
US3 Serine/Threonine Protein Kinase from MDV-1, MDV-2, and HVT Differentially Regulate Viral Gene Expression and Replication

 , and
, and
Abstract
:1. Introduction
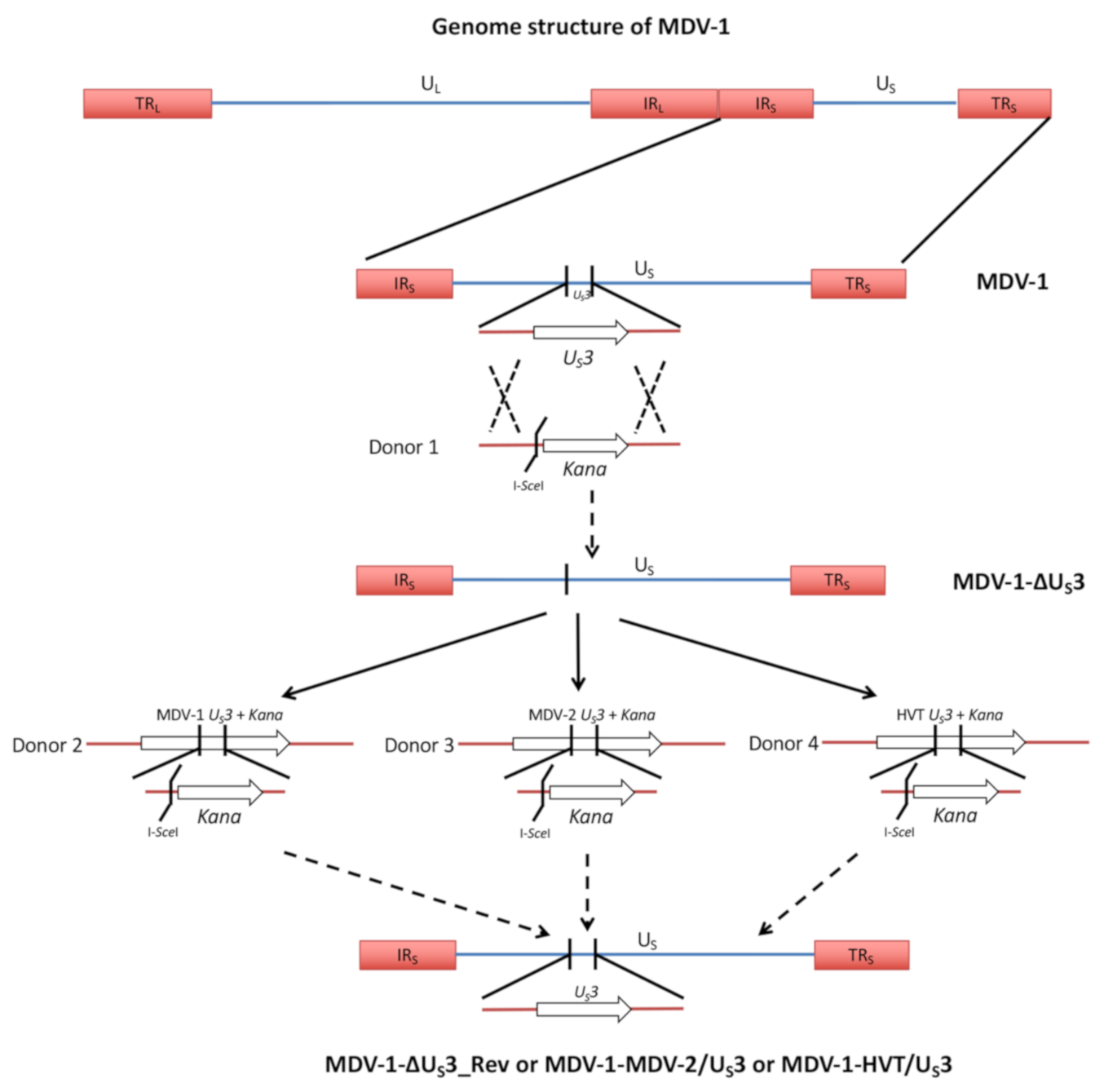
2. Materials and Methods
2.1. Viruses and Cell Culture
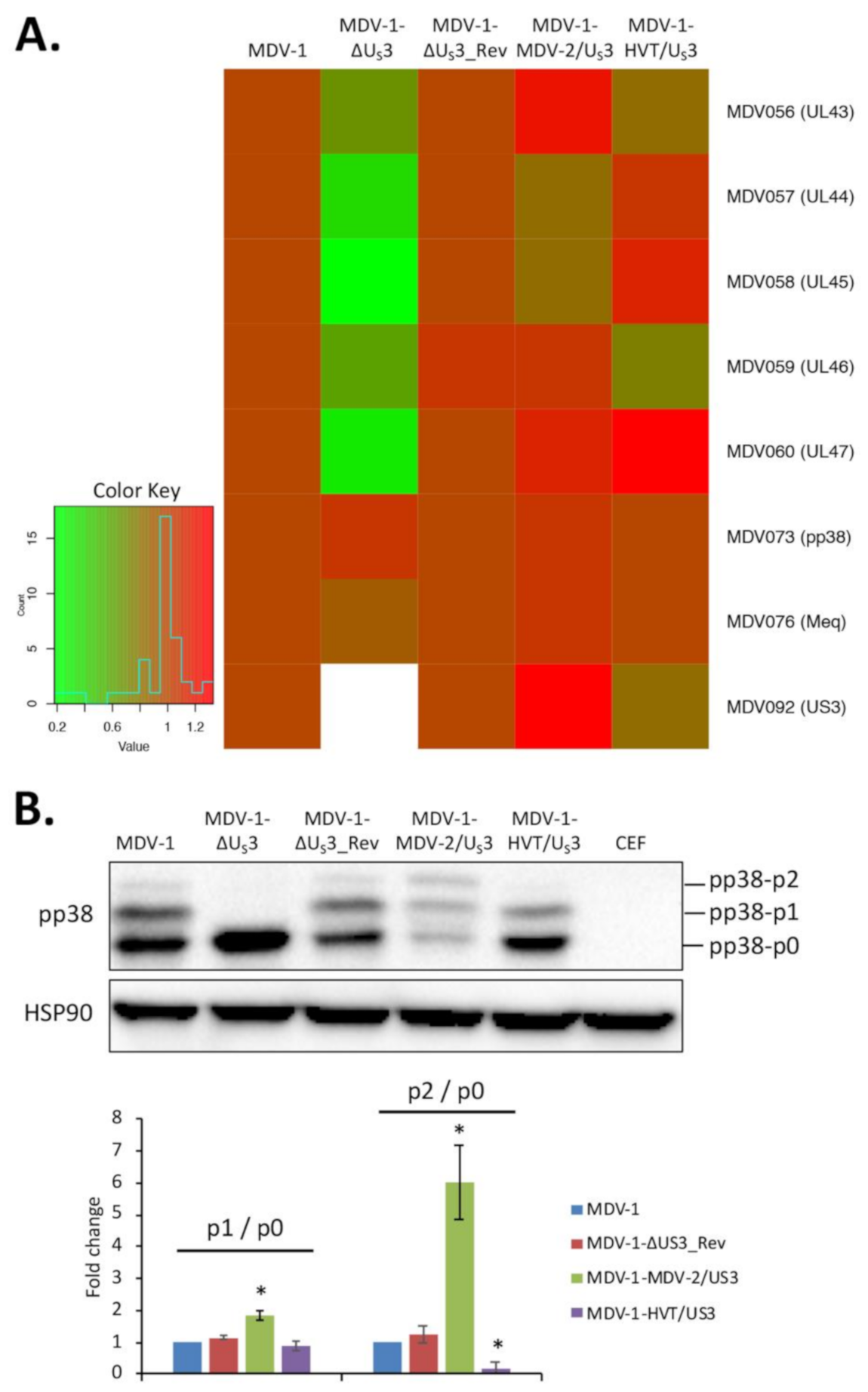
2.2. Quantitative Polymerase Chain Reaction (qPCR) and Western Blot
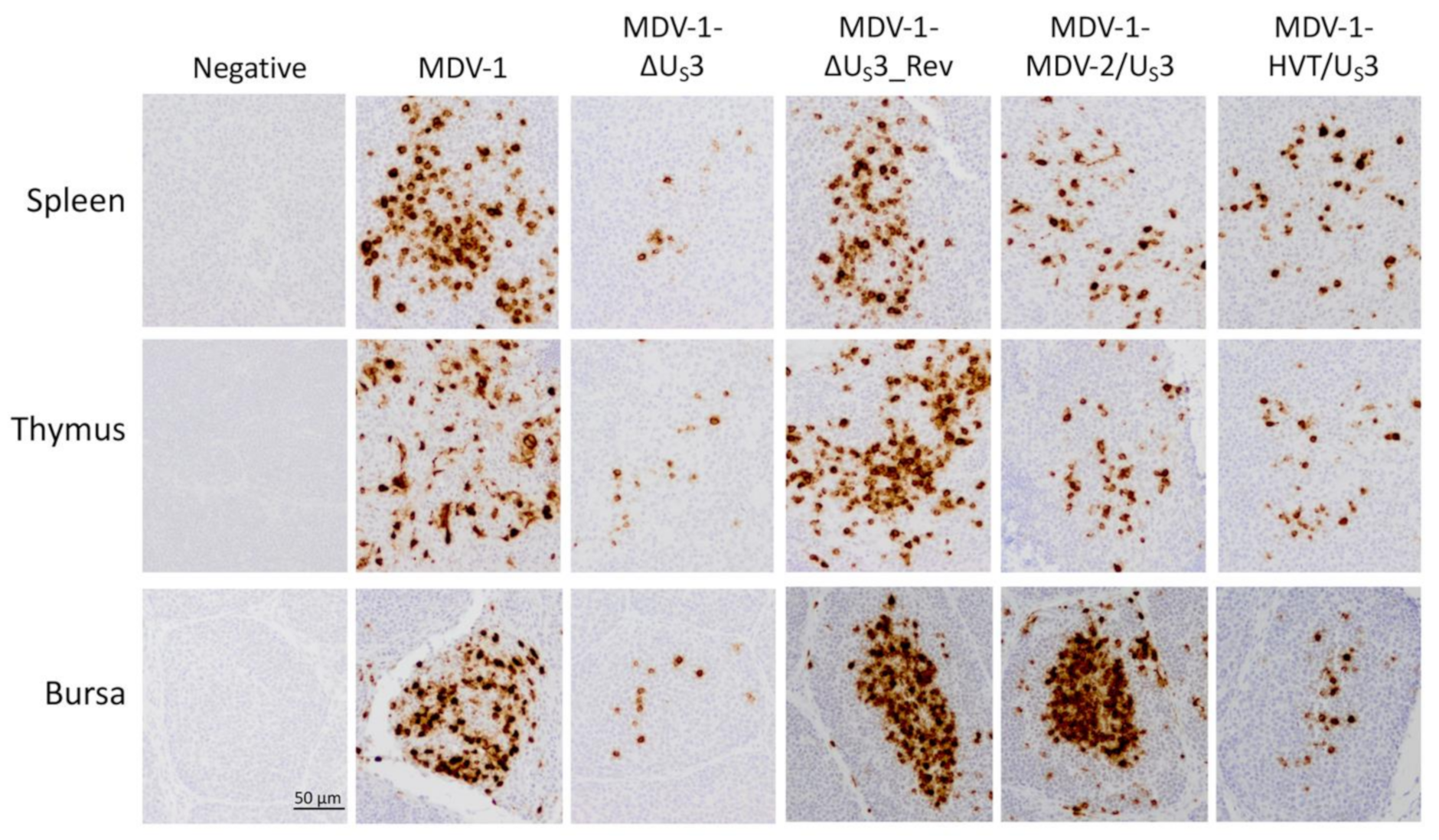
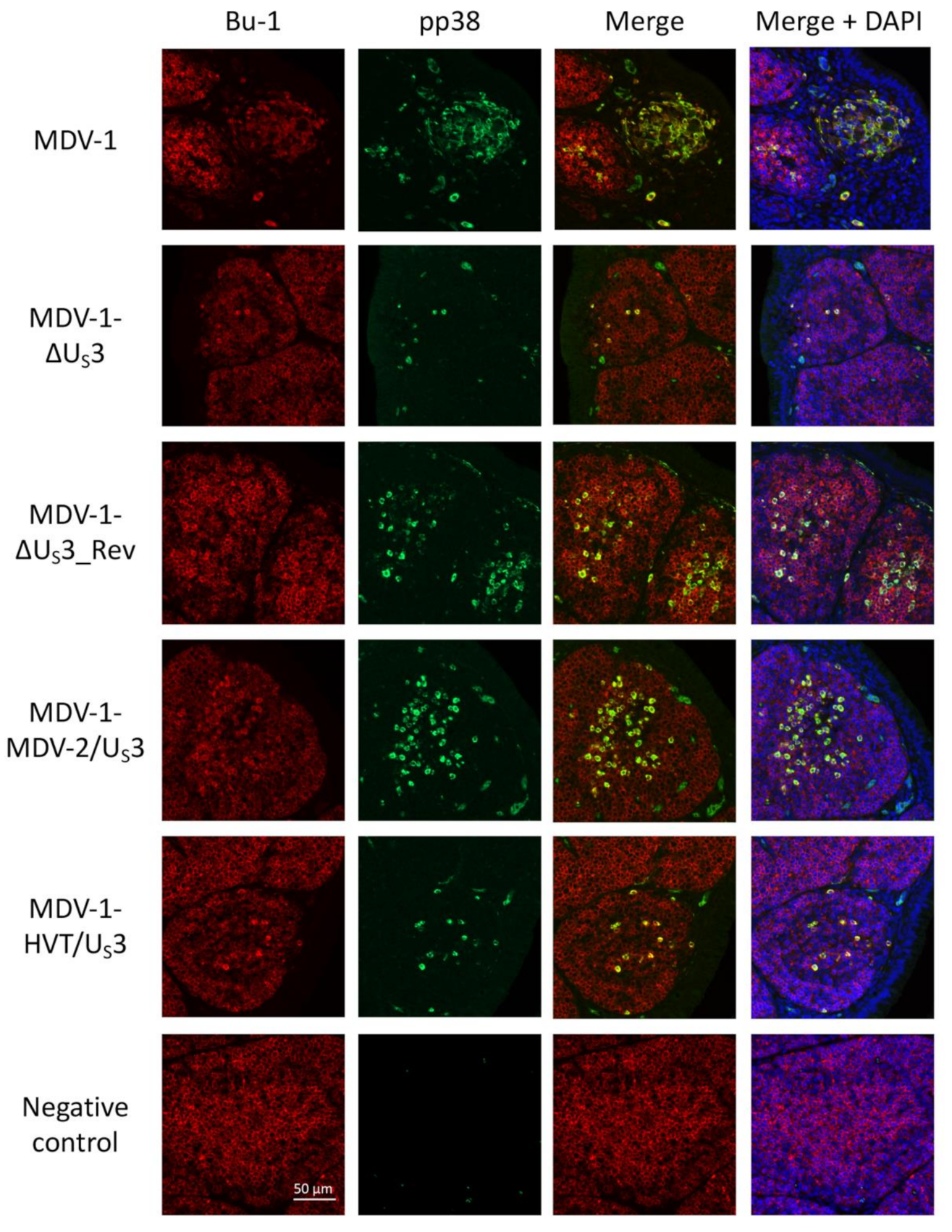
2.3. Immunohistochemistry (IHC) and Immunofluorescence Assay (IFA)
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s disease in chickens: A review with focus on immunology. Vet. Res. 2016, 47, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s disease virus: From miasma to model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Bertzbach, L.D.; Laparidou, M.; Hartle, S.; Etches, R.J.; Kaspers, B.; Schusser, B.; Kaufer, B.B. Unraveling the role of B cells in the pathogenesis of an oncogenic avian herpesvirus. Proc. Natl. Acad. Sci. USA 2018, 115, 11603–11607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deruelle, M.J.; Favoreel, H.W. Keep it in the subfamily: The conserved alphaherpesvirus US3 protein kinase. J. Gen. Virol. 2011, 92, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Arii, J.; Shiratori, I.; Akashi, H.; Arase, H.; Kawaguchi, Y. Herpes simplex virus 1 protein kinase Us3 phosphorylates viral envelope glycoprotein B and regulates its expression on the cell surface. J. Virol. 2009, 83, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, A.; Yamamoto, M.; Ohno, T.; Tanaka, M.; Sata, T.; Nishiyama, Y.; Kawaguchi, Y. Herpes simplex virus 1-encoded protein kinase UL13 phosphorylates viral Us3 protein kinase and regulates nuclear localization of viral envelopment factors UL34 and UL31. J. Virol. 2006, 80, 1476–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Patenode, C.; Roizman, B. US3 protein kinase of HSV-1 cycles between the cytoplasm and nucleus and interacts with programmed cell death protein 4 (PDCD4) to block apoptosis. Proc. Natl. Acad. Sci. USA 2011, 108, 14632–14636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, M.S.; Kinchington, P.R.; Banfield, B.W.; Silverstein, S. Hyperphosphorylation of histone deacetylase 2 by alphaherpesvirus US3 kinases. J. Virol. 2010, 84, 9666–9676. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ni, L.; Wang, S.; Zheng, C. Herpes simplex virus 1 protein kinase US3 hyperphosphorylates p65/RelA and dampens NF-kappaB activation. J. Virol. 2014, 88, 7941–7951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Li, T.; Zeng, M.; Peng, T. Herpes simplex virus type 1 infection activates the Epstein-Barr virus replicative cycle via a CREB-dependent mechanism. Cell Microbiol. 2012, 14, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Urakawa, T.; Hirayama, Y.; Miki, N.; Yamamoto, M.; Zhu, G.S.; Hirai, K. Marek’s disease virus protein kinase gene identified within the short unique region of the viral genome is not essential for viral replication in cell culture and vaccine-induced immunity in chickens. Virology 1993, 195, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, D.; Tischer, B.K.; Trapp, S.; Osterrieder, N. The protein encoded by the US3 orthologue of Marek’s disease virus is required for efficient de-envelopment of perinuclear virions and involved in actin stress fiber breakdown. J. Virol. 2005, 79, 3987–3997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, D.; McKinney, C.; Kaufer, B.B.; Osterrieder, N. Enzymatically inactive U(S)3 protein kinase of Marek’s disease virus (MDV) is capable of depolymerizing F-actin but results in accumulation of virions in perinuclear invaginations and reduced virus growth. Virology 2018, 375, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.M.; Lupiani, B.; Gimeno, I.M.; Silva, R.F.; Lee, L.F.; Witter, R.L. Rescue of a pathogenic Marek’s disease virus with overlapping cosmid DNAs: Use of a pp38 mutant to validate the technology for the study of gene function. Proc. Natl. Acad. Sci. USA 2002, 99, 7054–7059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimeno, I.M.; Witter, R.L.; Hunt, H.D.; Reddy, S.M.; Lee, L.F.; Silva, R.F. The pp38 gene of Marek’s disease virus (MDV) is necessary for cytolytic infection of B cells and maintenance of the transformed state but not for cytolytic infection of the feather follicle epithelium and horizontal spread of MDV. J. Virol. 2005, 79, 4545–4549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Lupiani, B.; Bajwa, K.; Khan, O.A.; Izumiya, Y.; Reddy, S.M. Role of Marek’s Disease Virus (MDV)-Encoded US3 Serine/Threonine Protein Kinase in Regulating MDV Meq and Cellular CREB Phosphorylation. J. Virol. 2020, 94, e00892-20. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lupiani, B.; Ai-Mahmood, M.; Reddy, S.M. Marek’s disease virus US3 protein kinase phosphorylates chicken HDAC 1 and 2 and regulates viral replication and pathogenesis. PLoS Pathog. 2021, 17, e1009307. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lupiani, B.; Reddy, S.M. Manipulation of Promyelocytic Leukemia Protein Nuclear Bodies by Marek’s Disease Virus Encoded US3 Protein Kinase. Microorganisms 2021, 9, 685. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Bursa/Body Weight × 100 | Thymus/Body Weight × 100 |
|---|---|---|
| Negative | 0.422 ± 0.100 a | 0.741 ± 0.098 a |
| MDV-1 | 0.213 ± 0.079 b | 0.283 ± 0.113 c |
| MDV-1-ΔUS3 | 0.419 ± 0.057 a | 0.495 ± 0.106 b |
| MDV-1-ΔUS3_Rev | 0.141 ± 0.072 b | 0.204 ± 0.050 c |
| MDV-1-MDV-2/US3 | 0.188 ± 0.078 b | 0.426 ± 0.152 b |
| MDV-1-HVT/US3 | 0.391 ± 0.084 a | 0.429 ± 0.091 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Fang, X.; AI-Mahmood, M.; Li, Q.; Lupiani, B.; Reddy, S.M. US3 Serine/Threonine Protein Kinase from MDV-1, MDV-2, and HVT Differentially Regulate Viral Gene Expression and Replication. Microorganisms 2021, 9, 785. https://doi.org/10.3390/microorganisms9040785
Liao Y, Fang X, AI-Mahmood M, Li Q, Lupiani B, Reddy SM. US3 Serine/Threonine Protein Kinase from MDV-1, MDV-2, and HVT Differentially Regulate Viral Gene Expression and Replication. Microorganisms. 2021; 9(4):785. https://doi.org/10.3390/microorganisms9040785
Chicago/Turabian StyleLiao, Yifei, Xin Fang, Mohammad AI-Mahmood, Qinglei Li, Blanca Lupiani, and Sanjay M. Reddy. 2021. "US3 Serine/Threonine Protein Kinase from MDV-1, MDV-2, and HVT Differentially Regulate Viral Gene Expression and Replication" Microorganisms 9, no. 4: 785. https://doi.org/10.3390/microorganisms9040785