
Carboxy-Terminal Processing Protease Controls Production of Outer Membrane Vesicles and Biofilm in Acinetobacter baumannii



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media and Culture Conditions
2.2. Molecular Cloning, Expression and Purification of Ctp
2.3. Polyclonal Antibody Production
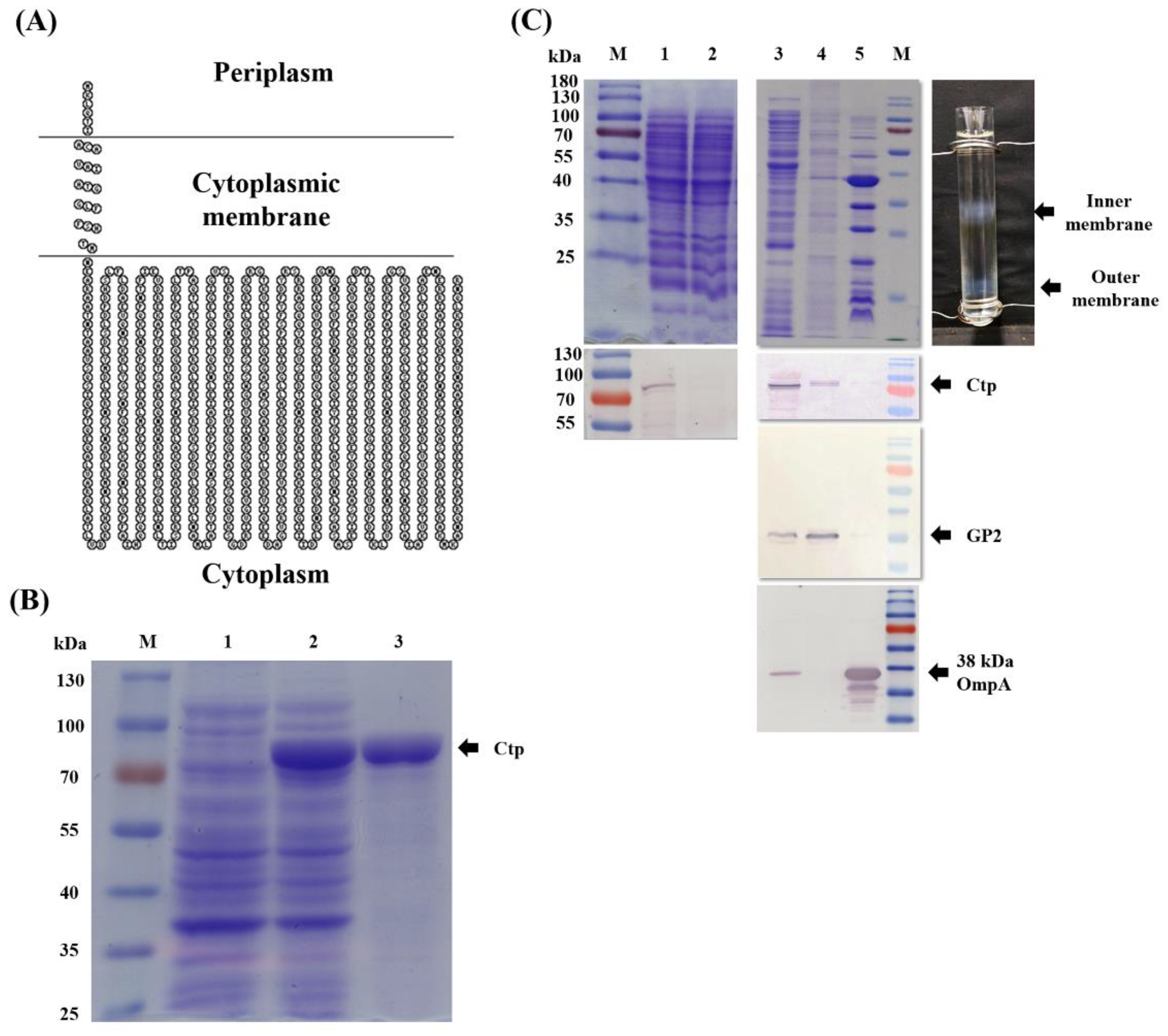
2.4. Subcellular Localization of Ctp
2.5. Capsular Polysaccharide Visualization and Quantification
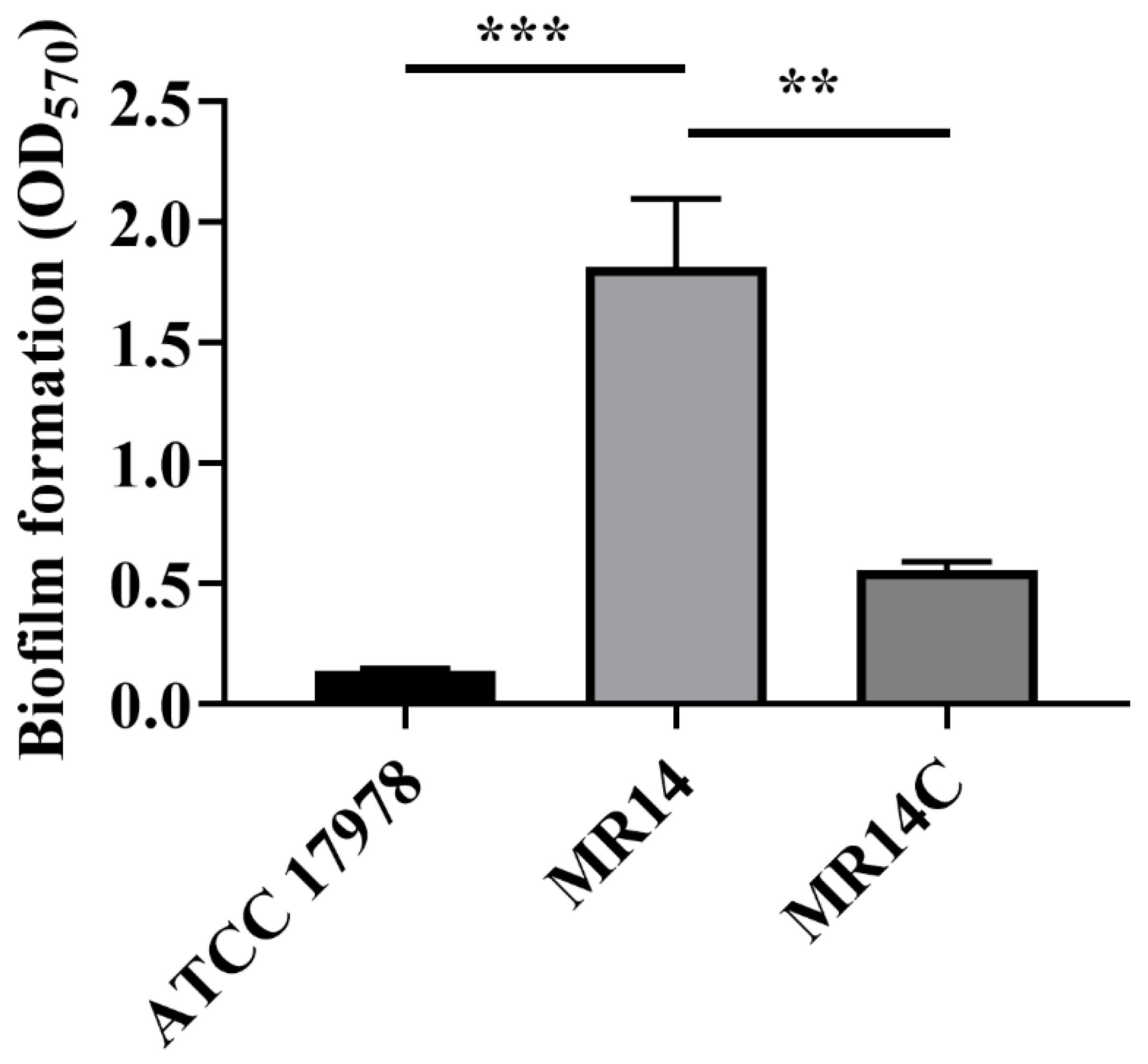
2.6. Biofilm Assay
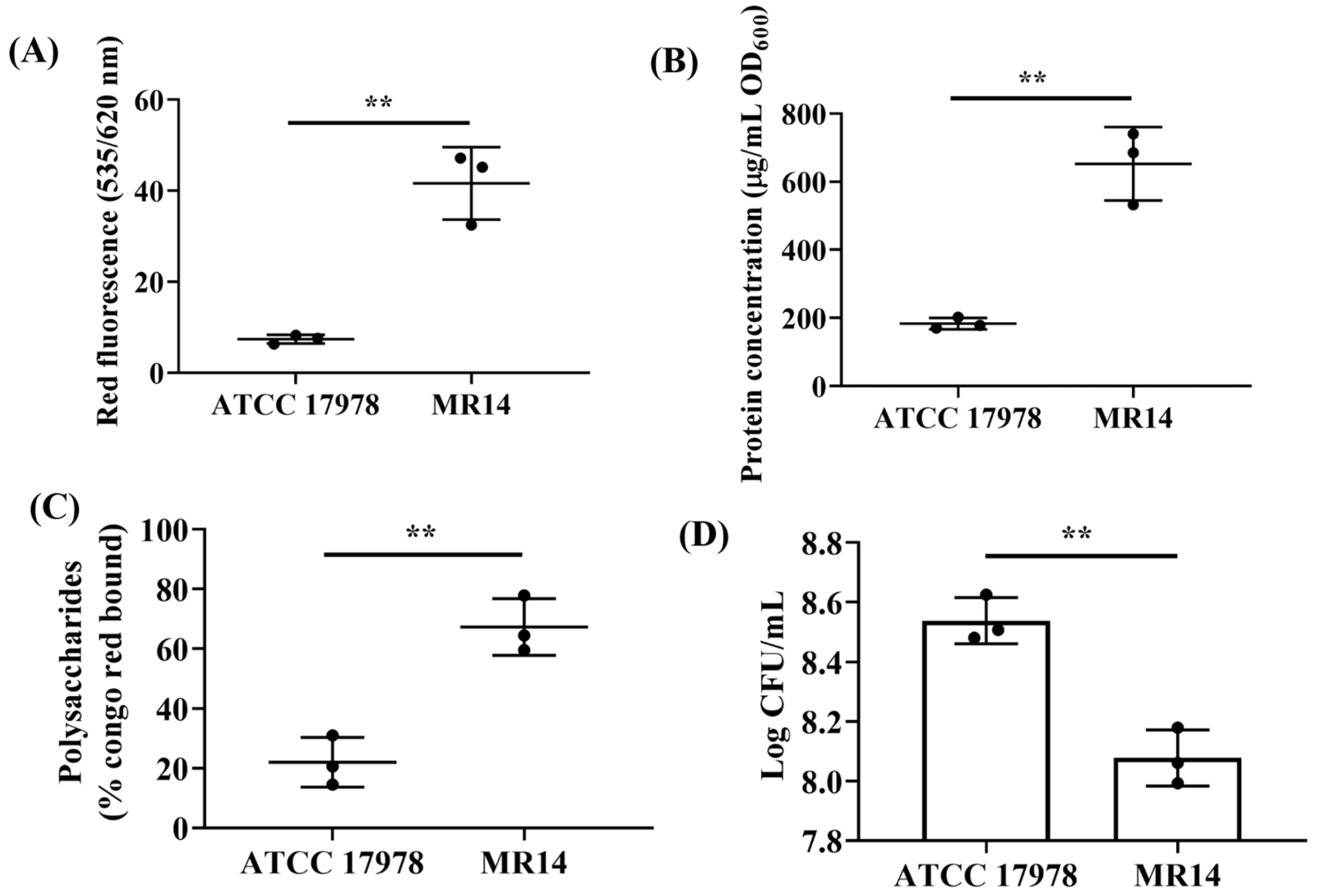
2.7. Quantification of the Biofilm Components
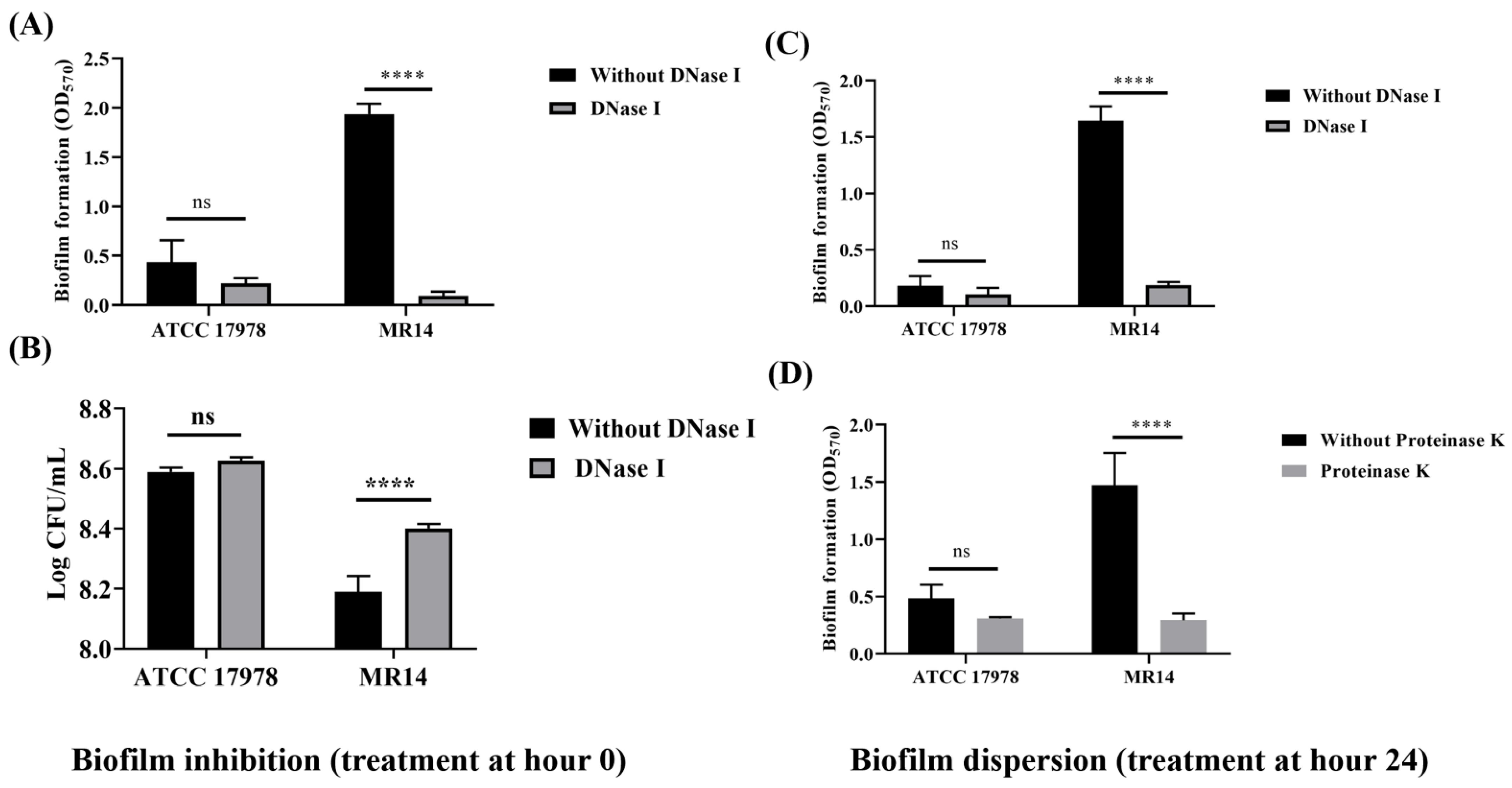
2.8. Biofilm Inhibition and Disruption Assay
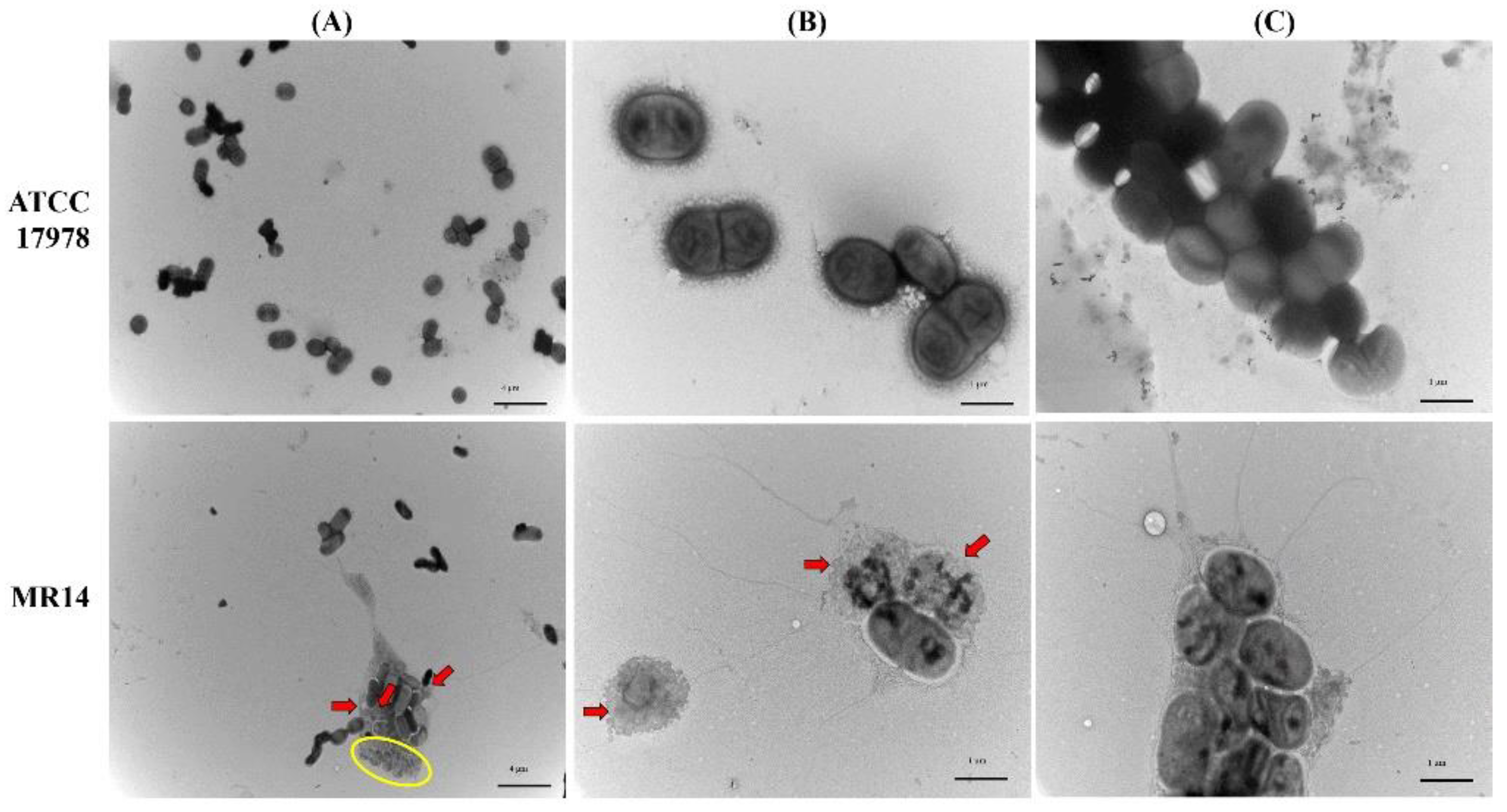
2.9. TEM Analysis of Biofilm and Planktonic Cells
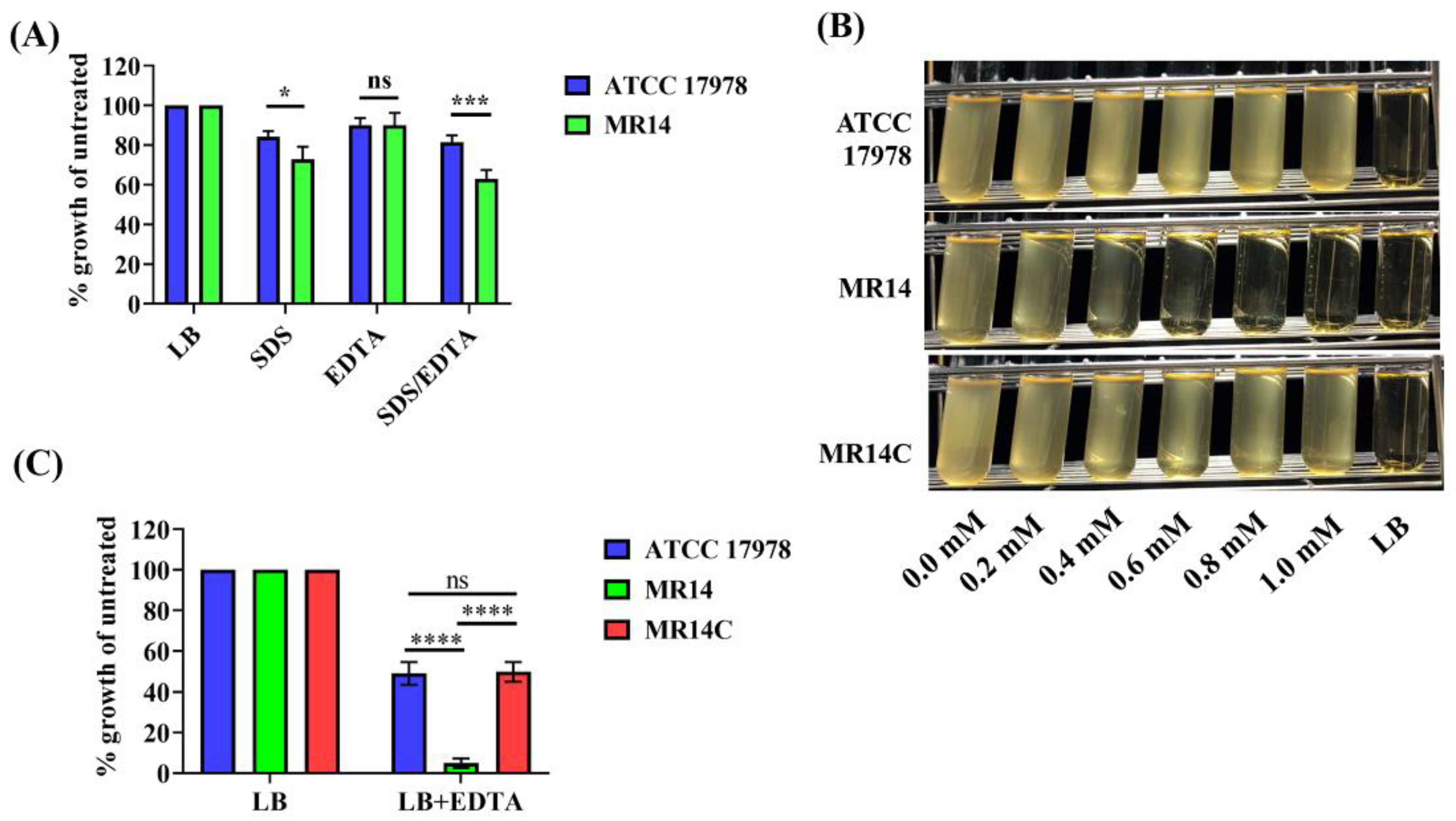
2.10. SDS and EDTA Sensitivity Assay
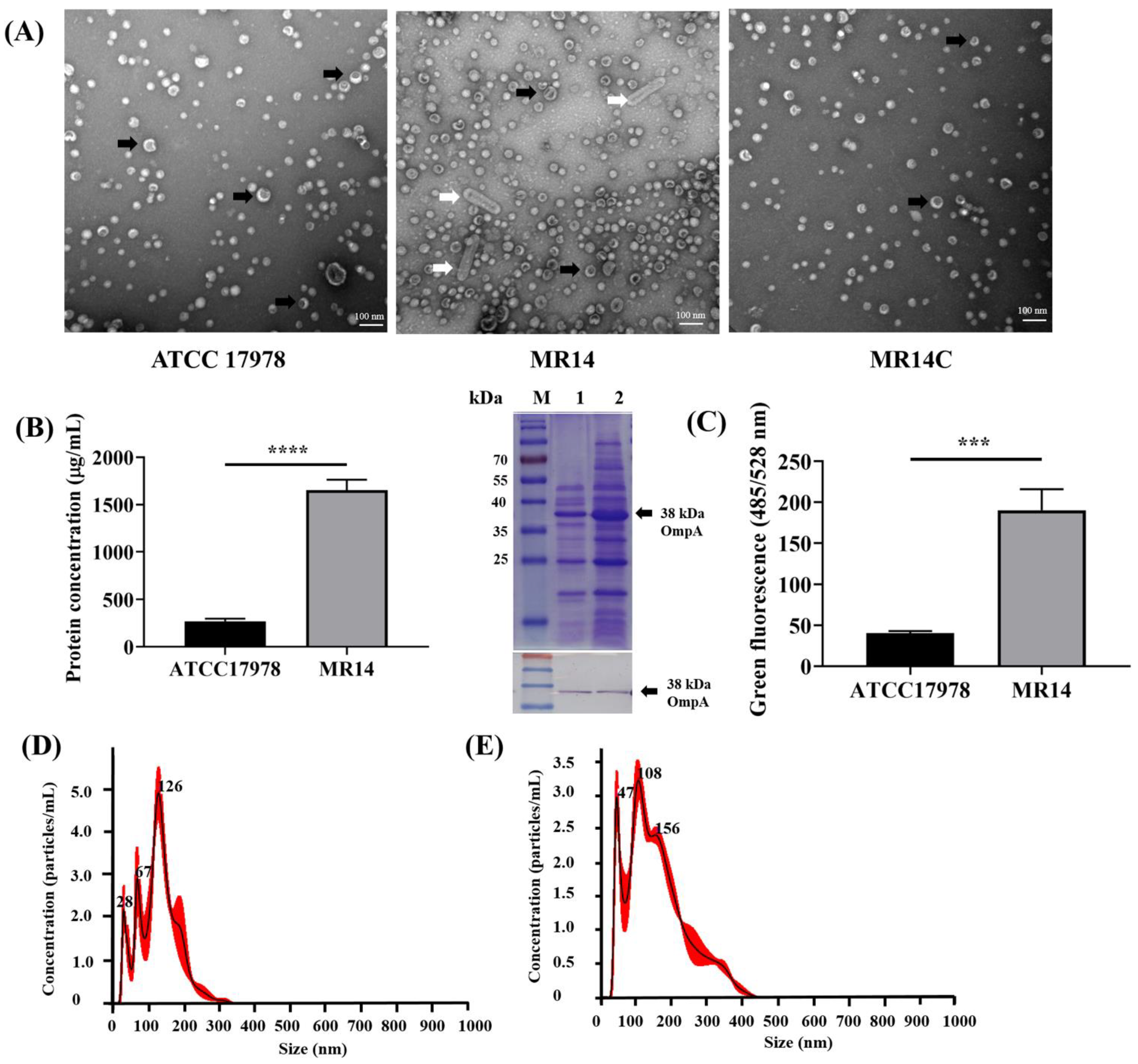
2.11. OMV Isolation, Visualization and Quantification
2.12. Nanoparticle Tracking Analysis of OMVs
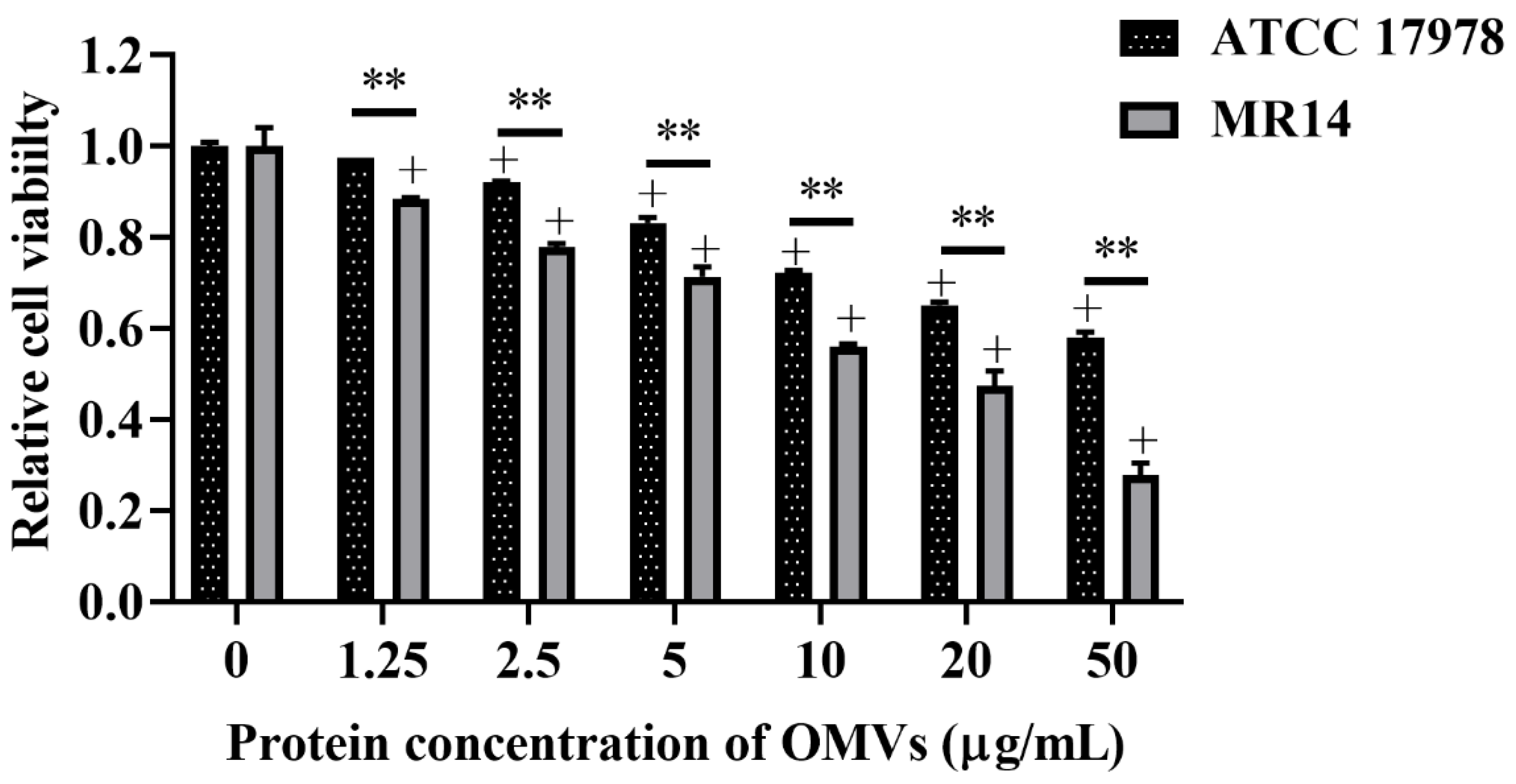
2.13. Cell Viability Test
2.14. Statistical Analyses
3. Results
3.1. Endogenous Ctp Is an Inner Membrane Protein
3.2. Ctp Mutation Promotes CPS Production and Cell Aggregation
3.3. More Biofilm Matrix Surrounded ctp Mutant Due to Cell Content Release by Autolysis
3.4. Microscopic Analysis of Biofilm Cells of ATCC 17978 and MR14
3.5. Ctp Mutant Is Sensitive to SDS and EDTA
3.6. Ctp Mutation Causes Hypervesiculation Phenotype
3.7. OMVs from ctp Mutant Are More Virulent Than Parental Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ctp | Carboxy-terminal processing protease |
| ATCC | American Type Culture Collection |
| eDNA | Extracellular DNA |
| SDS | Sodium dodecyl sulfate |
| EDTA | Ethylenediaminetetraacetic acid |
| OMVs | Outer membrane vesicles |
| PG | Peptidoglycan |
| PBPs | Penicillin-binding proteins |
| OM | Outer membrane |
| PCR | Polymerase chain reaction |
| LB | Luria broth |
| OD | Optical density |
| PBS | Phosphate-buffered saline |
| RT | Room temperature |
| BCA | Bicinchoninic acid |
| CPSs | Capsular polysaccharides |
| CR | Congo Red |
| CV | Crystal violet |
| PI | Propidium iodide |
| CFUs | Colony forming units |
| NTA | Nanoparticle tracking analysis |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| OMT | Outer membrane tubes |
| MVs | Membrane vesicles |
References
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a Successful Pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K. Antimicrobial agents targeting bacterial cell walls and cell membranes. Rev. Sci. Tech. l’OIE 2012, 31, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Angeles, D.M.; Scheffers, D.-J. The Cell Wall of Bacillus subtilis. Curr. Issues Mol. Biol. 2021, 41, 539–596. [Google Scholar] [CrossRef] [PubMed]
- Henderson, T.A.; Dombrosky, P.M.; Young, K.D. Artifactual processing of penicillin-binding proteins 7 and 1b by the OmpT protease of Escherichia coli. J. Bacteriol. 1994, 176, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Abe, N.; Nakakouji, M.; Nishimura, Y.; Horiuchi, K. Overproduction of penicillin-binding protein 7 suppresses thermosensitive growth defect at low osmolarity due to an spr mutation of Escherichia coli. Microb. Drug Resist. 1996, 2, 63–72. [Google Scholar] [CrossRef]
- Hernández, S.B.; Ayala, J.A.; Rico-Pérez, G.; García-Del Portillo, F.; Casadesús, J. Increased bile resistance in Salmonella enterica mutants lacking Prc periplasmic protease. Int. Microbiol. 2013, 16, 87–92. [Google Scholar] [PubMed] [Green Version]
- Wang, C.Y.; Wang, S.W.; Huang, W.C.; Kim, K.S.; Chang, N.S.; Wang, Y.H.; Wu, M.H.; Teng, C.H. Prc contributes to Escherichia coli evasion of classical complement-mediated serum killing. Infect. Immun. 2012, 80, 3399–3409. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Östberg, Y.; Carroll, J.A.; Pinne, M.; Krum, J.G.; Rosa, P.; Bergström, S. Pleiotropic effects of inactivating a carboxyl-terminal protease, CtpA, in Borrelia burgdorferi. J. Bacteriol. 2004, 186, 2074–2084. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, D.; Seo, J.; Rimal, B.; Kim, S.J.; Zhen, S.; Darwin, A.J. A Proteolytic Complex Targets Multiple Cell Wall Hydrolases in Pseudomonas aeruginosa. mBio 2018, 9, 00972-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.-Y.; Som, N.; Wu, C.-Y.; Su, S.-C.; Kuo, Y.-T.; Ke, L.-C.; Ho, M.-R.; Tzeng, S.-R.; Teng, C.-H.; Mengin-Lecreulx, D.; et al. Structural basis of adaptor-mediated protein degradation by the tail-specific PDZ-protease Prc. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Parveen, S.; SaiSree, L.; Reddy, M. Regulated proteolysis of a cross-link–specific peptidoglycan hydrolase contributes to bacterial morphogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 10956–10961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.-C.; Chen, C.-S.; Wang, S.; Hashimoto, M.; Huang, W.-C.; Teng, C.-H. Identification of MltG as a Prc Protease Substrate Whose Dysregulation Contributes to the Conditional Growth Defect of Prc-Deficient Escherichia coli. Front. Microbiol. 2020, 11, 2000. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.; Idem, U.; Dahms, T.E.S. Altered Envelope Structure and Nanomechanical Properties of a C-Terminal Protease A-Deficient Rhizobium leguminosarum. Microorganisms 2020, 8, 1421. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Shehata, M.M.; Mostafa, A.; Teubner, L.; Mahmoud, S.H.; Kandeil, A.; Elshesheny, R.; Boubak, T.A.; Frantz, R.; La Pietra, L.; Pleschka, S. Bacterial outer membrane vesicles (OMVs)-based dual vaccine for influenza a H1N1 virus and MERS-CoV. Vaccines 2019, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urashima, A.; Sanou, A.; Yen, H.; Tobe, T. Enterohaemorrhagic Escherichia coli produces outer membrane vesicles as an active defence system against antimicrobial peptide LL-37. Cell. Microbiol. 2017, 19, e12758. [Google Scholar] [CrossRef] [Green Version]
- Bauwens, A.; Kunsmann, L.; Karch, H.; Mellmann, A.; Bielaszewska, M. Antibiotic-Mediated Modulations of Outer Membrane Vesicles in Enterohemorrhagic Escherichia coli O104:H4 and O157:H7. Antimicrob. Agents Chemother. 2017, 61, e00937-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Robles, T.; Dillard, R.S.; Cairns, L.S.; Silva-Valenzuela, C.A.; Housman, M.; Ali, A.; Wright, E.R.; Camilli, A. Vibrio cholerae Outer Membrane Vesicles Inhibit Bacteriophage Infection. J. Bacteriol. 2018, 200, 00792-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamalier, J.P.; Silva, T.P.; Zarantonello, V.; Dias, F.F.; Melo, R.C. Increased production of outer membrane vesicles by cultured freshwater bacteria in response to ultraviolet radiation. Microbiol. Res. 2017, 194, 38–46. [Google Scholar] [CrossRef]
- Heredia, R.M.; Boeris, P.S.; Liffourrena, A.S.; Bergero, M.F.; López, G.A.; Lucchesi, G.I. Release of outer membrane vesicles in Pseudomonas putida as a response to stress caused by cationic surfactants. Microbiology 2016, 162, 813–822. [Google Scholar] [CrossRef]
- Bitto, N.J.; Chapman, R.; Pidot, S.; Costin, A.; Lo, C.; Choi, J.; D’Cruze, T.; Reynolds, E.C.; Dashper, S.G.; Turnbull, L.; et al. Bacterial membrane vesicles transport their DNA cargo into host cells. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Haurat, M.F.; Elhenawy, W.; Feldman, M.F. Prokaryotic membrane vesicles: New insights on biogenesis and biological roles. Biol. Chem. 2015, 396, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Zingl, F.; Kohl, P.; Cakar, F.; Leitner, D.R.; Mitterer, F.; Bonnington, K.E.; Rechberger, G.N.; Kuehn, M.; Guan, Z.; Reidl, J.; et al. Outer Membrane Vesiculation Facilitates Surface Exchange and In Vivo Adaptation of Vibrio cholerae. Cell Host Microb. 2020, 27, 225–237.e8. [Google Scholar] [CrossRef] [Green Version]
- Banzhaf, M.; Yau, H.C.; Verheul, J.; Lodge, A.; Kritikos, G.; Mateus, A.; Cordier, B.; Hov, A.K.; Stein, F.; Wartel, M.; et al. Outer membrane lipoprotein NlpI scaffolds peptidoglycan hydrolases within multi-enzyme complexes in Escherichia coli. EMBO J. 2020, 39, e102246. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. Microbiologyopen 2015, 4, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Torres, V.; Maeda, T.; Wood, T.K. Global regulator H-NS and lipoprotein NlpI influence production of extracellular DNA in Escherichia coli. Biochem. Biophys. Res. Commun. 2010, 401, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Rouf, S.F.; Ahmad, I.; Anwar, N.; Vodnala, S.K.; Kader, A.; Romling, U.; Rhen, M. Opposing Contributions of Polynucleotide Phosphorylase and the Membrane Protein NlpI to Biofilm Formation by Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2010, 193, 580–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2020, 10, 3. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, X.; Jiang, Q.; Xue, T.; Sun, B. Pfs promotes autolysis-dependent release of eDNA and biofilm formation in Staphylococcus aureus. Med. Microbiol. Immunol. 2014, 204, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.C.; Thurlow, L.R.; Boyle, D.; Hancock, L.E. Regulation of Autolysis-Dependent Extracellular DNA Release by Enterococcus faecalis Extracellular Proteases Influences Biofilm Development. J. Bacteriol. 2008, 190, 5690–5698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigurlásdóttir, S.; Wassing, G.M.; Zuo, F.; Arts, M.; Jonsson, A.-B. Deletion of D-Lactate Dehydrogenase A in Neisseria meningitidis Promotes Biofilm Formation Through Increased Autolysis and Extracellular DNA Release. Front. Microbiol. 2019, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; You, R.-I.; Lin, M.-D.; Lin, N.-T. Mutation of the Carboxy-Terminal Processing Protease in Acinetobacter baumannii Affects Motility, Leads to Loss of Membrane Integrity, and Reduces Virulence. Pathogens 2020, 9, 322. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.D.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis. Front. Microbiol. 2021, 12, 557902. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; van der Veen, S. Role of outer membrane vesicles in bacterial physiology and host cell interactions. Infect. Microb. Dis. 2020, 2, 3–9. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.-N.; Jia, H.-B.; Wang, P.; Dong, J.-F.; Deng, J.; Lu, F.-M.; Zou, Q.-H. The type VI secretion system protein AsaA in Acinetobacter baumannii is a periplasmic protein physically interacting with TssM and required for T6SS assembly. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Cian, M.B.; Giordano, N.P.; Mettlach, J.A.; Minor, K.E.; Dalebroux, Z.D. Separation of the Cell Envelope for Gram-negative Bacteria into Inner and Outer Membrane Fractions with Technical Adjustments for Acinetobacter baumannii. J. Vis. Exp. 2020, 10, e60517. [Google Scholar] [CrossRef]
- Iwashkiw, J.A.; Seper, A.; Weber, B.S.; Scott, N.; Vinogradov, E.; Stratilo, C.; Reiz, B.; Cordwell, S.; Whittal, R.; Schild, S.; et al. Identification of a General O-linked Protein Glycosylation System in Acinetobacter baumannii and Its Role in Virulence and Biofilm Formation. PLoS Pathog. 2012, 8, e1002758. [Google Scholar] [CrossRef]
- Yang, Z. Characterization of the Acinetobacter Baumannii Glycoprotein A1S_3744 and Effects of the Immune protein P64KN on the Immunogenicity of the Identified Protective Antigens. Master’s Thesis, National Chung Hsing University, Taichung City, Taiwan, 2015. [Google Scholar]
- Bouffartigues, E.; Moscoso, J.A.; Duchesne, R.; Rosay, T.; Fito-Boncompte, L.; Gicquel, G.; Maillot, O.; Bénard, M.; Bazire, A.; Brenner-Weiss, G.; et al. The absence of the Pseudomonas aeruginosa OprF protein leads to increased biofilm formation through variation in c-di-GMP level. Front. Microbiol. 2015, 6, 630. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Majumder, R.; Rather, I.A.; Kim, K. Extraction, isolation and purification of exopolysaccharide from lactic acid bacteria using ethanol precipitation method. Bangladesh J. Pharmacol. 2016, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Laurentin, A.; Edwards, C.A. A microtiter modification of the anthrone-sulfuric acid colorimetric assay for glucose-based carbohydrates. Anal. Biochem. 2003, 315, 143–145. [Google Scholar] [CrossRef]
- Selasi, G.N.; Nicholas, A.; Jeong, K.Y.; Na, S.H.; Kwon, H.I.; Kim, Y.J.; Heo, S.T.; Oh, M.H.; Lee, J.C. Differences in Biofilm Mass, Expression of Biofilm-Associated Genes, and Resistance to Desiccation between Epidemic and Sporadic Clones of Carbapenem-Resistant Acinetobacter baumannii Sequence Type 191. PLoS ONE 2016, 11, e0162576. [Google Scholar] [CrossRef]
- Desai, S.; Sanghrajka, K.; Gajjar, D. High Adhesion and Increased Cell Death Contribute to Strong Biofilm Formation in Klebsiella pneumoniae. Pathogens 2019, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Kimyon, Ö.; Das, T.; Ibugo, A.I.; Kutty, S.K.; Ho, K.K.; Tebben, J.; Kumar, N.; Manefield, M. Serratia Secondary Metabolite Prodigiosin Inhibits Pseudomonas aeruginosa Biofilm Development by Producing Reactive Oxygen Species that Damage Biological Molecules. Front. Microbiol. 2016, 7, 972. [Google Scholar] [CrossRef]
- Shi, L.; Wu, Y.; Yang, C.; Ma, Y.; Zhang, Q.-Z.; Huang, W.; Zhu, X.-Y.; Yan, Y.-J.; Wang, J.-X.; Zhu, T.; et al. Effect of nicotine on Staphylococcus aureus biofilm formation and virulence factors. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Lonergan, Z.; Nairn, B.L.; Wang, J.; Hsu, Y.-P.; Hesse, L.; Beavers, W.N.; Chazin, W.J.; Trinidad, J.C.; VanNieuwenhze, M.S.; Giedroc, D.P.; et al. An Acinetobacter baumannii, Zinc-Regulated Peptidase Maintains Cell Wall Integrity during Immune-Mediated Nutrient Sequestration. Cell Rep. 2019, 26, 2009–2018.e6. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.-T.; Kuo, S.-C.; Chiang, M.-H.; Lee, Y.-T.; Sung, W.-C.; Chen, Y.-H.; Chen, T.-L.; Fung, C.-P. Acinetobacter baumannii Extracellular OXA-58 Is Primarily and Selectively Released via Outer Membrane Vesicles after Sec-Dependent Periplasmic Translocation. Antimicrob. Agents Chemother. 2015, 59, 7346–7354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasperini, G.; Biagini, M.; Arato, V.; Gianfaldoni, C.; Vadi, A.; Norais, N.; Bensi, G.; Delany, I.; Pizza, M.; Aricò, B.; et al. Outer Membrane Vesicles (OMV)-based and Proteomics-driven Antigen Selection Identifies Novel Factors Contributing to Bordetella pertussis Adhesion to Epithelial Cells. Mol. Cell. Proteom. 2018, 17, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, M.H.; Kim, S.I.; Son, J.H.; Kim, S.; Lee, Y.C.; Shin, M.; Oh, M.H.; Lee, J.C. The sensor kinase BfmS controls production of outer membrane vesicles in Acinetobacter baumannii. BMC Microbiol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hoge, R.; Laschinski, M.; Jaeger, K.-E.; Wilhelm, S.; Rosenau, F. The subcellular localization of a C-terminal processing protease in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2011, 316, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K. Inactivation of cysL inhibits biofilm formation by activating the disulfide stress regulator Spx in Bacillus subtilis. J. Bacteriol. 2019, 201, e00712-18. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Shan, R.; Chen, G.; Liu, L. Linking flow velocity-regulated EPS production with early-stage biofilm formation in drinking water distribution systems. Water Supply 2020, 20, 1253–1263. [Google Scholar] [CrossRef]
- Nagakubo, T.; Nomura, N.; Toyofuku, M. Cracking Open Bacterial Membrane Vesicles. Front. Microbiol. 2019, 10, 3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, A.M.; Srikumar, T.; Silhavy, T.J. Cyclic Enterobacterial Common Antigen Maintains the Outer Membrane Permeability Barrier of Escherichia coli in a Manner Controlled by YhdP. mBio 2018, 9, 01321-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaig, W.D.; Koller, A.; Thanassi, D.G. Production of Outer Membrane Vesicles and Outer Membrane Tubes by Francisella novicida. J. Bacteriol. 2012, 195, 1120–1132. [Google Scholar] [CrossRef]
- Lin, M.-F.; Tsai, P.-W.; Chen, J.-Y.; Lin, Y.-Y.; Lan, C.-Y. OmpA Binding Mediates the Effect of Antimicrobial Peptide LL-37 on Acinetobacter baumannii. PLoS ONE 2015, 10, e0141107. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Yamamoto, Y.; Higashitani, A.; Suzuki, H.; Nishimura, Y. Cloning, mapping, and characterization of the Escherichia coli prc gene, which is involved in C-terminal processing of penicillin-binding protein 3. J. Bacteriol. 1991, 173, 4799–4813. [Google Scholar] [CrossRef] [Green Version]
- Carroll, R.K.; Rivera, F.E.; Cavaco, C.K.; Johnson, G.M.; Martin, D.; Shaw, L.N. The lone S41 family C-terminal processing protease in Staphylococcus aureus is localized to the cell wall and contributes to virulence. Microbiology 2014, 160, 1737–1748. [Google Scholar] [CrossRef] [Green Version]
- Regina, V.R.; Lokanathan, A.R.; Modrzyński, J.J.; Sutherland, D.S.; Meyer, R.L. Surface Physicochemistry and Ionic Strength Affects eDNA’s Role in Bacterial Adhesion to Abiotic Surfaces. PLoS ONE 2014, 9, e105033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, G.F.D.M.; Tognim, M.C.B.; Cardoso, C.L.; Carrara-Marrone, F.E.; Garcia, L.B. Preliminary evaluation of adherence on abiotic and cellular surfaces of Acinetobacter baumannii strains isolated from catheter tips. Braz. J. Infect. Dis. 2006, 10, 346–351. [Google Scholar] [CrossRef] [Green Version]
- Skerniškytė, J.; Krasauskas, R.; Péchoux, C.; Kulakauskas, S.; Armalytė, J.; Sužiedėlienė, E. Surface-Related Features and Virulence Among Acinetobacter baumannii Clinical Isolates Belonging to International Clones I and II. Front. Microbiol. 2019, 9, 3116. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, X.; Qin, J.; Cheng, S.; Yeo, W.-S.; He, L.; Ma, X.; Liu, X.; Li, M.; Bae, T. The ATP-Dependent Protease ClpP Inhibits Biofilm Formation by Regulating Agr and Cell Wall Hydrolase Sle1 in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2017, 7, 181. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Signo, K.S.L.; Vanderlinde, E.M.; Yost, C.; Dahms, T.E.S. Atomic force microscopy of a ctpA mutant in Rhizobium leguminosarum reveals surface defects linking CtpA function to biofilm formation. Microbiology 2011, 157, 3049–3058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okshevsky, M.; Meyer, R.L. The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit. Rev. Microbiol. 2013, 41, 341–352. [Google Scholar] [CrossRef]
- Schooling, S.R.; Hubley, A.; Beveridge, T.J. Interactions of DNA with Biofilm-Derived Membrane Vesicles. J. Bacteriol. 2009, 191, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, R.; Celia, C.; Mincione, G.; Stringaro, A.; Di Marzio, L.; Colone, M.; Di Marcantonio, M.C.; Savino, L.; Puca, V.; Santoliquido, R.; et al. Detection and Physicochemical Characterization of Membrane Vesicles (MVs) of Lactobacillus reuteri DSM 17938. Front. Microbiol. 2017, 8, 1040. [Google Scholar] [CrossRef]
- Baumgarten, T.; Sperling, S.; Seifert, J.; Von Bergen, M.; Steiniger, F.; Wick, L.Y.; Heipieper, H.J. Membrane Vesicle Formation as a Multiple-Stress Response Mechanism Enhances Pseudomonas putida DOT-T1E Cell Surface Hydrophobicity and Biofilm Formation. Appl. Environ. Microbiol. 2012, 78, 6217–6224. [Google Scholar] [CrossRef] [Green Version]
- Weski, J.; Ehrmann, M. Genetic Analysis of 15 Protein Folding Factors and Proteases of the Escherichia coli Cell Envelope. J. Bacteriol. 2012, 194, 3225–3233. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Kim, H.J.; Oh, M.H.; Kim, S.Y.; Kim, M.H.; Son, J.H.; Kim, S.I.; Shin, M.; Lee, Y.C.; Lee, J.C. The role of Zur-regulated lipoprotein A in bacterial morphology, antimicrobial susceptibility, and production of outer membrane vesicles in Acinetobacter baumannii. BMC Microbiol. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Shahryari, S.; Talaee, M.; Haghbeen, K.; Adrian, L.; Vali, H.; Zahiri, H.S.; Noghabi, K.A. New Provisional Function of OmpA from Acinetobacter sp. Strain SA01 Based on Environmental Challenges. mSystems 2021, 6, 01175-20. [Google Scholar] [CrossRef]
- Skerniškytė, J.; Karazijaitė, E.; Lučiūnaitė, A.; Sužiedėlienė, E. OmpA protein-deficient Acinetobacter baumannii outer membrane vesicles trigger reduced inflammatory response. Pathogens 2021, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Nie, D.; Hu, Y.; Chen, Z.; Li, M.; Hou, Z.; Luo, X.; Mao, X.; Xue, X. Outer membrane protein A (OmpA) as a potential therapeutic target for Acinetobacter baumannii infection. J. Biomed. Sci. 2020, 27, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.K.; Choi, C.H.; Oh, M.H. Zur-regulated lipoprotein A contributes to the fitness of Acinetobacter baumannii. J. Microbiol. 2020, 58, 67–77. [Google Scholar] [CrossRef]
- Huang, W.-C.; Lin, C.-Y.; Hashimoto, M.; Wu, J.-J.; Wang, M.-C.; Lin, W.-H.; Chen, C.-S.; Teng, C.-H. The role of the bacterial protease Prc in the uropathogenesis of extraintestinal pathogenic Escherichia coli. J. Biomed. Sci. 2020, 27, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, R.; You, R.-I.; Chang, C.-H.; Yang, C.-Y.; Lin, N.-T. Carboxy-Terminal Processing Protease Controls Production of Outer Membrane Vesicles and Biofilm in Acinetobacter baumannii. Microorganisms 2021, 9, 1336. https://doi.org/10.3390/microorganisms9061336
Roy R, You R-I, Chang C-H, Yang C-Y, Lin N-T. Carboxy-Terminal Processing Protease Controls Production of Outer Membrane Vesicles and Biofilm in Acinetobacter baumannii. Microorganisms. 2021; 9(6):1336. https://doi.org/10.3390/microorganisms9061336
Chicago/Turabian StyleRoy, Rakesh, Ren-In You, Chan-Hua Chang, Chiou-Ying Yang, and Nien-Tsung Lin. 2021. "Carboxy-Terminal Processing Protease Controls Production of Outer Membrane Vesicles and Biofilm in Acinetobacter baumannii" Microorganisms 9, no. 6: 1336. https://doi.org/10.3390/microorganisms9061336