
Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Virus Source, Mechanical Inoculation, and Symptom Evaluation
2.3. ToLCNDV Detection by Tissue Printing
2.4. ToLCNDV Detection by PCR and qPCR
2.5. Validation of Response to the Viral Infection and Generation of F1, F2, and BC Populations
2.6. Genotyping of Segregating Populations
3. Results
3.1. Response of the Spanish Landraces of C. sativus to the Mechanical Transmission of ToLCNDV
3.2. Response of the C. sativus Accessions from Different Origins to the Mechanical Transmission of ToLCNDV
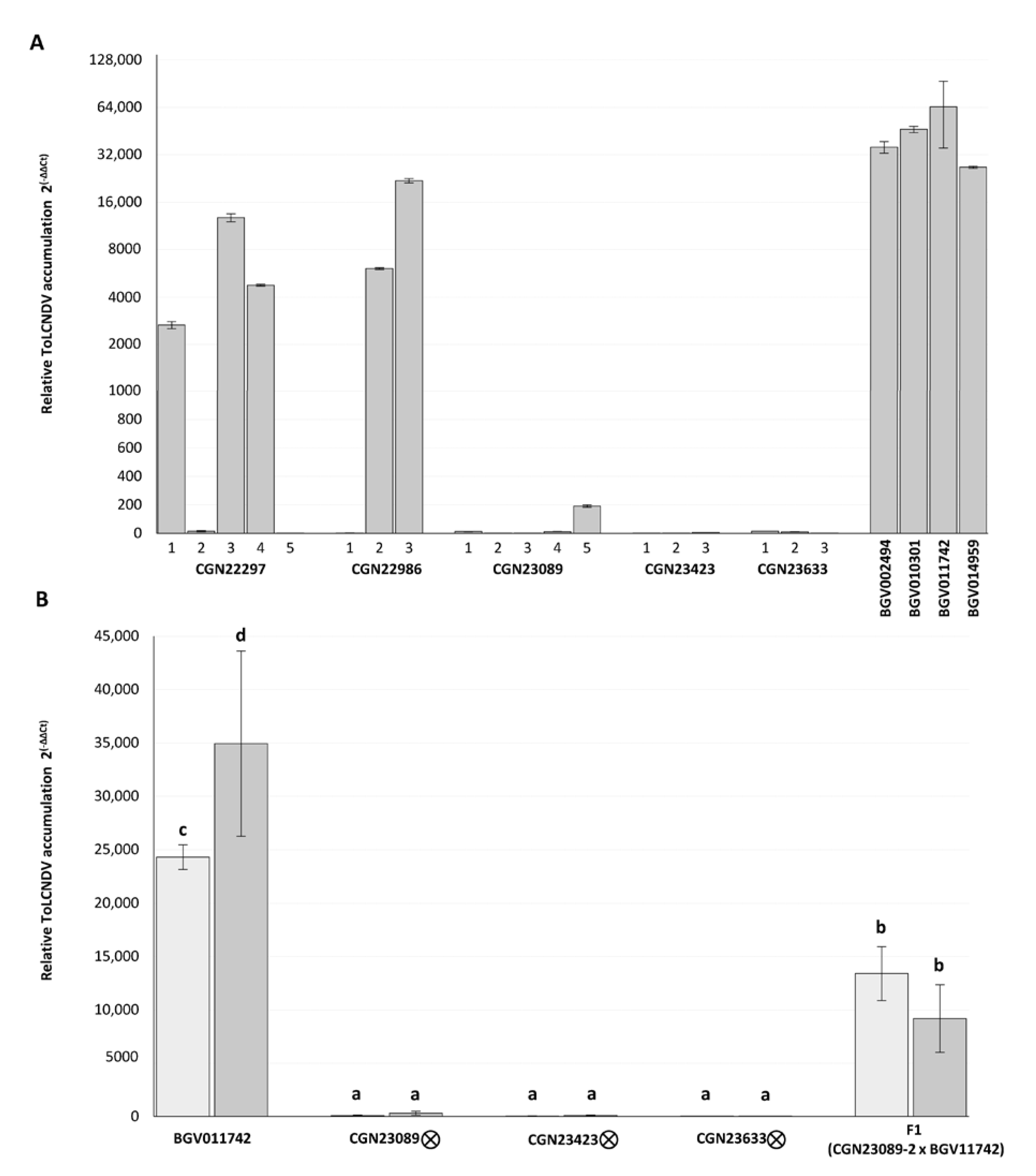
3.3. ToLCNDV Quantification in Resistant Genotypes
3.4. Response of Self-Pollinated and F1 Progenies to the Mechanical Transmission of ToLCNDV
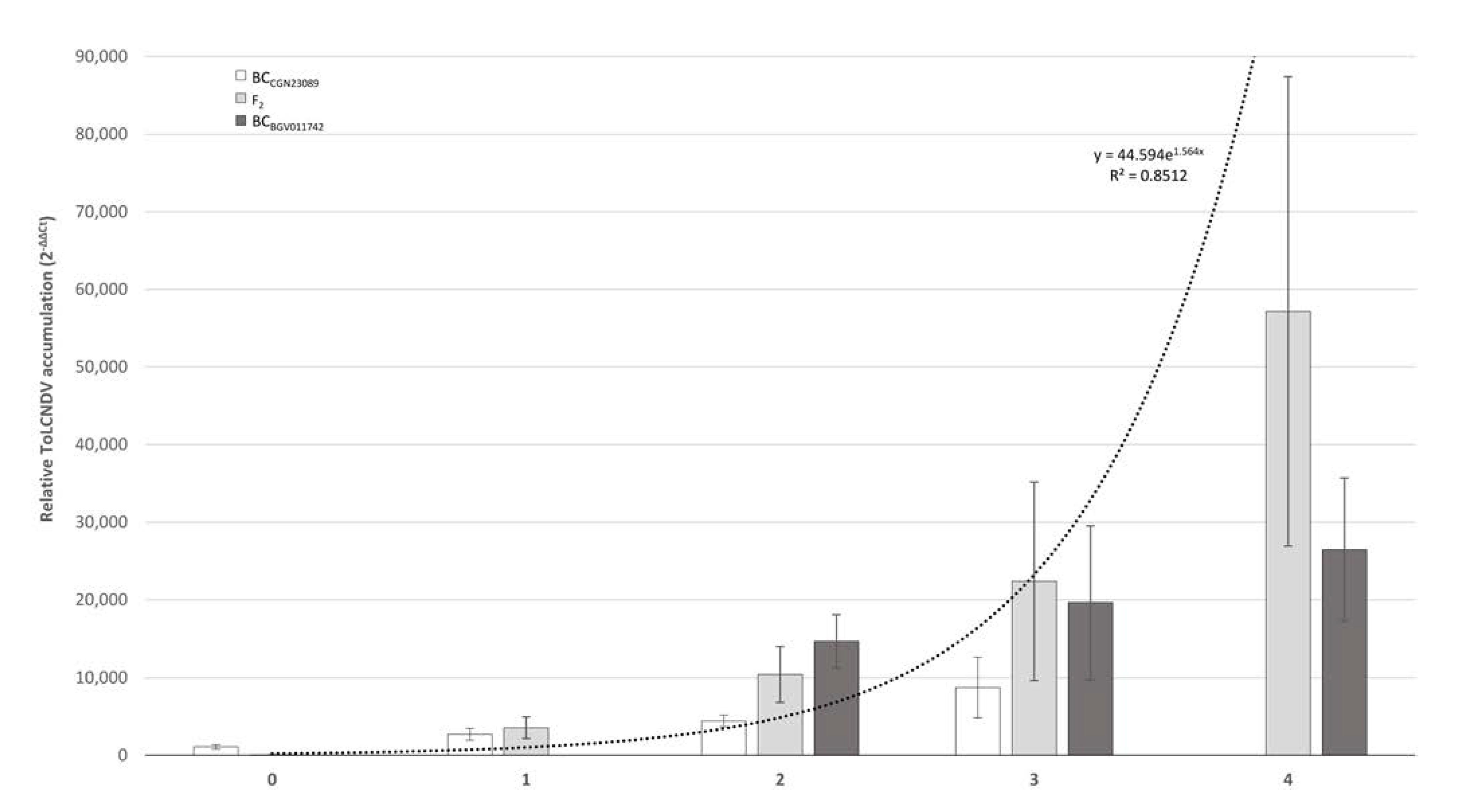
3.5. Response of Segregating Populations to the Mechanical Transmission of ToLCNDV
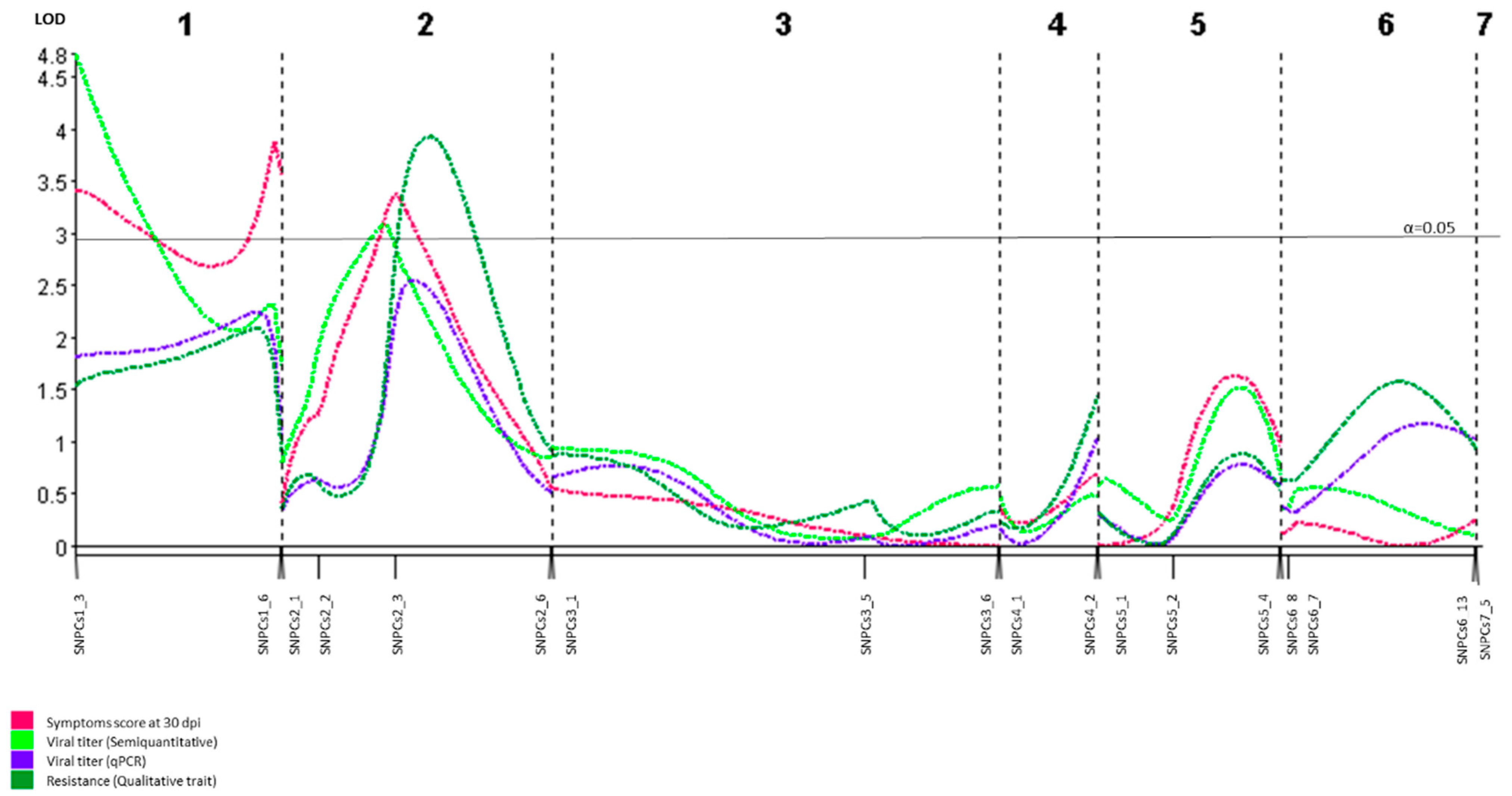
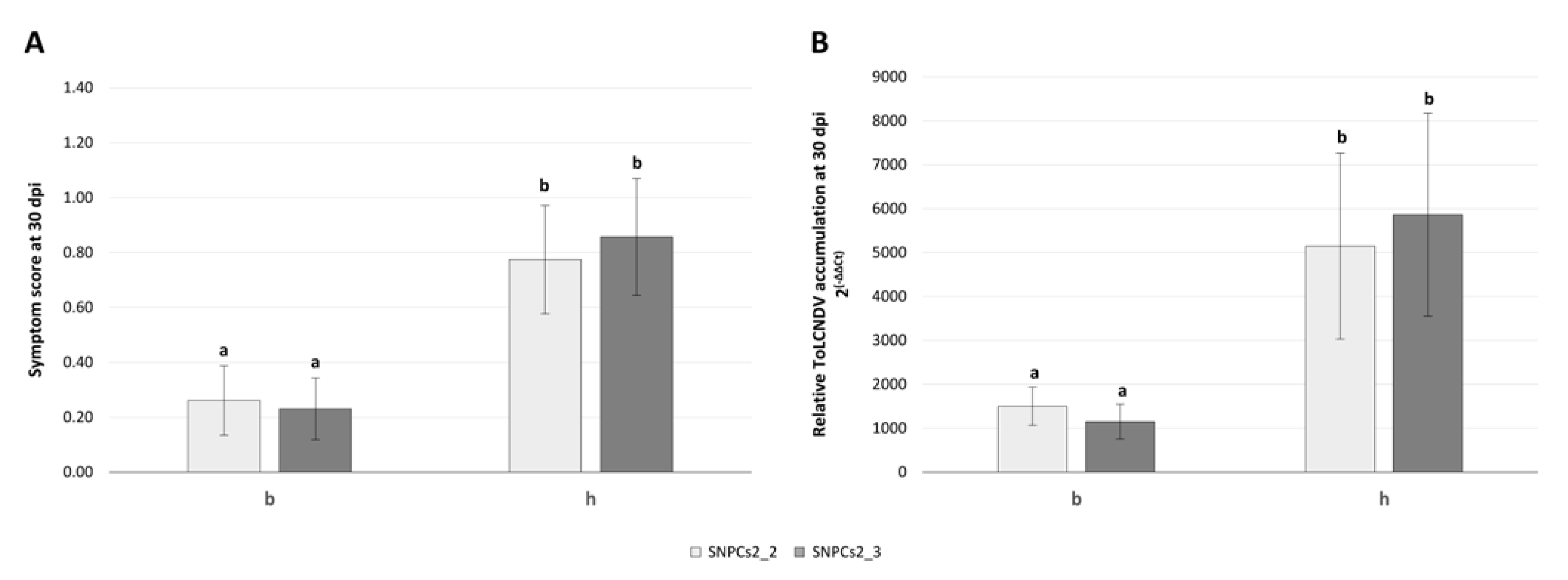
3.6. Genotyping and Linkage Analysis in Segregating Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations. Food and Agriculture Data. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 5 March 2021).
- Lecoq, H.; Katis, N. Control of cucurbit viruses. Adv.Virus Res. 2014, 90, 255–296. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, A.M.; Picó, B. Natural resistances to viruses in cucurbits. Agronomy 2021, 11, 23. [Google Scholar] [CrossRef]
- Srivastava, K.M.; Hallan, V.; Raizada, R.K.; Chandra, G.; Singh, B.P.; Sane, P.V. Molecular cloning of Indian tomato leaf curl virus genome following a simple method of concentrating the supercoiled replicative form of viral DNA. J. Virol. Methods 1995, 51, 297–304. [Google Scholar] [CrossRef]
- Moriones, E.; Praveen, S.; Chakraborty, S. Tomato leaf curl New Delhi virus: An emerging virus complex threatening vegetable and fiber crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato leaf curl New Delhi virus: A widespread bipartite begomovirus in the territory of monopartite begomoviruses. Mol. Plant Pathol. 2017, 18, 901–911. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of tomato leaf curl New Delhi virus infecting zucchini in Spain. Plant Dis. 2014, 98, 857–858. [Google Scholar] [CrossRef]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.S.; Bellon Doña, D.; ben Nahia, E.; Hajlaoui, M.R. First report of tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef] [Green Version]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33, 6. [Google Scholar] [CrossRef] [Green Version]
- Sifres, A.; Sáez, C.; Ferriol, M.; Selmani, E.A.; Riado, J.; Picó, B.; López, C. First report of tomato leaf curl New Delhi virus infecting zucchini in Morocco. Plant Dis. 2018, 102, 1045. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Malandraki, I.; Beris, D.; Keksidou, O.; Vassilakos, N.; Varveri, C.; Katis, N.; Maliogka, N.I. First report of tomato leaf curl New Delhi virus in zucchini crops in Greece. J. Plant Pathol. 2019, 101, 799. [Google Scholar] [CrossRef] [Green Version]
- Kheireddine, A.; Sifres, A.; Sáez, C.; Picó, B.; López, C. First report of tomato leaf curl New Delhi virus infecting cucurbit plants in Algeria. Plant Dis. 2019, 103, 3291. [Google Scholar] [CrossRef]
- EPPO. European and Mediterranean Plant Protection Organization. Available online: //gd.eppo.int/taxon/TOLCND/distribution (accessed on 20 November 2019).
- Luigi, M.; Bertin, S.; Manglli, A.; Troiano, E.; Davino, S.; Tomassoli, L.; Parrella, G. First report of tomato leaf curl New Delhi virus causing yellow leaf curl of pepper in Europe. Plant Dis. 2019, 103, 2970. [Google Scholar] [CrossRef]
- Jyothsna, P.; Haq, Q.M.I.; Singh, P.; Sumiya, K.V.; Praveen, S.; Rawat, R.; Briddon, R.W.; Malathi, V.G. Infection of tomato leaf curl New Delhi virus (ToLCNDV), a bipartite begomovirus with betasatellites, results in enhanced level of helper virus components and antagonistic interaction between DNA B and betasatellites. Appl. Microbiol. Biotechnol. 2013, 97, 5457–5471. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Ku, H.M.; Tsai, W.S.; Chien, R.C.; Jan, F.J. Identification and characterization of a mechanical transmissible begomovirus causing leaf curl on oriental melon. Eur. J. Plant Pathol. 2010, 127, 219–228. [Google Scholar] [CrossRef]
- Islam, S.; Munshi, A.D.; Verma, M.; Arya, L.; Mandal, B.; Behera, T.K.; Kumar, R.; Lal, S.K. Genetics of resistance in Luffa cylindrica Roem. against tomato leaf curl New Delhi virus. Euphytica 2010, 174, 83–89. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Karim, S.; Varma, A.; Abuzenadah, A.M.; Chaudhary, A.G.; Damanhouri, G.A.; Mandal, B. Characterization of tomato leaf curl New Delhi virus infecting cucurbits: Evidence for sap transmission in a host specific manner. Afr. J. Biotechnol. 2013, 12, 5000–5009. [Google Scholar] [CrossRef] [Green Version]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical transmission of tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 279–691. [Google Scholar] [CrossRef]
- Sangeetha, B.; Malathi, V.G.; Alice, D.; Suganthy, M.; Renukadevi, P. A distinct seed-transmissible strain of tomato leaf curl New Delhi virus infecting chayote in India. Virus Res. 2018, 251, 81–91. [Google Scholar] [CrossRef]
- Kil, E.J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S.K. Seed transmission of tomato leaf curl New Delhi virus from zucchini squash in Italy. Plants 2020, 9, 563. [Google Scholar] [CrossRef]
- EPPO. European and Mediterranean Plant Protection Organization. Available online: https://www.eppo.int/QUARANTINE/Alert_List/viruses/ToLCNDV.htm (accessed on 28 February 2017).
- Islam, S.; Munshi, A.D.; Verma, M.; Arya, L.; Mandal, B.; Behera, T.K.; Kumar, R.; Lal, S.K. Screening of Luffa cylindrica Roem. for resistance against tomato leaf curl New Delhi virus, inheritance of resistance, and identification of SRAP markers linked to the single dominant resistance gene. J. Hortic. Sci. Biotechnol. 2011, 86, 661–667. [Google Scholar] [CrossRef]
- Sáez, C.; Esteras, C.; Martínez, C.; Ferriol, M.; Narinder, P.S.D.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in melon is controlled by a major QTL located in chromosome 11. Plant Cell Rep. 2017, 36, 1571–1584. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Montero-Pau, J.; Esteras, C.; Blanca, J.; Sifres, A.; Ferriol, M.; López, C.; Picó, B. A major QTL located in chromosome 8 of Cucurbita moschata is responsible for resistance to tomato leaf curl New Delhi virus (ToLCNDV). Front. Plant Sci. 2020, 11, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcárcel, J.V.; Peiró, R.M.; Pérez-de-Castro, A.; Díez, M.J. Morphological characterization of the cucumber (Cucumis sativus L.) collection of the COMAV’s Genebank. Genet. Resour.Crop Evol. 2018, 65, 1293–1306. [Google Scholar] [CrossRef]
- Valcárcel, J.V.; Pérez-de-Castro, A.; Díez, M.J.; Peiró, R.M. Molecular characterization of the cucumber (Cucumis sativus L.) accessions of the COMAV’s genebank. Span. J. Agric. Res. 2018, 16, e0701. [Google Scholar] [CrossRef]
- Aparicio, F.; Soler, S.; Aramburu, J.; Galipienso, L.; Nuez, F.; Pallás, V.; López, C. Simultaneous detection of six RNA plant viruses affecting tomato crops using a single digoxigenin-labelled polyprobe. Eur.J. Plant Pathol. 2009, 123, 117–123. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, L.; Cheng, F.; Huang, Y.; Bie, Z. Evaluation of appropriate reference genes for gene expression normalization during watermelon fruit development. PLoS ONE 2015, 10, e0130865. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta Ct) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bao, K.; Reddy, U.K.; Bai, Y.; Hammar, S.A.; Jiao, C.; Wehner, T.C.; Ramírez-Madera, A.O.; Weng, Y.; Grumet, R.; et al. The USDA cucumber (Cucumis sativus L.) collection: Genetic diversity, population structure, genome-wide association studies, and core collection development. Hortic. Res. 2018, 5, 64. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. [Google Scholar]
- Lincoln, S.; Daly, M.; Lander, E. Constructing Genetic Maps with MAPMAKER/EXP 3.0. Whitehead Institute Technical Report, 3rd ed.; Whitehead Institute: Cambridge, MA, USA, 1992. [Google Scholar]
- Joehanes, R.; Nelson, J.C. QGene 4.0, an extensible Java QTL-analysis platform. Bioinformatics 2008, 24, 2788–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ooijen, J.W. MapQTL® 6 Software for the Mapping of Quantitative Trait Loci in Experimental Population of Diploid Species; Kayzma BV: Wageningen, The Netherlands, 2009. [Google Scholar]
- RAIF, Red de Alerta e Información Fitosanitaria de Andalucía. Situación Actual del Virus del Rizado de Nueva Delhi (ToLCNDV) Bajo Invernadero en Cultivo de PEPINO en la Provincia de Almería. 2015. Available online: //www.juntadeandalucia.es/agriculturapescaydesarrollorural/raif/ca/novedades/-/asset_publisher/4rpcT3lrh8uL/content/situacion-actual-del-virus-del-rizado-de-nueva-delhi-tolcndv-bajo-invernadero-en-cultivo-de-pepino-en-la-provincia-de-almeri-1?inheritRedirect=false (accessed on 25 March 2021).
- Weng, Y.; Wehner, T.C. Cucumber gene catalog 2017. Rep. Cucurbit Genet. Coop. 2017, 39–40, 17–54. [Google Scholar]
- Tian, G.L.; Yang, Y.H.; Zhang, S.P.; Miao, H.; Lu, H.W.; Wang, Y.; Xie, B.Y.; Gu, X.F. Genetic analysis and gene mapping of papaya ring spot virus resistance in cucumber. Mol. Breed. 2015, 35, 110. [Google Scholar] [CrossRef]
- Tian, G.L.; Miao, H.; Yang, Y.H.; Zhou, J.; Lu, H.W.; Wang, Y.; Xie, B.Y.; Zhang, S.P.; Gu, X.F. Genetic analysis and fine mapping of watermelon mosaic virus resistance gene in cucumber. Mol. Breed. 2016, 36, 131. [Google Scholar] [CrossRef]
- Shi, L.; Yang, Y.; Xie, Q.; Miao, H.; Bo, K.; Song, Z.; Wang, Y.; Xie, B.; Zhang, S.; Gu, X. Inheritance and QTL mapping of cucumber mosaic virus resistance in cucumber (Cucumis sativus L.). PLoS ONE 2018, 13, e0200571. [Google Scholar] [CrossRef]
- Munshi, A.D.; Panda, B.; Mandal, B.; Bisht, I.S.; Rao, E.S.; Kumar, R. Genetics of resistance to cucumber mosaic virus in Cucumis sativus var. hardwickii R. Alef. Euphytica 2008, 164, 501–507. [Google Scholar] [CrossRef]
- Picó, B.; Villar, C.; Nuez, F.; Weber, W.E. Screening Cucumis sativus landraces for resistance to cucumber vein yellowing virus. Plant Breed. 2003, 122, 426–430. [Google Scholar] [CrossRef]
- Aguilar, J.M.; Abad, J.; Aranda, M.A. Resistance to cucurbit yellow stunting disorder virus in cucumber. Plant Dis. 2006, 90, 583–586. [Google Scholar] [CrossRef]
- Crespo, O.; Janssen, D.; Robles, C.; Ruiz, L. Resistance to cucumber green mottle mosaic virus in Cucumis sativus. Euphytica 2018, 214, 201. [Google Scholar] [CrossRef]
- Romero-Masegosa, J.; Martínez, C.; Aguado, E.; García, A.; Cebrián, G.; Iglesias-Moya, J.; Paris, H.S.; Jamilena, M. Response of Cucurbita spp. to tomato leaf curl New Delhi virus inoculation and identification of a dominant source of resistance in Cucurbita moschata. Plant Pathol. 2021, 70, 206–218. [Google Scholar] [CrossRef]
- Dhillon, N.P.S.; Monforte, A.J.; Pitrat, M.; Pandey, S.; Singh, P.K.; Reitsma, K.R.; García-Mas, J.; Sharma, A.; McCreight, J.D. Melon landraces of India: Contributions and importance. Plant Breed. Rev. 2012, 35, 85–150. [Google Scholar] [CrossRef]
- Díaz-Pendón, J.A.; Truniger, V.; Nieto, C.; García-Mas, J.; Bendahmane, A.; Aranda, M.A. Advances in understanding recessive resistance to plant viruses. Mol. Plant Pathol. 2004, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Wege, C. Movement and localization of Tomato yellow leaf curl viruses in the infected plant; In Tomato Yellow Leaf Curl Virus Disease; Springer: Dordrecht, The Netherlands, 2007; pp. 185–206. [Google Scholar] [CrossRef]
- Marco, C.F.; Aguilar, J.M.; Abad, J.; Gómez-Guillamón, M.L.; Aranda, M.A. Melon resistance to cucurbit yellow stunting disorder virus in characterized by reduced virus accumulation. Phytopathology 2003, 93, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Pendón, J.A.; Fernández-Muñoz, R.; Gómez-Guillamón, M.L.; Moriones, E. Inheritance of resistance to watermelon mosaic virus in Cucumis melo that impairs virus accumulation, symptom expression, and aphid transmission. Phytopathology 2005, 95, 840–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, D.A.; Rezende, J.A.M.; Piedade, S.M.dS. Biomass, virus concentration, and symptomatology of cucurbits infected by mild and severe strains of papaya ringspot virus. Sci. Agric. 2003, 60, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Romay, G.; Pitrat, M.; Lecoq, H.; Wipf-Scheibel, C.; Millot, P.; Girardot, G.; Desbiez, C. Resistance against melon chlorotic mosaic virus and tomato leaf curl New Delhi virus in melon. Plant Dis. 2019, 103, 2913–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, A.; Pal, A. Identification and characterization of elite inbred lines with MYMIV-resistance in Vigna mungo. Field Crops Res. 2012, 135, 116–125. [Google Scholar] [CrossRef]
- Kushwaha, N.; Singh, A.K.; Basu, S.; Chakraborty, S. Differential response of diverse solanaceous hosts to tomato leaf curl New Delhi virus infection indicates coordinated action of NBS-LRR and RNAi-mediated host defense. Arch. Virol. 2015, 160, 1499–1509. [Google Scholar] [CrossRef]
- Jeevalatha, A.; Siddappa, S.; Kumar, A.; Kaundal, P.; Guleria, A.; Sharma, S.; Nagesh, M.; Singh, B.P. An insight into differentially regulated genes in resistant and susceptible genotypes of potato in response to tomato leaf curl New Delhi virus-[potato] infection. Virus Res. 2017, 232, 22–33. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Ohnishi, J.; Saito, A.; Ohyama, A.; Nunome, T.; Miyatake, K.; Fukuoka, H. An NB-LRR gene, TYNBS1, is responsible for resistance mediated by the Ty-2 Begomovirus resistance locus of tomato. Theor. Appl. Genet. 2018, 131, 1345–1362. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Das, R.; Mandal, A.; Kundu, P. Functional characterization of tomato membrane-bound NAC transcription factors. Plant Mol. Biol. 2017, 93, 511–532. [Google Scholar] [CrossRef]
- Soler-Garzón, A.; Oladzad, A.; Beaver, J.; Beebe, S.; Lee, R.; Lobaton, J.D.; Macea, E.; McClean, P.; Raatz, B.; Rosas, J.C.; et al. NAC Candidate gene marker for bgm-1 and interaction with QTL for resistance to bean golden yellow mosaic virus in common bean. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, R.M.; Picard, C.L. RNA-Directed DNA Methylation. PLoS Genet. 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.P.; Sharma, N.; Puranik, S.; Chakraborty, S.; Prasad, M. Tomato 26S proteasome subunit RPT4a regulates ToLCNDV transcription and activates hypersensitive response in tomato. Sci. Rep. 2016, 6, 27078. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symptoms at 15 Dpi | Symptoms at 30 Dpi | Viral Load | |||||
|---|---|---|---|---|---|---|---|
| Type | Genebank Code | Spanish Province | Mean | Range | Mean | Range | Tissue Printing |
| Short | BGV000047 | Zaragoza | 1.2 | (1–2) | 1.8 | (1–2) | +++ |
| BGV000408 | Cádiz | 1.6 | (1–2) | 1.6 | (1–3) | +++ | |
| BGV000437 | Jaén | 1.2 | (0–3) | 1.8 | (1–2) | ++ | |
| BGV000467 | Jaén | 1.0 | (0–2) | 1.4 | (0–3) | +++ | |
| BGV000479 | Córdoba | 1.0 | (1) | 1.0 | (0–2) | +++ | |
| BGV000512 | Huelva | 1.0 | (0–2) | 1.4 | (1–2) | +++ | |
| BGV002495 | Tenerife | n/a | 0.6 | (0–2) | ++ | ||
| BGV003714 | Cuenca | 2.0 | (1–3) | 1.7 | (1–2) | +++ | |
| BGV004026 | Cáceres | 1.4 | (1–2) | 2.5 | (1–3) | +++ | |
| BGV004304 | Murcia | n/a | 2.0 | (1–3) | +++ | ||
| BGV008299 | Valencia | 2.0 | (1–3) | 2.4 | (2–3) | ++ | |
| BGV010301 | Guadalajara | 1.4 | (0–3) | 1.6 | (0–3) | +++ | |
| BGV010314 | Guadalajara | 1.7 | (1–2) | 1.2 | (0–2) | + | |
| BGV010636 | Soria | 2.4 | (1–4) | 3.4 | (2–4) | ++ | |
| BGV011582 | Teruel | 0.8 | (0–2) | 1.8 | (1–2) | ++ | |
| BGV011734 | Valladolid | 2.0 | (0–4) | 3.2 | (3–4) | ++ | |
| BGV011736 | Ávila | 2.6 | (0–4) | 2.8 | (2–4) | ++ | |
| BGV011742 | Albacete | 3.4 | (3–4) | 3.8 | (3–4) | ++ | |
| BGV014959 | Huesca | 3.8 | (3–4) | 3.6 | (3–4) | ++ | |
| BGV015469 | Cáceres | 1.6 | (0–2) | 2.8 | (2–4) | + | |
| Long | BGV000372 | Granada | 0.6 | (0–1) | 1.2 | (0–2) | ++ |
| BGV000381 | Málaga | 0.6 | (0–1) | 2.4 | (2–3) | +++ | |
| BGV000416 | Cádiz | 1.4 | (0–2) | 2.0 | (1–3) | ++ | |
| BGV001310 | Asturias | 1.0 | (0–2) | 1.6 | (1–2) | +++ | |
| BGV002494 | Tenerife | 2.0 | (2) | 1.0 | (0–2) | +++ | |
| BGV004305 | Murcia | 1.6 | (1–2) | 1.2 | (1–2) | +++ | |
| BGV004309 | Murcia | 1.4 | (1–2) | 3.0 | (3) | +++ | |
| BGV004851 | Castellón | 0.0 | (0) | 1.0 | (0–2) | ++ | |
| BGV004926 | Valencia | 2.0 | (2) | 2.0 | (2) | +++ | |
| BGV004936 | Valencia | 1.4 | (1–2) | 1.3 | (1–2) | ++ | |
| BGV011586 | Orense | 0.6 | (0–3) | 0.6 | (0–3) | + | |
| BGV011724 | Teruel | 1.8 | (0–4) | 1.8 | (0–4) | + | |
| BGV014967 | Guadalajara | 2.2 | (0–4) | 2.4 | (1–4) | ++ | |
| BGV015229 | Vizcaya | 2.5 | (1–3) | 2.5 | (2–3) | ++ | |
| BGV015696 | Alicante | 2.4 | (0–4) | 2.6 | (0–4) | ++ | |
| BGV015700 | Girona | 2.8 | (1–4) | 3.4 | (2–4) | ++ | |
| French | BGV010290 | Granada | 2.8 | (0–4) | 3.8 | (3–4) | +++ |
| BGV011735 | Zaragoza | 2.3 | (0–3) | 2.3 | (0–4) | ++ | |
| BGV014961 | Castellón | 3.0 | (0–4) | 2.6 | (0–4) | +++ | |
| BGV014969 | Cantabria | 1.6 | (0–3) | 2.4 | (0–4) | +++ | |
| Symptoms at 15 Dpi | Symptoms at 30 Dpi | Viral Load | ||||||
|---|---|---|---|---|---|---|---|---|
| Type | Genebank Code | Country | Local Name | Mean | Range | Mean | Range | Tissue Printing |
| Short | CGN19748 | India | Khira | 3.5 | (3–4) | 3.0 | (3) | ++ |
| CGN19817 | India | Cucumber Medium | 1.4 | (0–3) | 3.0 | (2–4) | +++ | |
| CGN20512 | Netherlands | 752 | 2.5 | (2–3) | 2.8 | (2–3) | ++ | |
| CGN20517 | Sri Lanka | Yellow 1 | 1.0 | (0–2) | 1.3 | (0–2) | + | |
| CGN21585 | India | Saharanpur | 0.3 | (0–1) | 2.3 | (1–4) | n/a | |
| CGN21691 | D.R. Congo | N2/81 | 2.8 | (0–4) | 3.2 | (1–4) | +++ | |
| CGN22280 | India | Shuei Huang Kua | 1.0 | (0–3) | 1.0 | (1) | +++ | |
| CGN22986 | India | Smallgreen | 0.4 | (0–1) | 0.4 | (0–1) | ++ | |
| CGN23089 | India | Anthracnose 197087 | 0.2 | (0–1) | 0.0 | (0) | - | |
| CGN23411 | India | Khira Cheshuicchatyi | 0.6 | (0–2) | 1.0 | (0–3) | ++ | |
| CGN23423 | India | JL-2 Dhillon | 0.0 | (0) | 0.3 | (0–1) | - | |
| CGN23633 | India | Jaipur Balam | 0.0 | (0) | 0.7 | (0-1) | - | |
| Medium | CGN19819 | India | Puneri Klura | 0.8 | (0–1) | 1.3 | (1–2) | +++ |
| CGN20853 | Japan | Sagami Hanpaku Fushinari Kyuri | 1.5 | (0–2) | 1.5 | (1–2) | +++ | |
| CGN21616 | Iran | Rasht | 3.3 | (2–4) | 3.7 | (3–4) | +++ | |
| CGN22281 | India | Long Green | 0.8 | (0–2) | 1.5 | (0–2) | ++ | |
| CGN22297 | India | K-75 | 0.4 | (0–1) | 0.8 | (0–2) | + | |
| Long | BGV015107 | China | Hei Wu She | 1.2 | (0–2) | 1.6 | (1–3) | ++ |
| BGV015113 | China | Shou Guang Qiu Gua | 2.0 | (1–3) | 1.6 | (1–2) | ++ | |
| BGV015115 | China | Long Quan Qing Huang Gua | 1.6 | (0–4) | 1.6 | (1–3) | ++ | |
| BGV015116 | China | De Hui Huang Gua | 2.0 | (0–3) | 2.6 | (1–4) | +++ | |
| BGV015118 | China | San Ye Zao | 2.5 | (0–4) | 3.3 | (1–4) | +++ | |
| - | CGN19655 | U.S.A. | SC 53-B (6) | 1.4 | (0–4) | 2.4 | (0–4) | +++ |
| Populations | Resistant | Susceptible | Expected Frequencies | X2 |
|---|---|---|---|---|
| F2 | 31 | 65 | 1:3 | 2.722 (p = 0.0990) |
| BCCGN23089 | 21 | 33 | 1:1 | 2.667 (p = 0.1025) |
| BCBGV011742 | 0 | 11 | - | - |
| Trait | Chr a | Nearest Marker b | CIM | Kruskal–Wallis | |||||
|---|---|---|---|---|---|---|---|---|---|
| Interval c (cM) | Add Effect d | Dom Effect e | LOD f | R2g | K*h | Significance i | |||
| Symptoms 30 dpi | 2 | SNPCs2_3 | 28–40 | 0.46 | 0.73 | 3.38 | 0.15 | 16.94 | ****** |
| Viral load (Semiquantitative) | 2 | SNPCs2_3 | 24–32 | 0.18 | 0.79 | 3.07 | 0.14 | 13.01 | **** |
| Viral load (Quantitative, ΔΔCt) | 2 | SNPCs2_3 | - | −1.45 | −4.61 | 2.54 | 0.12 | 14.05 | ***** |
| Resistance (Qualitative trait) | 2 | SNPCs2_3 | 34–54 | −0.23 | −0.57 | 3.93 | 0.17 | 13.02 | **** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáez, C.; Ambrosio, L.G.M.; Miguel, S.M.; Valcárcel, J.V.; Díez, M.J.; Picó, B.; López, C. Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber. Microorganisms 2021, 9, 913. https://doi.org/10.3390/microorganisms9050913
Sáez C, Ambrosio LGM, Miguel SM, Valcárcel JV, Díez MJ, Picó B, López C. Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber. Microorganisms. 2021; 9(5):913. https://doi.org/10.3390/microorganisms9050913
Chicago/Turabian StyleSáez, Cristina, Laura G. M. Ambrosio, Silvia M. Miguel, José Vicente Valcárcel, María José Díez, Belén Picó, and Carmelo López. 2021. "Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber" Microorganisms 9, no. 5: 913. https://doi.org/10.3390/microorganisms9050913