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk, M.; Kaczor, U.; Kulisa, M. MSTN gene polymorphism in livestock animals. Postepy Hig. Med. Doswiadczalnej Online 2014, 68, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ding, F.; Wang, J.; Dai, G.; Xie, K.; Zhang, L.; Wang, W.; Zhou, S. Polymorphism in Exons of the Myostatin Gene and its Relationship with Body Weight Traits in the Bian Chicken. Biochem. Genet. 2011, 49, 9–19. [Google Scholar] [CrossRef]
- Wu, W.; Guo, X.; Zhang, L.; Hu, D. Association between Single Nucleotide Polymorphisms of Fatty Acid Synthase and Fat Deposition in the Liver of the Overfed Goose. Asian Australas. J. Anim. Sci. 2014, 27, 1244–1249. [Google Scholar] [CrossRef] [Green Version]
- Matoulkova, E.; Michalova, E.; Vojtesek, B.; Hrstka, R. The Role of the 3′ Untranslated Region in Post-Transcriptional Regulation of Protein Expression in Mammalian Cells. RNA Biol. 2012, 9, 563–576. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Tm * (°C) | Product Length (bp) | |
|---|---|---|---|---|
| MSTNex3 F | Exon 3 | 5′ TTCCGGTTCCTTTTCCTCTT 3′ | 61 | 577 |
| MSTNex3 R | 5′ TCTGCAGCTTGTGTTGCTCT 3′ | |||
| MSTNex1 F | Exon 1 | 5′ TCAGATTGCATTTGCTTTCA 3′ | 61 | 497 |
| MSTNex1 R | 5′ AGACGAAAGCAGCAGGGTTA 3′ | |||
| MSTNex2 F | Exon 2 | 5′ TTTTTGTTCCCTGTTCAGTAATC 3′ | 61 | 473 |
| MSTNex2 R | 5′ TGCTTTCCAATAAAATGCAAGA 3′ | |||
| Breed | Genotypes | Alleles | HWE p-Value | |||
|---|---|---|---|---|---|---|
| CC | CT | TT | C | T | ||
| Landes | 0.292 | 0.496 | 0.212 | 0.54 | 0.46 | 0.39 |
| Kielecka | 0.248 | 0.458 | 0.294 | 0.477 | 0.523 | 0.99 |
| Genotype | Sex | Breed | BW 8 Week [g] | std | N | Min [g] | Max [g] |
|---|---|---|---|---|---|---|---|
| CC | F | KIELECKA | 3051 | 352 | 24 | 2240 | 4030 |
| CC | F | LANDES | 3678 | 264 | 29 | 3340 | 4610 |
| CC | M | KIELECKA | 3623 | 248 | 3 | 3340 | 3800 |
| CC | M | LANDES | 4032 | 318 | 11 | 3660 | 4630 |
| CT | F | KIELECKA | 3043 | 253 | 33 | 2470 | 3700 |
| CT | F | LANDES | 3732 | 237 | 49 | 3250 | 4240 |
| CT | M | KIELECKA | 3479 | 270 | 17 | 2960 | 4000 |
| CT | M | LANDES | 4245 | 289 | 19 | 3720 | 4780 |
| TT | F | KIELECKA | 3079 | 256 | 24 | 2610 | 3750 |
| TT | F | LANDES | 3665 | 298 | 22 | 3060 | 4260 |
| TT | M | KIELECKA | 3440 | 342 | 7 | 2960 | 4040 |
| TT | M | LANDES | 4246 | 495 | 7 | 3760 | 5070 |
| Genotype | Sex | Breed | BW 12 Week [g] | std | N | Min [g] | Max [g] |
|---|---|---|---|---|---|---|---|
| CC | F | KIELECKA | 3540 | 285 | 24 | 3090 | 4400 |
| CC | F | LANDES | 4363 | 305 | 29 | 3930 | 5510 |
| CC | M | KIELECKA | 4477 | 536 | 3 | 3960 | 5030 |
| CC | M | LANDES | 4989 | 381 | 11 | 4170 | 5400 |
| CT | F | KIELECKA | 3463 | 272 | 33 | 2900 | 4070 |
| CT | F | LANDES | 4287 | 717 | 49 | 4195 | 5070 |
| CT | M | KIELECKA | 4053 | 357 | 17 | 3470 | 4770 |
| CT | M | LANDES | 5233 | 326 | 19 | 4760 | 6010 |
| TT | F | KIELECKA | 3471 | 308 | 24 | 2480 | 4000 |
| TT | F | LANDES | 4405 | 434 | 22 | 3400 | 5100 |
| TT | M | KIELECKA | 3843 | 315 | 7 | 3320 | 4370 |
| TT | M | LANDES | 5207 | 747 | 7 | 4350 | 6460 |
| Genotype | Sex | Breed | BW 95 Week [g] | std | N | Min [g] | Max [g] |
|---|---|---|---|---|---|---|---|
| CC | F | KIELECKA | 4446 | 289 | 24 | 3935 | 5070 |
| CC | F | LANDES | 6152 | 697 | 29 | 4260 | 7945 |
| CC | M | KIELECKA | 4905 | 748 | 3 | 4160 | 5655 |
| CC | M | LANDES | 5776 | 451 | 11 | 5145 | 6425 |
| CT | F | KIELECKA | 4581 | 519 | 33 | 3555 | 5695 |
| CT | F | LANDES | 6345 | 625 | 49 | 5340 | 8260 |
| CT | M | KIELECKA | 4673 | 405 | 17 | 3940 | 5400 |
| CT | M | LANDES | 5968 | 469 | 19 | 5275 | 6870 |
| TT | F | KIELECKA | 4489 | 383 | 25 | 3890 | 5265 |
| TT | F | LANDES | 6191 | 973 | 22 | 4445 | 9085 |
| TT | M | KIELECKA | 4659 | 467 | 7 | 4250 | 5625 |
| TT | M | LANDES | 6236 | 722 | 7 | 5520 | 7430 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smołucha, G.; Kozubska-Sobocińska, A.; Koseniuk, A.; Żukowski, K.; Lisowski, M.; Grajewski, B. Polymorphism of the Myostatin (MSTN) Gene in Landes and Kielecka Geese Breeds. Animals 2020, 10, 10. https://doi.org/10.3390/ani10010010
Smołucha G, Kozubska-Sobocińska A, Koseniuk A, Żukowski K, Lisowski M, Grajewski B. Polymorphism of the Myostatin (MSTN) Gene in Landes and Kielecka Geese Breeds. Animals. 2020; 10(1):10. https://doi.org/10.3390/ani10010010
Chicago/Turabian StyleSmołucha, Grzegorz, Anna Kozubska-Sobocińska, Anna Koseniuk, Kacper Żukowski, Mirosław Lisowski, and Bartosz Grajewski. 2020. "Polymorphism of the Myostatin (MSTN) Gene in Landes and Kielecka Geese Breeds" Animals 10, no. 1: 10. https://doi.org/10.3390/ani10010010