
Comparative Exoproteome Analysis of Streptococcus suis Human Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions and the Preparation of Exoproteomes
2.2. Sample Preparation for Proteomic Analysis
2.3. LC-MS/MS Analysis and Protein Identification through a Database Search
2.4. Computational Analysis of the Protein Sequences
2.5. Data and Statistical Analysis
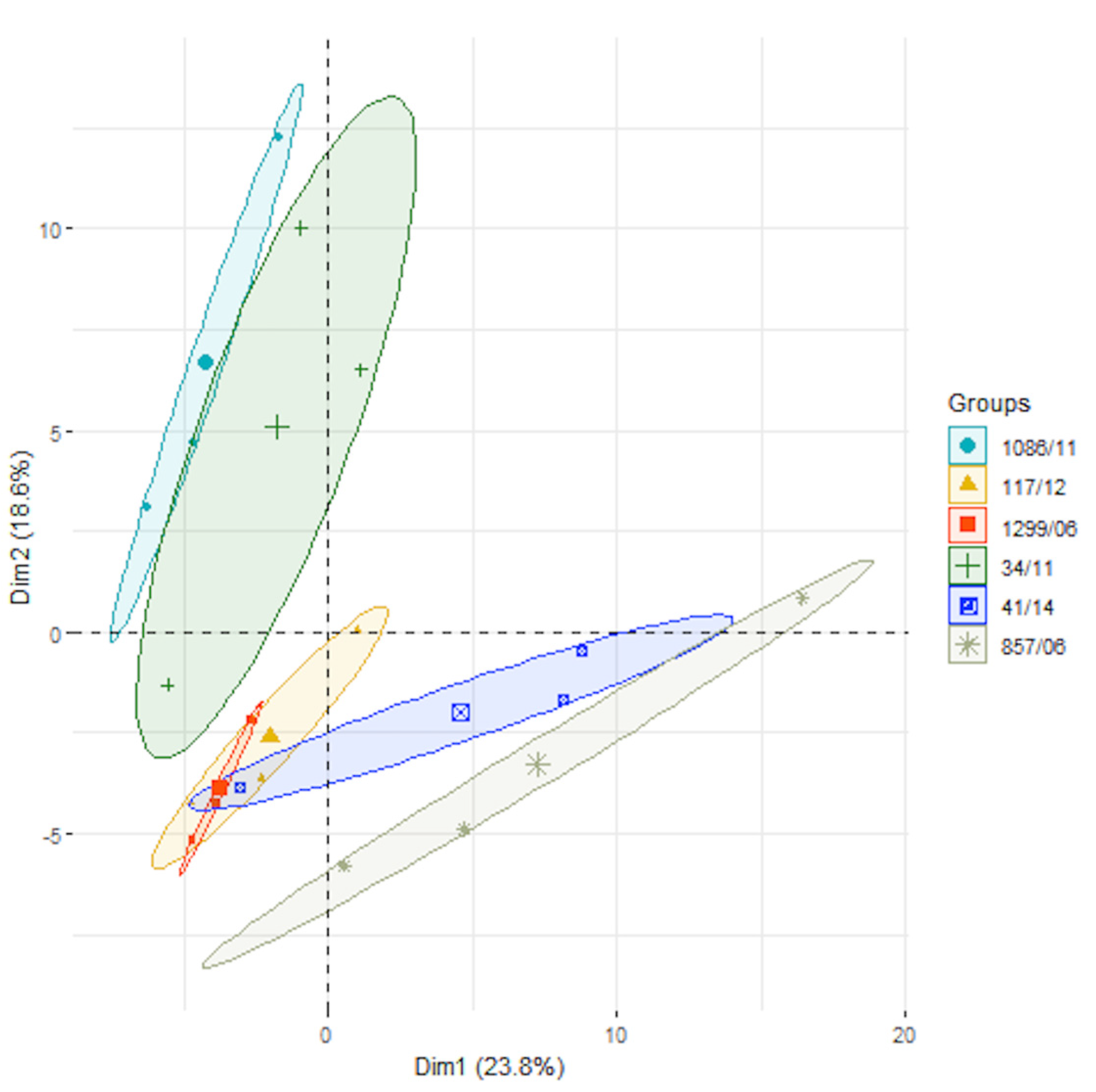
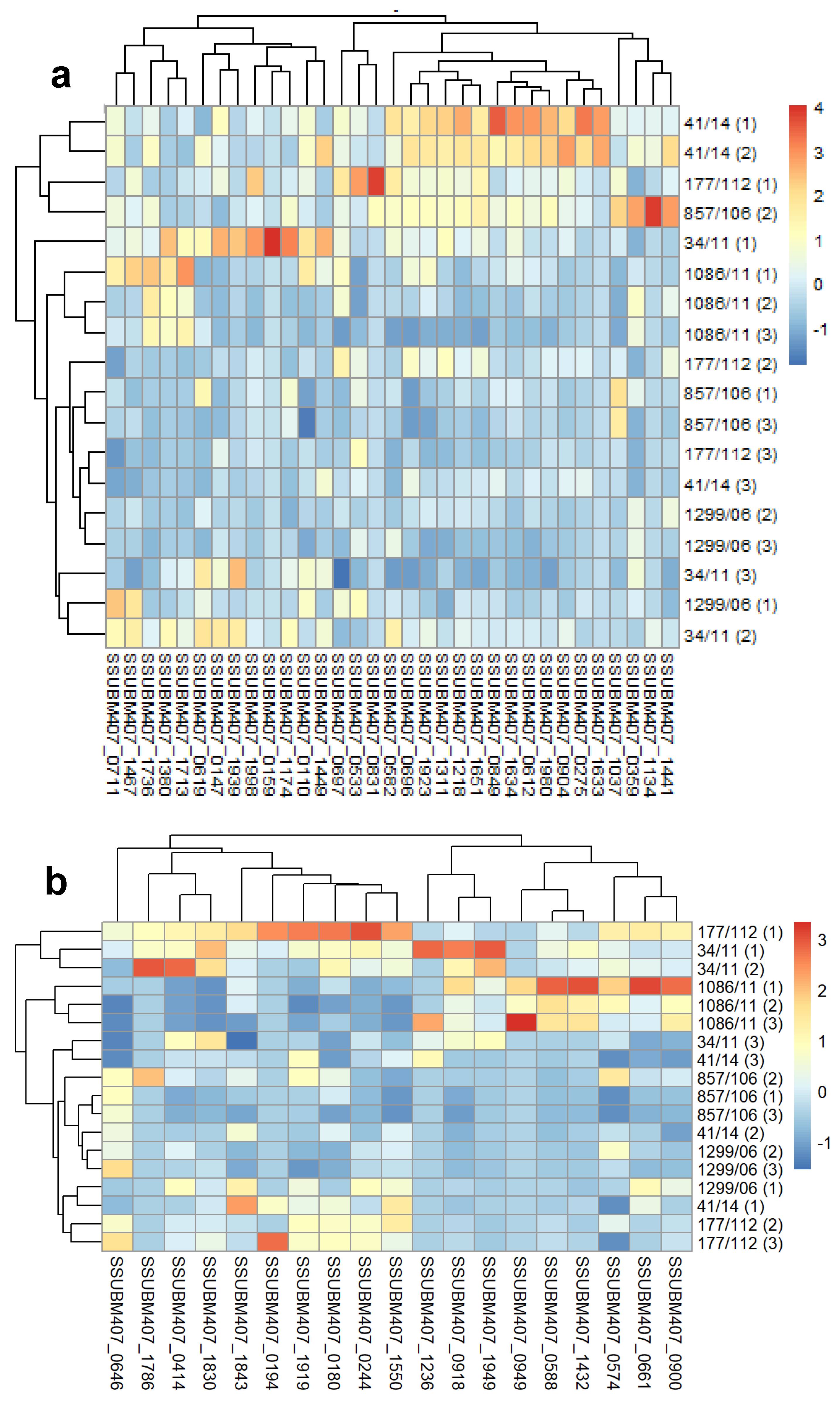
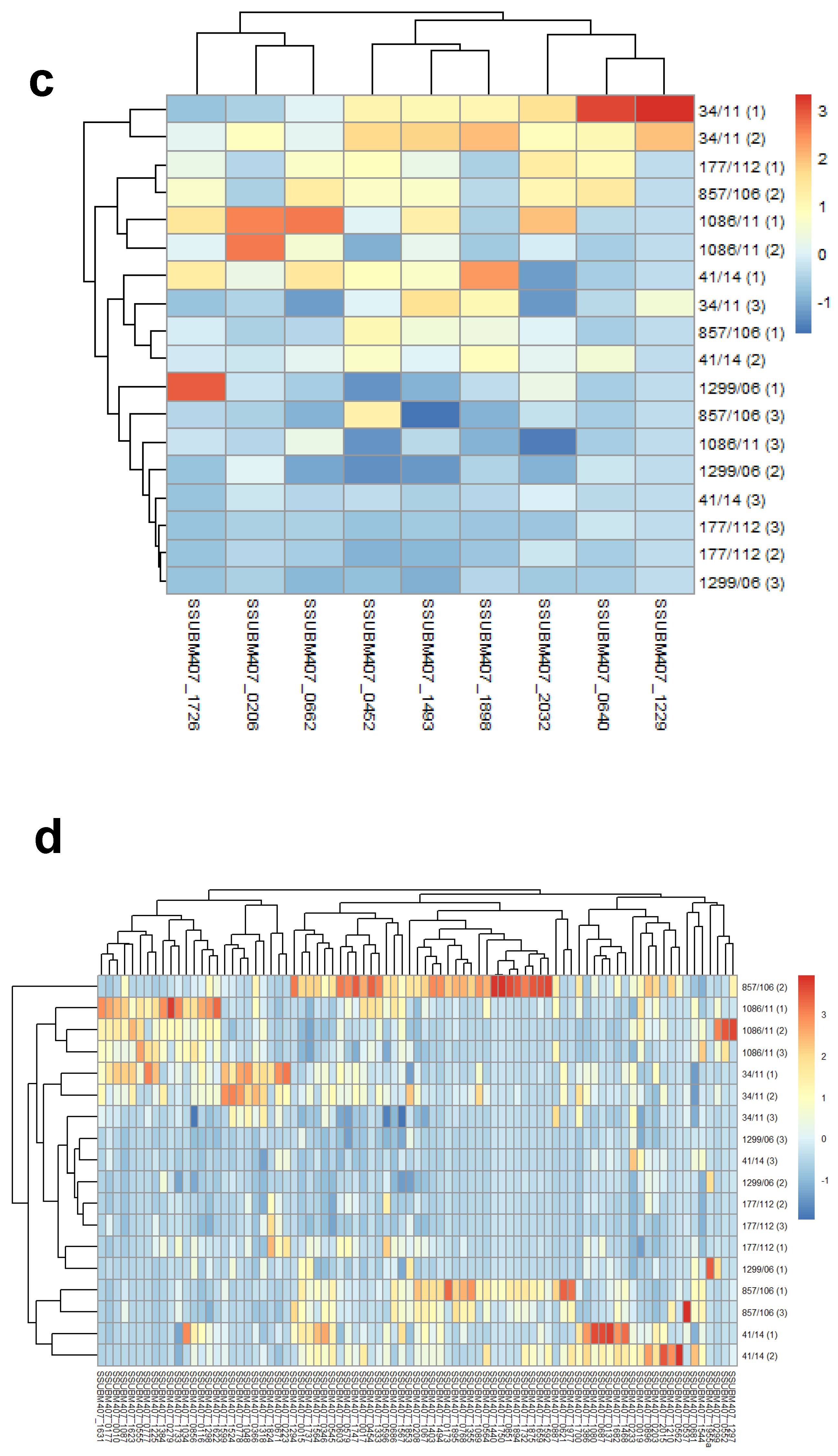
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vötsch, D.; Willenborg, M.; Weldearegay, Y.B.; Valentin-Weigand, P. Streptococcus Suis-The “Two Faces” of a Pathobiont in the Porcine Respiratory Tract. Front. Microbiol. 2018, 9, 480. [Google Scholar] [CrossRef]
- Weinert, L.A.; Chaudhuri, R.R.; Wang, J.; Peters, S.E.; Corander, J.; Jombart, T.; Baig, A.; Howell, K.J.; Vehkala, M.; Valimaki, N.; et al. Genomic Signatures of Human and Animal Disease in the Zoonotic Pathogen Streptococcus Suis. Nat. Commun. 2015, 6, 6740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerdsin, A.; Takeuchi, D.; Nuangmek, A.; Akeda, Y.; Gottschalk, M.; Oishi, K. Genotypic Comparison between Streptococcus Suis Isolated from Pigs and Humans in Thailand. Pathogens 2020, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nghia, H.D.; Tu Le, T.P.; Wolbers, M.; Thai, C.Q.; Hoang, N.V.; Nga, T.V.; Thao Le, T.P.; Phu, N.H.; Chau, T.T.; Sinh, D.X.; et al. Risk Factors of Streptococcus Suis Infection in Vietnam. A Case-Control Study. PLoS ONE 2011, 6, e17604. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus Suis Virulence Factors: Are They All Really Critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus Suis: An Emerging Zoonotic Pathogen. Lancet Infect Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Segura, M. Streptococcus Suis Vaccines: Candidate Antigens and Progress. Expert Rev. Vaccines 2015, 14, 1587–1608. [Google Scholar] [CrossRef]
- Olaya-Abril, A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Rodríguez-Ortega, M.J. Surfomics: Shaving Live Organisms for a Fast Proteomic Identification of Surface Proteins. J. Proteom. 2014, 97, 164–176. [Google Scholar] [CrossRef]
- Gómez-Gascón, L.; Luque, I.; Olaya-Abril, A.; Jiménez-Munguía, I.; Orbegozo-Medina, R.A.; Peralbo, E.; Tarradas, C.; Rodríguez-Ortega, M.J. Exploring the Pan-Surfome of Streptococcus Suis: Looking for Common Protein Antigens. J. Proteom. 2012, 75, 5654–5666. [Google Scholar] [CrossRef]
- Prados de la Torre, E.; Rodríguez-Franco, A.; Rodríguez-Ortega, M.J. Proteomic and Bioinformatic Analysis of Streptococcus Suis Human Isolates: Combined Prediction of Potential Vaccine Candidates. Vaccines 2020, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ortega, M.J.; Norais, N.; Bensi, G.; Liberatori, S.; Capo, S.; Mora, M.; Scarselli, M.; Doro, F.; Ferrari, G.; Garaguso, I.; et al. Characterization and Identification of Vaccine Candidate Proteins through Analysis of the Group A Streptococcus Surface Proteome. Nat. Biotechnol. 2006, 24, 191–197. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J. “Shaving” Live Bacterial Cells with Proteases for Proteomic Analysis of Surface Proteins. Methods Mol. Biol. 2018, 1722, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Zhu, L.; Yuan, Y.; Zhang, W.; Li, W.; Wang, J.; Zheng, Y.; Wei, K.; Cao, W.; Wang, H.; et al. Identification and Characterization of Novel Immunogenic Proteins of Streptococcus Suis Serotype 2. J. Proteome Res. 2008, 7, 4132–4142. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, W.; Lu, C. Comparative Proteome Analysis of Secreted Proteins of Streptococcus Suis Serotype 9 Isolates from Diseased and Healthy Pigs. Microb. Pathog. 2008, 45, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lu, C.P. Immunoproteomics of Extracellular Proteins of Chinese Virulent Strains OfStreptococcus Suis Type 2. PROTEOMICS 2007, 7, 4468–4476. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- UniProt. Available online: https://www.uniprot.org/proteomes/UP000009077 (accessed on 17 July 2019).
- Zhou, M.; Boekhorst, J.; Francke, C.; Siezen, R.J. LocateP: Genome-Scale Subcellular-Location Predictor for Bacterial Proteins. BMC Bioinform. 2008, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Juncker, A.S.; Willenbrock, H.; Von Heijne, G.; Brunak, S.; Nielsen, H.; Krogh, A. Prediction of Lipoprotein Signal Peptides in Gram-Negative Bacteria. Protein Sci. 2003, 12, 1652–1662. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A Server for Prediction of Protective Antigens, Tumour Antigens and Subunit Vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Aranda, J.; Garrido, M.E.; Cortés, P.; Llagostera, M.; Barbé, J. Analysis of the Protective Capacity of Three Streptococcus Suis Proteins Induced under Divalent-Cation-Limited Conditions. Infect. Immun. 2008, 76, 1590–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Cheng, G.; Wang, C.; Pan, X.; Cong, Y.; Pan, Q.; Wang, J.; Zheng, F.; Hu, F.; Tang, J. Identification and Experimental Verification of Protective Antigens against Streptococcus Suis Serotype 2 Based on Genome Sequence Analysis. Curr. Microbiol. 2009, 58, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; Deng, L.; Zheng, C.; Yuan, F.; Chen, H.; Bei, W.; Li, J. Evaluation of the Protective Efficacy of Four Novel Identified Membrane Associated Proteins of Streptococcus Suis Serotype 2. Vaccine 2015, 33, 2254–2260. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.S.; Rosa, I.I.R.; Novaes, E.; de Oliveira, L.S.; Baeza, L.C.; Borges, C.L.; Marlinghaus, L.; de Soares, C.M.A.; Giambiagi-deMarval, M.; Parente-Rocha, J.A. The Exoproteome Profiles of Three Staphylococcus Saprophyticus Strains Reveal Diversity in Protein Secretion Contents. Microbiol. Res. 2018, 216, 85–96. [Google Scholar] [CrossRef]
- Monteiro, R.; Hébraud, M.; Chafsey, I.; Chambon, C.; Viala, D.; Torres, C.; Poeta, P.; Igrejas, G. Surfaceome and Exoproteome of a Clinical Sequence Type 398 Methicillin Resistant Staphylococcus Aureus Strain. Biochem. Biophys. Rep. 2015, 3, 7–13. [Google Scholar] [CrossRef]
- Monteiro, R.; Chafsey, I.; Ageorges, V.; Leroy, S.; Chambon, C.; Hébraud, M.; Livrelli, V.; Pizza, M.; Pezzicoli, A.; Desvaux, M. The Secretome Landscape of Escherichia Coli O157:H7: Deciphering the Cell-Surface, Outer Membrane Vesicle and Extracellular Subproteomes. J. Proteom. 2021, 232, 104025. [Google Scholar] [CrossRef]
- Cavanagh, J.P.; Pain, M.; Askarian, F.; Bruun, J.-A.; Urbarova, I.; Wai, S.N.; Schmidt, F.; Johannessen, M. Comparative Exoproteome Profiling of an Invasive and a Commensal Staphylococcus Haemolyticus Isolate. J. Proteom. 2019, 197, 106–114. [Google Scholar] [CrossRef]
- Gómez-Gascón, L.; Luque, I.; Tarradas, C.; Olaya-Abril, A.; Astorga, R.J.; Huerta, B.; Rodríguez-Ortega, M.J. Comparative Immunosecretome Analysis of Prevalent Streptococcus Suis Serotypes. Comp. Immunol. Microbiol. Infect. Dis. 2018, 57, 55–61. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J.; Luque, I.; Tarradas, C.; Bárcena, J.A. Overcoming Function Annotation Errors in the Gram-Positive Pathogen Streptococcus Suis by a Proteomics-Driven Approach. BMC Genom. 2008, 9, 588. [Google Scholar] [CrossRef] [Green Version]
- Olaya-Abril, A.; Prados-Rosales, R.; McConnell, M.J.; Martín-Peña, R.; González-Reyes, J.A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Fernández, J.; Luque-García, J.L.; García-Lidón, C.; et al. Characterization of Protective Extracellular Membrane-Derived Vesicles Produced by Streptococcus Pneumoniae. J. Proteom. 2014, 106, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Grenier, D. Isolation, Characterization and Biological Properties of Membrane Vesicles Produced by the Swine Pathogen Streptococcus Suis. PLoS ONE 2015, 10, e0130528. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein Category a | #b Identified Proteins | #b Predicted Proteins in S. suis BM407 Genome | Identified/Predicted (%) |
|---|---|---|---|
| Lipoprotein | 33 | 40 | 82.5 |
| Cell wall | 19 | 20 | 95 |
| Secreted | 9 | 18 | 50 |
| Membrane | 83 | 479 | 17.3 |
| Cytoplasmic | 680 | 1375 | 49.4 |
| Total | 824 | 1932 | 42.7 |
| Protein Locus | Protein Annotation | VaxiJen Score a | # Strains b |
|---|---|---|---|
| SSUBM407_0206 | Putative exported protein | 0.6462 | 5 |
| SSUBM407_0452 | Putative exported protein | 0.6648 | 6 |
| SSUBM407_0640 | Putative exported protein | 0.4292 | 3 |
| SSUBM407_0662 | Putative N-acetylmuramoyl-L-alanine amidase | 0.8672 | 6 |
| SSUBM407_1229 | Putative exported protein | 0.5271 | 1 |
| SSUBM407_1493 | Putative exported protein | 0.6369 | 6 |
| SSUBM407_1726 | LytR family regulatory protein | 0.5444 | 3 |
| SSUBM407_1898 | UTP--glucose-1-phosphate uridylyltransferase | 0.4444 | 6 |
| SSUBM407_2032 | Serine protease | 0.5736 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Torre, E.P.; Rodríguez-Franco, A.; Rodríguez-Ortega, M.J. Comparative Exoproteome Analysis of Streptococcus suis Human Isolates. Microorganisms 2021, 9, 1287. https://doi.org/10.3390/microorganisms9061287
de la Torre EP, Rodríguez-Franco A, Rodríguez-Ortega MJ. Comparative Exoproteome Analysis of Streptococcus suis Human Isolates. Microorganisms. 2021; 9(6):1287. https://doi.org/10.3390/microorganisms9061287
Chicago/Turabian Stylede la Torre, Esther Prados, Antonio Rodríguez-Franco, and Manuel J. Rodríguez-Ortega. 2021. "Comparative Exoproteome Analysis of Streptococcus suis Human Isolates" Microorganisms 9, no. 6: 1287. https://doi.org/10.3390/microorganisms9061287