
Human Herpesvirus and the Immune Checkpoint PD-1/PD-L1 Pathway: Disorders and Strategies for Survival

{kind=link}
Abstract
:1. Introduction
2. HSV
3. VZV
4. CMV
5. HHV-6A, 6B, and 7
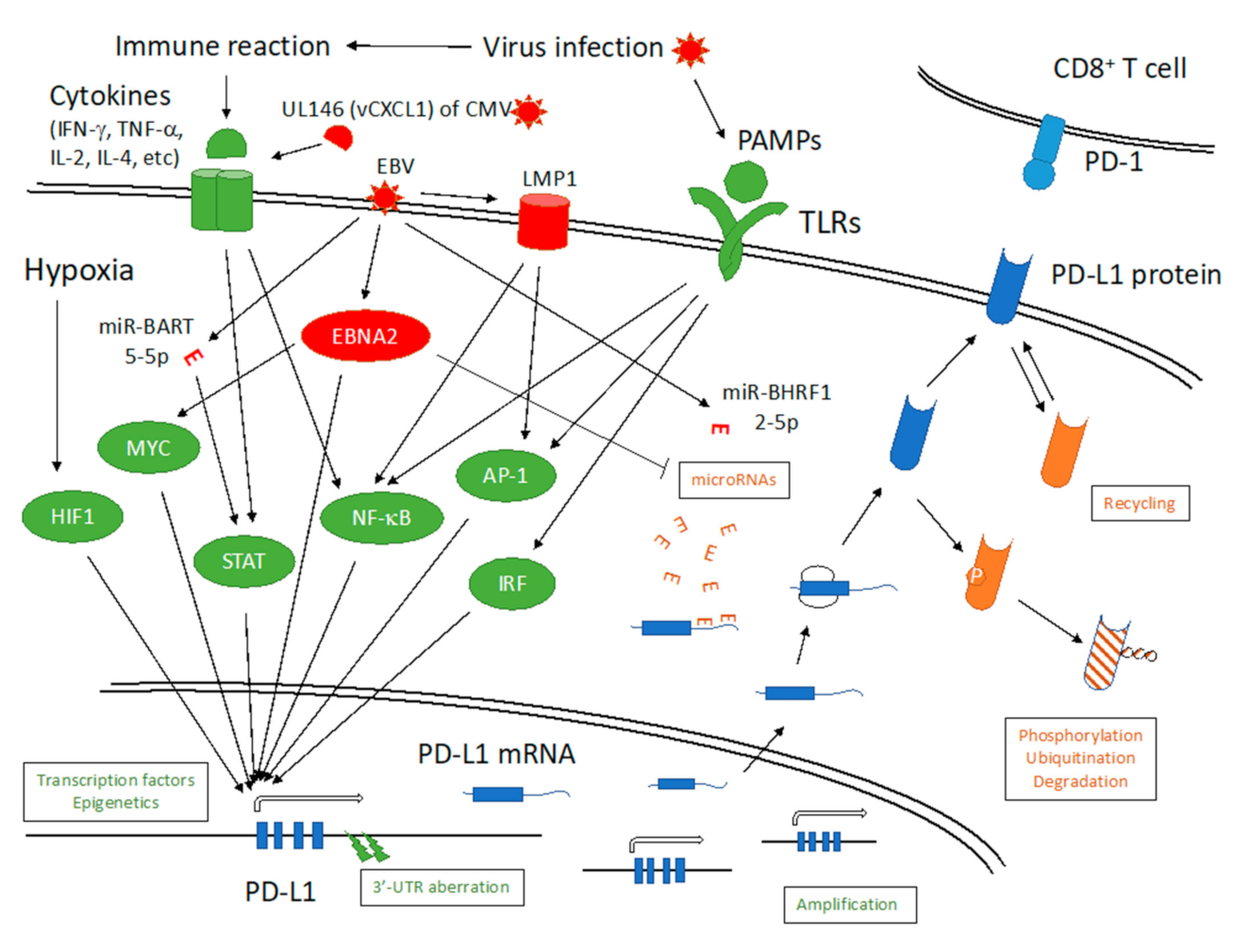
6. EBV
7. KSHV
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Grinde, B. Herpesviruses: Latency and reactivation–viral strategies and host response. J. Oral. Microbiol. 2013, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Knipe, D.M.; Howley, P.M. Fields Virology, 6th ed; Wolters Kluwer/Lippincott Williams & Wil-kins Health: Philadelphia, PA, USA, 2013. [Google Scholar]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2008, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Weinberg, A.; Schmid, D.S. Herpes Simplex Virus and Varicella-Zoster Virus. Microbiol. Spectr. 2016, 4, 135–156. [Google Scholar] [CrossRef]
- Santos, C.A. Cytomegalovirus and other beta-Herpesviruses. Semin. Nephrol. 2016, 36, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Longnecker, R.; Neipel, F. Introduction to the human gamma-herpesviruses. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Sun, C.; Mezzadra, R.; Schumacher, T.N. Regulation and Function of the PD-L1 Checkpoint. Immunity 2018, 48, 434–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestipino, A.; Zeiser, R. Clinical implications of tumor-intrinsic mechanisms regulating PD-L1. Sci. Transl. Med. 2019, 11, eaav4810. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lin, W.; Tang, X.; Li, S.; Guo, L.; Lin, Y.; Kwok, H.F. The Roles of microRNAs in Regulating the Expression of PD-1/PD-L1 Immune Checkpoint. Int. J. Mol. Sci. 2017, 18, 2540. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, H.; Yao, H.; Li, C.; Fang, J.-Y.; Xu, J. Regulation of PD-L1: Emerging Routes for Targeting Tumor Immune Evasion. Front. Pharmacol. 2018, 9, 536. [Google Scholar] [CrossRef]
- Mofrad, M.G.; Maleki, D.T.; Faghihloo, E. The roles of programmed death ligand 1 in virus-associated cancers. Infect. Genet. Evol. 2020, 84, 104368. [Google Scholar] [CrossRef]
- Schönrich, G.; Raftery, M.J. The PD-1/PD-L1 Axis and Virus Infections: A Delicate Balance. Front. Cell. Infect. Microbiol. 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Jun, H.; Seo, S.K.; Jeong, H.-Y.; Seo, H.-M.; Zhu, G.; Chen, L.; Choi, I.-H. B7-H1 (CD274) inhibits the development of herpetic stromal keratitis (HSK). FEBS Lett. 2005, 579, 6259–6264. [Google Scholar] [CrossRef] [Green Version]
- Frank, G.M.; Lepisto, A.J.; Freeman, M.L.; Sheridan, B.S.; Cherpes, T.L.; Hendricks, R.L. Early CD4+T Cell Help Prevents Partial CD8+T Cell Exhaustion and Promotes Maintenance of Herpes Simplex Virus 1 Latency. J. Immunol. 2009, 184, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Hamrah, P.; Gate, D.; Mott, K.R.; Mantopoulos, D.; Zheng, L.; Town, T.; Jones, C.; Von Andrian, U.H.; Freeman, G.J.; et al. The Role of LAT in Increased CD8+ T Cell Exhaustion in Trigeminal Ganglia of Mice Latently Infected with Herpes Simplex Virus 1. J. Virol. 2011, 85, 4184–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chentoufi, A.A.; Kritzer, E.; Tran, M.V.; Dasgupta, G.; Lim, C.H.; David, C.Y.; Afifi, R.E.; Jiang, X.; Carpenter, D.; Osorio, N.; et al. The herpes simplex virus 1 latency-associated transcript promotes functional exhaustion of vi-rus-specific CD8+ T cells in latently infected trigeminal ganglia: A novel immune evasion mechanism. J. Virol. 2011, 85, 9127–9138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant-Hudson, K.M.; Carr, D.J. PD-L1-expressing dendritic cells contribute to viral resistance during acute HSV-1 infection. Clin. Dev. Immunol. 2012, 2012, 924619. [Google Scholar] [CrossRef] [PubMed]
- Channappanavar, R.; Twardy, B.S.; Suvas, S. Blocking of PDL-1 Interaction Enhances Primary and Secondary CD8 T Cell Response to Herpes Simplex Virus-1 Infection. PLoS ONE 2012, 7, e39757. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Rowe, A.M.; Carroll, K.L.; Harvey, S.A.K.; Hendricks, R.L. PD-L1/B7-H1 Inhibits Viral Clearance by Macrophages in HSV-1–Infected Corneas. J. Immunol. 2018, 200, 3711–3719. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Wang, P.-Y.; Hutzen, B.; Sprague, L.; Swain, H.M.; Love, J.K.; Stanek, J.R.; Boon, L.; Conner, J.; Cripe, T.P. Cooperation of Oncolytic Herpes Virotherapy and PD-1 Blockade in Murine Rhabdomyosarcoma Models. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Saha, D.; Martuza, R.L.; Rabkin, S.D. Macrophage Polarization Contributes to Glioblastoma Eradication by Combination Immunovirotherapy and Immune Checkpoint Blockade. Cancer Cell 2017, 32, 253–267.e5. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Tateishi, R.; Iwai, M.; Koike, K.; Todo, T. Neoadjuvant use of oncolytic herpes virus G47Delta enhances the antitumor efficacy of radiofrequency ablation. Mol. Ther. Oncolytics 2020, 18, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.; Blackmon, A.; Neff, C.P.; Palmer, B.E.; Gilden, D.; Badani, H.; Nagel, M.A. Varicella-Zoster Virus Downregulates Programmed Death Ligand 1 and Major Histocompatibility Complex Class I in Human Brain Vascular Adventitial Fibroblasts, Perineurial Cells, and Lung Fibroblasts. J. Virol. 2016, 90, 10527–10534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.; Como, C.N.; Jing, L.; Blackmon, A.; Neff, C.P.; Krueger, O.; Bubak, A.N.; Palmer, B.E.; Koelle, D.M.; Nagel, M.A. Varicella zoster virus productively infects human peripheral blood mononuclear cells to modulate expres-sion of immunoinhibitory proteins and blocking PD-L1 enhances virus-specific CD8+ T cell effector function. PLoS Pathog. 2019, 15, e1007650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, R.; Shirai, T.; Namkoong, H.; Zhang, H.; Berry, G.J.; Wallis, B.B.; Schaefgen, B.; Harrison, D.G.; Tremmel, J.A.; Giacomini, J.C.; et al. Pyruvate controls the checkpoint inhibitor PD-L1 and suppresses T cell immunity. J. Clin. Investig. 2017, 127, 2725–2738. [Google Scholar] [CrossRef]
- Benedict, C.A.; Loewendorf, A.; Garcia, Z.; Blazar, B.R.; Janssen, E.M. Dendritic Cell Programming by Cytomegalovirus Stunts Naive T Cell Responses via the PD-L1/PD-1 Pathway. J. Immunol. 2008, 180, 4836–4847. [Google Scholar] [CrossRef] [Green Version]
- Sester, U.; Presser, D.; Dirks, J.; Gärtner, B.C.; Köhler, H.; Sester, M. PD-1 Expression and IL-2 Loss of Cytomegalovirus- Specific T Cells Correlates with Viremia and Reversible Functional Anergy. Arab. Archaeol. Epigr. 2008, 8, 1486–1497. [Google Scholar] [CrossRef]
- Hu, L.; Wen, Z.; Chen, J.; Chen, Y.; Jin, L.; Shi, H.; Chen, J.; Chen, J. The cytomegalovirus UL146 gene product vCXCL1 promotes the resistance of hepatic cells to CD8+ T cells through up-regulation of PD-L1. Biochem. Biophys. Res. Commun. 2020, 532, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Romeo, M.A.; Montani, M.S.G.; Benedetti, R.; Giambelli, L.; D’Aprile, R.; Gaeta, A.; Faggioniab, A.; Cirone, M. The cross-talk between STAT1/STAT3 and ROS up-regulates PD-L1 and promotes the release of pro-inflammatory/immune suppressive cytokines in primary monocytes infected by HHV-6B. Virus Res. 2021, 292, 198231. [Google Scholar] [CrossRef]
- Abe, H.; Kaneda, A.; Fukayama, M. Epstein-Barr Virus-Associated Gastric Carcinoma: Use of Host Cell Machineries and Somatic Gene Mutations. Pathobiology 2015, 82, 212–223. [Google Scholar] [CrossRef]
- Romeo, M.A.; Montani, M.S.G.; Benedetti, R.; Giambelli, L.; D’Aprile, R.; Gaeta, A.; Faggioni, A.; Cirone, M. PD-L1 expression is characteristic of a subset of aggressive B-cell lymphomas and virus-associated malig-nancies. Clin. Cancer Res. 2013, 19, 3462–3473. [Google Scholar]
- Fang, W.; Zhang, J.; Hong, S.; Zhan, J.; Chen, N.; Qin, T.; Zhang, L.; Tang, Y.; Zhang, Y.; Kang, S.; et al. EBV-driven LMP1 and IFN-gamma up-regulate PD-L1 in nasopharyngeal carcinoma: Implications for onco-targeted therapy. Oncotarget 2014, 5, 12189–12202. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Monti, S.; Rodig, S.J.; Juszczynski, P.; Currie, T.; O’Donnell, E.; Chapuy, B.; Takeyama, K.; Neuberg, D.; Golub, T.R.; et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood 2010, 116, 3268–3277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.R.; Rodig, S.; Juszczynski, P.; Ouyang, J.; Sinha, P.; O’Donnell, E.; Neuberg, D.; Shipp, M.A. Constitutive AP-1 Activity and EBV Infection Induce PD-L1 in Hodgkin Lymphomas and Posttransplant Lymphoproliferative Disorders: Implications for Targeted Therapy. Clin. Cancer Res. 2012, 18, 1611–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, K.; Miyoshi, H.; Sakata, S.; Dobashi, A.; Couronné, L.; Kogure, Y.; Sato, Y.; Nishida, K.; Gion, Y.; Shiraishi, Y.; et al. Frequent structural variations involving programmed death ligands in Epstein-Barr virus-associated lymphomas. Leukemia 2019, 33, 1687–1699. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.; Kim, S.; Kim, P.-J.; Go, H.; Nam, S.J.; Paik, J.H.; Kim, Y.A.; Kim, T.M.; Heo, D.S.; Kim, C.W.; et al. Clinicopathological analysis of programmed cell death 1 and programmed cell death ligand 1 expression in the tumour microenvironments of diffuse large B cell lymphomas. Histopathology 2015, 68, 1079–1089. [Google Scholar] [CrossRef]
- Okuno, Y.; Murata, T.; Sato, Y.; Muramatsu, H.; Ito, Y.; Watanabe, T.; Okuno, T.; Murakami, N.; Yoshida, K.; Sawada, A.; et al. Defective Epstein–Barr virus in chronic active infection and haematological malignancy. Nat. Microbiol. 2019, 4, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Nishikawa, J.; Sakai, K.; Iizasa, H.; Yoshiyama, H.; Yanagihara, M.; Shuto, T.; Shimokuri, K.; Kanda, T.; Suehiro, Y.; et al. EBV-associated gastric cancer evades T-cell immunity by PD-1/PD-L1 interactions. Gastric Cancer 2019, 22, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Veloza, L.; Teixido, C.; Castrejon, N.; Climent, F.; Carrió, A.; Marginet, M.; Martinez, A. Clinicopathological evaluation of the programmed cell death 1 (PD1)/programmed cell death-ligand 1 (PD-L1) axis in post-transplant lymphoproliferative disorders: Association with Epstein-Barr virus, PD-L1 copy number al-terations, and outcome. Histopathology 2019, 75, 799–812. [Google Scholar] [CrossRef]
- Xue, T.; Wang, W.-G.; Zhou, X.-Y.; Li, X.-Q. EBV-positive diffuse large B-cell lymphoma features PD-L1 protein but not mRNA overexpression. Pathology 2018, 50, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.; Patel, S.P.; Kurzrock, A.G.S.P.P.R. PD-1–PD-L1 immune-checkpoint blockade in B-cell lymphomas. Nat. Rev. Clin. Oncol. 2017, 14, 203–220. [Google Scholar] [CrossRef]
- Johnson, D.; Ma, B.B.Y. Targeting the PD-1/PD-L1 interaction in nasopharyngeal carcinoma. Oral. Oncol. 2021, 113, 105–127. [Google Scholar] [CrossRef]
- Joshi, S.S.; Maron, S.B.; Catenacci, D.V. Pembrolizumab for treatment of advanced gastric and gastroesophageal junction adenocarcinoma. Future Oncol. 2018, 14, 417–430. [Google Scholar] [CrossRef]
- Merryman, R.W.; Armand, P.; Wright, K.T.; Rodig, S.J. Checkpoint blockade in Hodgkin and non-Hodgkin lymphoma. Blood Adv. 2017, 1, 2643–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, R. NK/T Cell Lymphoma: Updates in Therapy. Curr. Hematol. Malign-Rep. 2018, 13, 7–12. [Google Scholar] [CrossRef]
- Takahara, T.; Satou, A.; Ishikawa, E.; Kohno, K.; Kato, S.; Suzuki, Y.; Takahashi, E.; Ohashi, A.; Asano, N.; Tsuzuki, T.; et al. Clinicopathological analysis of neoplastic PD-L1-positive EBV+ diffuse large B cell lymphoma, not otherwise specified, in a Japanese cohort. Virchows Archiv. 2021, 478, 541–552. [Google Scholar] [CrossRef]
- Bi, X.W.; Wang, H.; Zhang, W.W.; Wang, J.H.; Liu, W.J.; Xia, Z.-J.; Huang, H.; Jiang, W.; Zhang, Y.; Wang, L. PD-L1 is upregulated by EBV-driven LMP1 through NF-kappaB pathway and correlates with poor prognosis in natural killer/T-cell lymphoma. J. Hematol. Oncol. 2016, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Anastasiadou, E.; Stroopinsky, D.; Alimperti, S.; Jiao, A.L.; Pyzer, A.R.; Cippitelli, C.; Pepe, G.; Severa, M.; Rosenblatt, J.; Etna, M.P.; et al. Epstein−Barr virus-encoded EBNA2 alters immune checkpoint PD-L1 expression by downregulating miR-34a in B-cell lymphomas. Leukemia 2019, 33, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Yanagi, Y.; Okuno, Y.; Narita, Y.; Al Masud, H.A.; Watanabe, T.; Sato, Y.; Kanda, T.; Kimura, H.; Murata, T. RNAseq analysis identifies involvement of EBNA2 in PD-L1 induction during Epstein-Barr virus infection of primary B cells. Virology 2021, 557, 44–54. [Google Scholar] [CrossRef]
- Kataoka, K.; Shiraishi, Y.; Takeda, Y.; Sakata, S.; Matsumoto, M.; Nagano, S.; Maeda, T.; Nagata, Y.; Kitanaka, A.; Mizuno, S.; et al. Aberrant PD-L1 expression through 3′-UTR disruption in multiple cancers. Nature 2016, 534, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Yoshida, K.; Suzuki, Y.; Iriyama, C.; Inoue, Y.; Sanada, M.; Kiyoi, H. Frequent genetic alterations in immune checkpoint-related genes in intravascular large B-cell lympho-ma. Blood 2020, 137, 1491–1502. [Google Scholar] [CrossRef]
- Cristino, A.S.; Nourse, J.; West, R.A.; Sabdia, M.B.; Law, S.C.; Gunawardana, J.; Gandhi, M.K. EBV microRNA-BHRF1-2-5p targets the 3’UTR of immune checkpoint ligands PD-L1 and PD-L2. Blood 2019, 134, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.J.; Chang, M.S.; Kim, D.H.; Kim, W.; Koo, B.K.; Yun, S.C.; Kim, S.H.; Kim, Y.S.; Woo, J.H. Epstein-Barr virus-encoded miR-BART5-5p upregulates PD-L1 through PIAS3/pSTAT3 modulation, wors-ening clinical outcomes of PD-L1-positive gastric carcinomas. Gastric Cancer 2020, 23, 780–795. [Google Scholar] [CrossRef] [PubMed]
- Ronaghy, A.; Wang, H.-Y.; Thorson, J.A.; Medeiros, L.J.; Xie, Y.; Zhang, X.; Sheikh-Fayyaz, S. PD-L1 and Notch1 expression in KSHV/HHV-8 and EBV associated germinotropic lymphoproliferative disorder: Case report and review of the literature. Pathology 2017, 49, 430–435. [Google Scholar] [CrossRef]
- Sanchez, S.; Veloza, L.; Wang, L.; López, M.; López-Guillermo, A.; Marginet, M.; Martínez, A.; Balagué, O.; Campo, E. HHV8-positive, EBV-positive Hodgkin lymphoma-like large B cell lymphoma: Expanding the spectrum of HHV8 and EBV-associated lymphoproliferative disorders. Int. J. Hematol. 2020, 112, 734–740. [Google Scholar] [CrossRef]
- Chen, J.; Del Valle, L.; Lin, H.Y.; Plaisance-Bonstaff, K.; Forrest, J.C.; Post, S.R.; Qin, Z. Expression of PD-1 and PD-Ls in Kaposi’s sarcoma and regulation by oncogenic herpesvirus lytic reactiva-tion. Virology 2019, 536, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Montani, M.S.G.; Falcinelli, L.; Santarelli, R.; Granato, M.; Romeo, M.A.; Cecere, N.; Gonnella, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. KSHV infection skews macrophage polarisation towards M2-like/TAM and activates Ire1 α-XBP1 axis up-regulating pro-tumorigenic cytokine release and PD-L1 expression. Br. J. Cancer 2020, 123, 298–306. [Google Scholar] [CrossRef]
- Host, K.M.; Jacobs, S.R.; West, J.A.; Zhang, Z.; Costantini, L.M.; Stopford, C.M.; Dittmer, D.P.; Damania, B. Kaposi’s Sarcoma-Associated Herpesvirus Increases PD-L1 and Proinflammatory Cytokine Expression in Human Monocytes. mBio 2017, 8, e00917-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murata, T. Human Herpesvirus and the Immune Checkpoint PD-1/PD-L1 Pathway: Disorders and Strategies for Survival. Microorganisms 2021, 9, 778. https://doi.org/10.3390/microorganisms9040778
Murata T. Human Herpesvirus and the Immune Checkpoint PD-1/PD-L1 Pathway: Disorders and Strategies for Survival. Microorganisms. 2021; 9(4):778. https://doi.org/10.3390/microorganisms9040778
Chicago/Turabian StyleMurata, Takayuki. 2021. "Human Herpesvirus and the Immune Checkpoint PD-1/PD-L1 Pathway: Disorders and Strategies for Survival" Microorganisms 9, no. 4: 778. https://doi.org/10.3390/microorganisms9040778