
Human Endogenous Retroviruses in Glioblastoma Multiforme

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
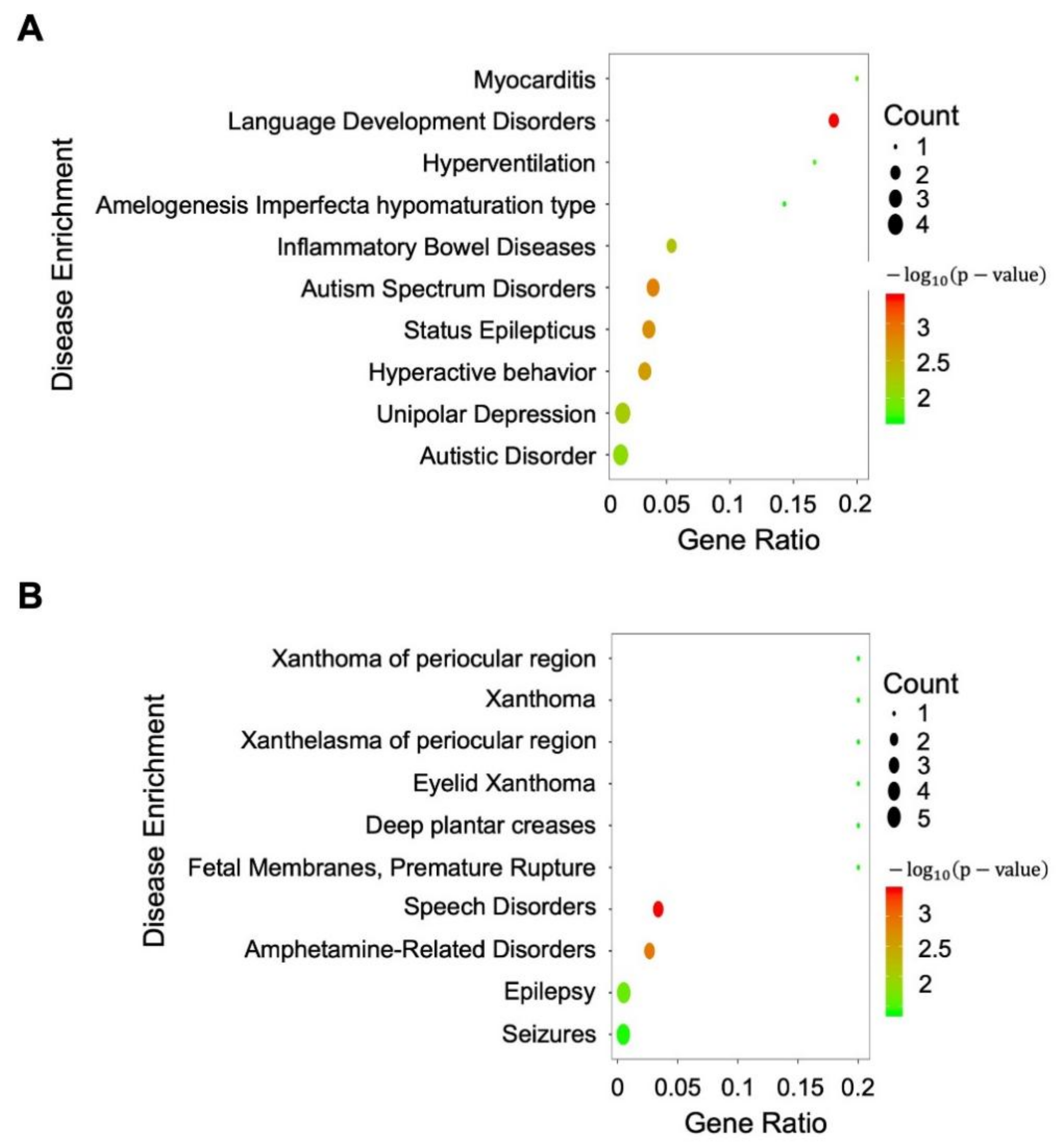
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2008–2012. Neuro-Oncology 2015, 17 (Suppl. 4), iv1–iv62. [Google Scholar] [CrossRef] [PubMed]
- Ohgaki, H.; Kleihues, P. Epidemiology and etiology of gliomas. Acta Neuropathol. 2005, 109, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.; LaBombardi, V.; Green, S.; Pessin-Minsley, M.S.; Germano, I.M.; Rosenzweig, K.E. No circulating cytomegalovirus in five patients with glioblastoma multiforme. Anticancer Res. 2011, 31, 959–960. [Google Scholar] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011–2015. Neuro-Oncology 2018, 20 (Suppl. 4), iv1–iv86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, J.L.; Schwartzbaum, J.A.; Wrensch, M.; Wiemels, J.L. Epidemiology of brain tumors. Neurol. Clin. 2007, 25, 867–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchana, M.; Margaliot, M.; Liphshitz, I. Changes in Brain Glioma Incidence and Laterality Correlates with Use of Mobile Phones–A Nationwide Population Based Study in Israel. Asian Pac. J. Cancer Prev. 2012, 13, 5857–5863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deltour, I.; Auvinen, A.; Feychting, M.; Johansen, C.; Klaeboe, L.; Sankila, R.; Schüz, J. Mobile Phone Use and Incidence of Glioma in the Nordic Countries 1979–2008: Consistency Check. Epidemiology 2012, 23, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Little, M.P.; Azizova, T.V.; Bazyka, D.; Bouffler, S.D.; Cardis, E.; Chekin, S.; Chumak, V.V.; Cucinotta, F.A.; de Vathaire, F.; Hall, P. Systematic review and meta-analysis of circulatory disease from exposure to low-level ionizing radiation and estimates of potential population mortality risks. Environ. Health Perspect. 2012, 120, 1503–1511. [Google Scholar] [CrossRef] [Green Version]
- Cobbs, C.S.; Harkins, L.; Samanta, M.; Gillespie, G.Y.; Bharara, S.; King, P.H.; Nabors, L.B.; Cobbs, C.G.; Britt, W.J. Human cytomegalovirus infection and expression in human malignant glioma. Cancer Res. 2002, 62, 3347–3350. [Google Scholar]
- Wrensch, M.; Minn, Y.; Chew, T.; Bondy, M.; Berger, M.S. Epidemiology of primary brain tumors: Current concepts and review of the literature. Neuro-Oncology 2002, 4, 278–299. [Google Scholar] [CrossRef]
- Saddawi-Konefka, R.; Crawford, J.R. Chronic Viral Infection and Primary Central Nervous System Malignancy. J. Neuroimmune Pharmacol. 2010, 5, 387–403. [Google Scholar] [CrossRef] [Green Version]
- Alexander, B.M.; Cloughesy, T.F. Adult glioblastoma. J. Clin. Oncol. 2017, 35, 2402–2409. [Google Scholar] [CrossRef]
- Hochhalter, C.B.; Carr, C.; O’Neill, B.E.; Ware, M.L.; Strong, M.J. The association between human cytomegalovirus and glioblastomas: A review. Neuroimmunol. Neuroinflammation 2017, 4, 96. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.; Xia, Y.; Bettegowda, C.; Weller, M. Current state of immunotherapy for glioblastoma. Nat. Rev. Clin. Oncol. 2018, 15, 422–442. [Google Scholar] [CrossRef]
- Nelson, P.N.; Carnegie, P.; Martin, J.; Ejtehadi, H.D.; Hooley, P.; Roden, D.; Rowland-Jones, S.; Warren, P.; Astley, J.; Murray, P.G. Demystified… Human endogenous retroviruses. Mol. Pathol. 2003, 56, 11. [Google Scholar] [CrossRef] [Green Version]
- Kojima, K.K. Human transposable elements in Repbase: Genomic footprints from fish to humans. Mob. DNA 2018, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S. Initial sequencing and analysis of the human genome. International Human Genome Sequencing Consortium. Nature 2001, 409, 860–921. [Google Scholar]
- Cegolon, L.; Salata, C.; Weiderpass, E.; Vineis, P.; Palù, G.; Mastrangelo, G. Human endogenous retroviruses and cancer prevention: Evidence and prospects. BMC Cancer 2013, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Bannert, N.; Kurth, R. Retroelements and the human genome: New perspectives on an old relation. Proc. Natl. Acad. Sci. USA 2004, 101 (Suppl. 2), 14572–14579. [Google Scholar] [CrossRef] [Green Version]
- Mager, D.; Stoye, J. Mammalian endogenous retroviruses. Microbiol. Spectr. 2015, 3, MDNA3-0009-2014. [Google Scholar] [CrossRef] [Green Version]
- Gilboa, E.; Mitra, S.W.; Goff, S.; Baltimore, D. A detailed model of reverse transcription and tests of crucial aspects. Cell 1979, 18, 93–100. [Google Scholar] [CrossRef]
- Swanstrom, R.; DeLorbe, W.J.; Bishop, J.M.; Varmus, H.E. Nucleotide sequence of cloned unintegrated avian sarcoma virus DNA: Viral DNA contains direct and inverted repeats similar to those in transposable elements. Proc. Natl. Acad. Sci. USA 1981, 78, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; De Crombrugghe, B.; Pastan, I. Identification of a functional promoter in the long terminal repeat of Rous sarcoma virus. Cell 1980, 22, 787–797. [Google Scholar] [CrossRef]
- Jern, P.; Coffin, J.M. Effects of Retroviruses on Host Genome Function. Annu. Rev. Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef] [Green Version]
- Leib-Mösch, C.; Seifarth, W.; Schön, U. Influence of human endogenous retroviruses on cellular gene expression. In Retroviruses and Primate Genome Evolution; Sverdlov, E., Ed.; Landes Bioscience: Georgetown, TX, USA, 2004; pp. 124–145. [Google Scholar]
- Medstrand, P.; Landry, J.-R.; Mager, D.L. Long Terminal Repeats Are Used as Alternative Promoters for the Endothelin B Receptor and Apolipoprotein C-I Genes in Humans. J. Biol. Chem. 2001, 276, 1896–1903. [Google Scholar] [CrossRef] [Green Version]
- Boller, K.; Schönfeld, K.; Lischer, S.; Fischer, N.; Hoffmann, A.; Kurth, R.; Tönjes, R.R. Human endogenous retrovirus HERV-K113 is capable of producing intact viral particles. J. Gen. Virol. 2008, 89, 567–572. [Google Scholar] [CrossRef]
- Faff, O.; Murray, A.B.; Schmidt, J.; Leib-Mösch, C.; Erfle, V.; Hehlmann, R. Retrovirus-like particles from the human T47D cell line are related to mouse mammary tumour virus and are of human endogenous origin. J. Gen. Virol. 1992, 73, 1087–1097. [Google Scholar] [CrossRef]
- Andersson, A.C.; Svensson, A.C.; Rolny, C.; Andersson, G.; Larsson, E. Expression of human endogenous retrovirus ERV3 (HERV-R) mRNA in normal and neoplastic tissues. Int. J. Oncol. 1998, 12, 309–322. [Google Scholar] [CrossRef]
- Löwer, R.; Löwer, J.; Tondera-Koch, C.; Kurth, R. A General Method for the Identification of Transcribed Retrovirus Sequences (R-U5 PCR) Reveals the Expression of the Human Endogenous Retrovirus Loci HERV-H and HERV-K in Teratocarcinoma Cells. Virology 1993, 192, 501–511. [Google Scholar] [CrossRef]
- Larsson, E.; Andersson, G. Beneficial Role of Human Endogenous Retroviruses: Facts and Hypotheses. Scand. J. Immunol. 1998, 48, 329–338. [Google Scholar] [CrossRef]
- Lindeskog, M.; Medstrand, P.; Blomberg, J. Sequence variation of human endogenous retrovirus ERV9-related elements in an env region corresponding to an immunosuppressive peptide: Transcription in normal and neoplastic cells. J. Virol. 1993, 67, 1122–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauter, M.; Roemer, K.; Best, B.; Afting, M.; Schommer, S.; Seitz, G.; Hartmann, M.; Mueller-Lantzsch, N. Specificity of antibodies directed against Env protein of human endogenous retroviruses in patients with germ cell tumors. Cancer Res. 1996, 56, 4362–4365. [Google Scholar] [PubMed]
- Sauter, M.; Schommer, S.; Kremmer, E.; Remberger, K.; Dölken, G.; Lemm, I.; Buck, M.; Best, B.; Neumann-Haefelin, D.; Mueller-Lantzsch, N. Human endogenous retrovirus K10: Expression of Gag protein and detection of antibodies in patients with seminomas. J. Virol. 1995, 69, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavetti, F.; Thonnard, J.; Colau, D.; Boon, T.; Coulie, P.G. A human endogenous retroviral sequence encoding an antigen recognized on melanoma by cytolytic T lymphocytes. Cancer Res. 2002, 62, 5510–5516. [Google Scholar] [PubMed]
- Contreras-Galindo, R.; Kaplan, M.H.; Leissner, P.; Verjat, T.; Ferlenghi, I.; Bagnoli, F.; Giusti, F.; Dosik, M.H.; Hayes, D.F.; Gitlin, S.D.; et al. Human Endogenous Retrovirus K (HML-2) Elements in the Plasma of People with Lymphoma and Breast Cancer. J. Virol. 2008, 82, 9329–9336. [Google Scholar] [CrossRef] [Green Version]
- Pichon, J.-P.; Bonnaud, B.; Cleuziat, P.; Mallet, F. Multiplex degenerate PCR coupled with an oligo sorbent array for human endogenous retrovirus expression profiling. Nucleic Acids Res. 2006, 34, e46. [Google Scholar] [CrossRef] [Green Version]
- Wang-Johanning, F.; Liu, J.; Rycaj, K.; Huang, M.; Tsai, K.; Rosen, D.G.; Chen, D.-T.; Lu, D.W.; Barnhart, K.F.; Johanning, G.L. Expression of multiple human endogenous retrovirus surface envelope proteins in ovarian cancer. Int. J. Cancer 2006, 120, 81–90. [Google Scholar] [CrossRef]
- Wang-Johanning, F.; Frost, A.R.; Jian, B.; Azerou, R.; Lu, D.W.; Chen, D.T.; Johanning, G.L. Detecting the expression of human endogenous retrovirus E envelope transcripts in human prostate adenocarcinoma. Cancer 2003, 98, 187–197. [Google Scholar] [CrossRef]
- Gimenez, J.; Montgiraud, C.; Pichon, J.-P.; Bonnaud, B.; Arsac, M.; Ruel, K.; Bouton, O.; Mallet, F. Custom human endogenous retroviruses dedicated microarray identifies self-induced HERV-W family elements reactivated in testicular cancer upon methylation control. Nucleic Acids Res. 2010, 38, 2229–2246. [Google Scholar] [CrossRef]
- Pérot, P.; Mullins, C.S.; Naville, M.; Bressan, C.; Hühns, M.; Gock, M.; Kuhn, F.; Volff, J.-N.; Trillet-Lenoir, V.; Linnebacher, M.; et al. Expression of young HERV-H loci in the course of colorectal carcinoma and correlation with molecular subtypes. Oncotarget 2015, 6, 40095–40111. [Google Scholar] [CrossRef] [Green Version]
- Perron, H.; Geny, C.; Laurent, A.; Mouriquand, C.; Pellat, J.; Perret, J.; Seigneurin, J. Leptomeningeal cell line from multiple sclerosis with reverse transcriptase activity and viral particles. Res. Virol. 1989, 140, 551–561. [Google Scholar] [CrossRef]
- McCormick, A.L.; Brown, R.H.; Cudkowicz, M.E.; Al-Chalabi, A.; Garson, J.A. Quantification of reverse transcriptase in ALS and elimination of a novel retroviral candidate. Neurology 2008, 70, 278–283. [Google Scholar] [CrossRef]
- Yolken, R.H.; Karlsson, H.; Yee, F.; Johnston-Wilson, N.; Torrey, E. Endogenous retroviruses and schizophrenia. Brain Res. Rev. 2000, 31, 193–199. [Google Scholar] [CrossRef]
- Singh, S.K. Endogenous retroviruses: Suspects in the disease world. Futur. Microbiol. 2007, 2, 269–275. [Google Scholar] [CrossRef]
- Gruchot, J.; Kremer, D.; Küry, P. Neural Cell Responses Upon Exposure to Human Endogenous Retroviruses. Front. Genet. 2019, 10, 655. [Google Scholar] [CrossRef]
- Díaz-Carballo, D.; Klein, J.; Acikelli, A.H.; Wilk, C.; Saka, S.; Jastrow, H.; Wennemuth, G.; Dammann, P.; Giger-Pabst, U.; Khosrawipour, V.; et al. Cytotoxic stress induces transfer of mitochondria-associated human endogenous retroviral RNA and proteins between cancer cells. Oncotarget 2017, 8, 95945–95964. [Google Scholar] [CrossRef] [Green Version]
- Diem, O.; Schäffner, M.; Seifarth, W.; Leib-Mösch, C. Influence of antipsychotic drugs on human endogenous retrovirus (HERV) transcription in brain cells. PLoS ONE 2012, 7, e30054. [Google Scholar] [CrossRef] [Green Version]
- Flockerzi, A.; Ruggieri, A.; Frank, O.; Sauter, M.; Maldener, E.; Kopper, B.; Wullich, B.; Seifarth, W.; Müller-Lantzsch, N.; Leib-Mösch, C. Expression patterns of transcribed human endogenous retrovirus HERV-K (HML-2) loci in human tissues and the need for a HERV Transcriptome Project. BMC Genom. 2008, 9, 354. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.F.; Wiesner, M.; Denner, J.; Kämmerer, U.; Vince, G.H.; Linsenmann, T.; Löhr, M.; Ernestus, R.I.; Hagemann, C. Expression-analysis of the human endogenous retrovirus HERV-K in human astrocytic tumors. BMC Res. Notes 2014, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zheng, W.J. Informatics, data science, and artificial intelligence. JAMA 2018, 320, 1103–1104. [Google Scholar] [CrossRef]
- Criscione, S.W.; Zhang, Y.; Thompson, W.; Sedivy, J.M.; Neretti, N. Transcriptional landscape of repetitive elements in normal and cancer human cells. BMC Genom. 2014, 15, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, G.J.; Forrest, A.R.; Chalk, A.M.; Schroder, K.; Hayashizaki, Y.; Carninci, P.; Hume, D.A.; Grimmond, S.M. A rescue strategy for multimapping short sequence tags refines surveys of transcriptional activity by CAGE. Genomics 2008, 91, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huda, A.; Lunyak, V.V.; Jordan, I.K. A Gibbs sampling strategy applied to the mapping of ambiguous short-sequence tags. Bioinformatics 2010, 26, 2501–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Law, C.W.; Alhamdoosh, M.; Su, S.; Dong, X.; Tian, L.; Smyth, G.K.; Ritchie, M.E. RNA-Seq analysis is easy as 1-2-3 with limma, Glimma and edgeR. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Morpheus. Available online: https://software.broadinstitute.org/morpheus (accessed on 27 February 2021).
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, A.D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Anand, L. chromoMap: An R package for Interactive Visualization and Annotation of Chromosomes. bioRxiv 2019, 605600. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Jang, I.; Choi, J.; Kim, M.-S.; Lee, A.J.; Kim, H.; Eom, J.; Kim, D.; Jung, I.; Lee, B. 3DIV: A 3D-genome Interaction Viewer and database. Nucleic Acids Res. 2018, 46, D52–D57. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2019, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Lättekivi, F.; Kõks, S.; Keermann, M.; Reimann, E.; Prans, E.; Abram, K.; Silm, H.; Kõks, G.; Kingo, K. Transcriptional landscape of human endogenous retroviruses (HERVs) and other repetitive elements in psoriatic skin. Sci. Rep. 2018, 8, 4358. [Google Scholar] [CrossRef] [Green Version]
- Ho, X.D.; Nguyen, H.G.; Trinh, L.H.; Reimann, E.; Prans, E.; Kõks, G.; Maasalu, K.; Le, V.Q.; Nguyen, V.H.; Le, N.T.N.; et al. Analysis of the Expression of Repetitive DNA Elements in Osteosarcoma. Front. Genet. 2017, 8, 193. [Google Scholar] [CrossRef] [Green Version]
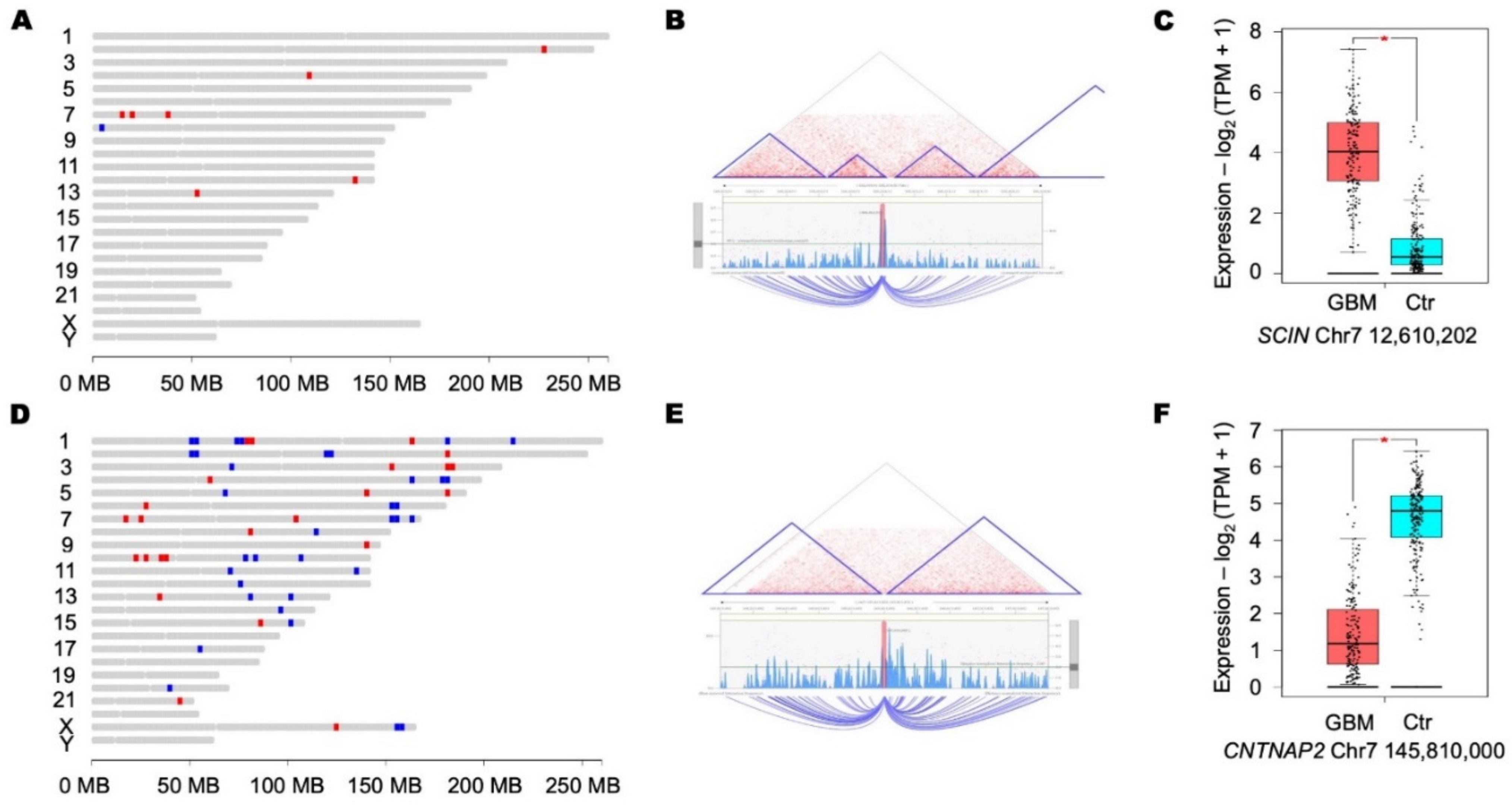
- Bralten, L.B.C.; Gravendeel, A.M.; Kloosterhof, N.K.; Sacchetti, A.; Vrijenhoek, T.; Veltman, J.A.; Bent, M.J.V.D.; Kros, J.M.; Hoogenraad, C.C.; Smitt, P.A.E.S.; et al. The CASPR2 cell adhesion molecule functions as a tumor suppressor gene in glioma. Oncogene 2010, 29, 6138–6148. [Google Scholar] [CrossRef] [Green Version]
- Sakthikumar, S.; Roy, A.; Haseeb, L.; Pettersson, M.E.; Sundström, E.; Marinescu, V.D.; Lindblad-Toh, K.; Forsberg-Nilsson, K. Whole-genome sequencing of glioblastoma reveals enrichment of non-coding constraint mutations in known and novel genes. Genome Biol. 2020, 21, 1–22. [Google Scholar] [CrossRef]
- Derks, J.; Reijneveld, J.C.; Douw, L. Neural network alterations underlie cognitive deficits in brain tumor patients. Curr. Opin. Oncol. 2014, 26, 627–633. [Google Scholar] [CrossRef]
- Heimans, J.J.; Reijneveld, J.C. Factors affecting the cerebral network in brain tumor patients. J. Neuro-Oncol. 2012, 108, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.N.; Bieniasz, P.D. Reconstitution of an infectious human endogenous retrovirus. PLoS Pathog. 2007, 3, e10. [Google Scholar] [CrossRef] [Green Version]
- Guenette, S.Y.; Chen, J.; Jondro, P.D.; Tanzi, R.E. Association of a novel human FE65-like protein with the cytoplasmic domain of the beta-amyloid precursor protein. Proc. Natl. Acad. Sci. USA 1996, 93, 10832–10837. [Google Scholar] [CrossRef] [Green Version]
- Solans, A.; Estivill, X.; De La Luna, S. A new aspartyl protease on 21q22.3, BACE2, is highly similar to Alzheimer’s amyloid precursor protein beta-secretase. Cytogenet. Cell Genet. 2000, 89, 177–184. [Google Scholar] [CrossRef]
- Camporesi, E.; Lashley, T.; Gobom, J.; Lantero-Rodriguez, J.; Hansson, O.; Zetterberg, H.; Blennow, K.; Becker, B. Neuroligin-1 in brain and CSF of neurodegenerative disorders: Investigation for synaptic biomarkers. Acta Neuropathol. Commun. 2021, 9, 19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
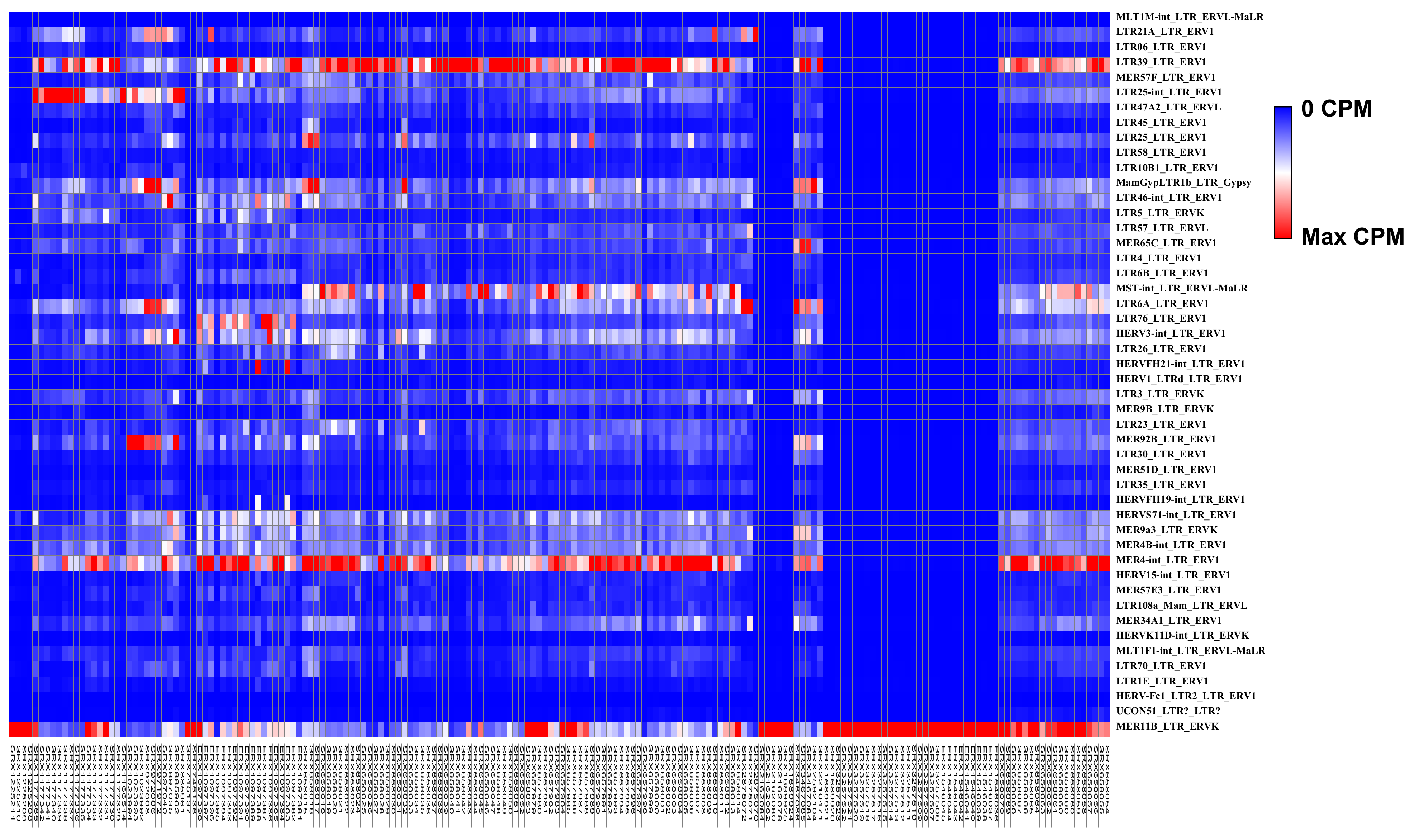
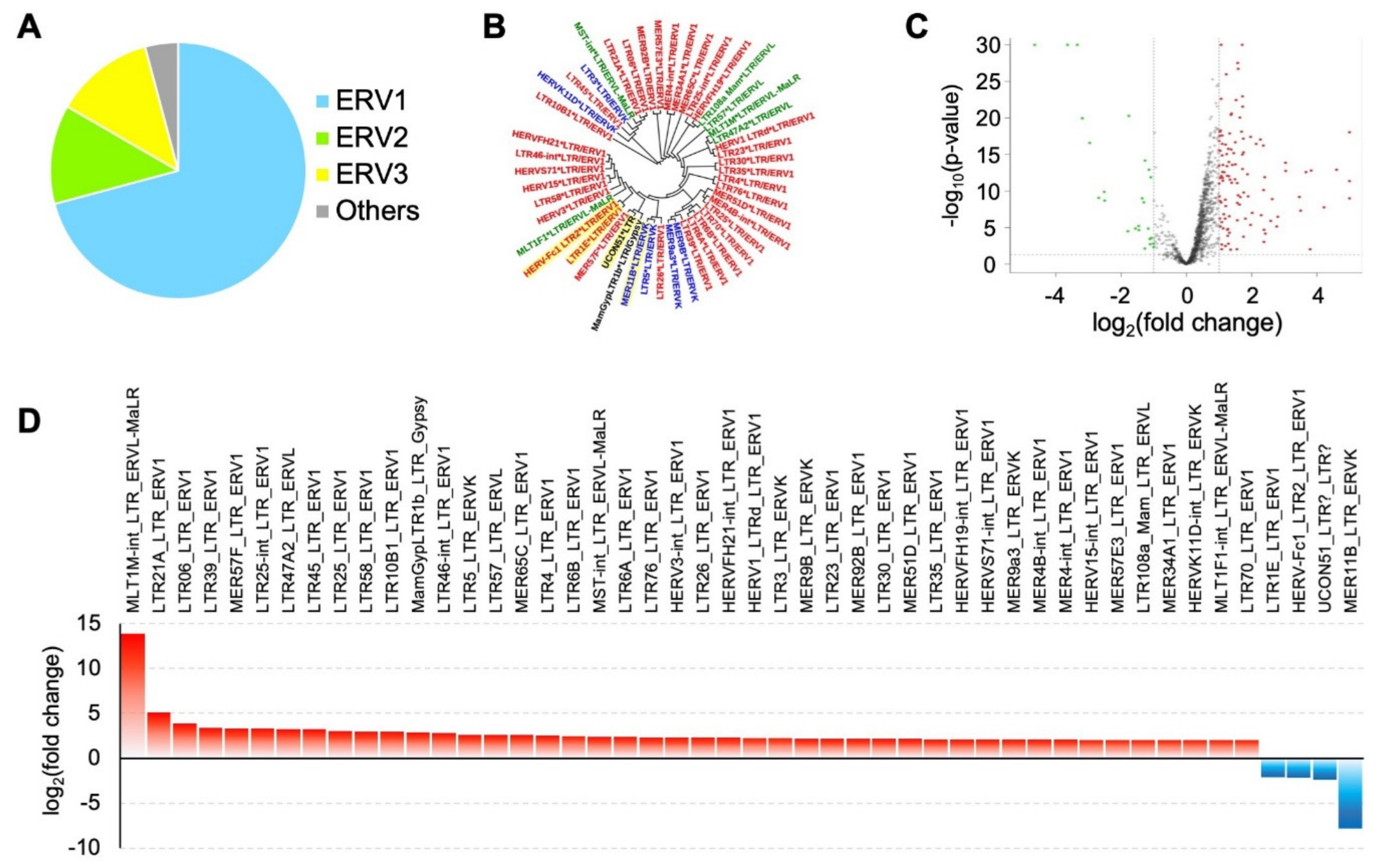
| LTR Elements | Superfamily | NB Mean CPM | GBM Mean CPM | Fold Changes GBM vs. NB |
|---|---|---|---|---|
| MLT1M-int_LTR_ERVL-MaLR | ERV3 | 0 | 0.03 | 13.85 |
| LTR21A_LTR_ERV1 | ERV1 | 48.57 | 249.19 | 5.13 |
| LTR06_LTR_ERV1 | ERV1 | 9.29 | 36.37 | 3.91 |
| LTR39_LTR_ERV1 | ERV1 | 281.42 | 947.75 | 3.37 |
| MER57F_LTR_ERV1 | ERV1 | 44.07 | 145.12 | 3.29 |
| LTR25-int_LTR_ERV1 | ERV1 | 98.72 | 324.11 | 3.28 |
| LTR47A2_LTR_ERVL | ERV3 | 26.76 | 87.46 | 3.27 |
| LTR45_LTR_ERV1 | ERV1 | 12.9 | 41.61 | 3.23 |
| LTR25_LTR_ERV1 | ERV1 | 53.83 | 161.48 | 3 |
| LTR58_LTR_ERV1 | ERV1 | 13.2 | 39.2 | 2.97 |
| LTR10B1_LTR_ERV1 | ERV1 | 31.4 | 92.43 | 2.94 |
| MamGypLTR1b_LTR_Gypsy | Gypsy | 106.56 | 305.69 | 2.87 |
| LTR46-int_LTR_ERV1 | ERV1 | 70.41 | 199.2 | 2.83 |
| LTR5_LTR_ERVK | ERV2 | 32.82 | 85.7 | 2.61 |
| LTR57_LTR_ERVL | ERV3 | 49.21 | 127.45 | 2.59 |
| MER65C_LTR_ERV1 | ERV1 | 60.88 | 156.72 | 2.57 |
| LTR4_LTR_ERV1 | ERV1 | 24.56 | 61.49 | 2.5 |
| LTR6B_LTR_ERV1 | ERV1 | 42.14 | 103.63 | 2.46 |
| MST-int LTR ERVL-MaLR | ERV3 | 132.56 | 320 | 2.41 |
| LTR6A_LTR_ERV1 | ERV1 | 137.45 | 325.28 | 2.37 |
| LTR76_LTR_ERV1 | ERV1 | 60.3 | 141.55 | 2.35 |
| HERV3-int_LTR_ERV1 | ERV1 | 118.4 | 275.59 | 2.33 |
| LTR26_LTR_ERV1 | ERV1 | 48.93 | 112.36 | 2.3 |
| HERVFH21-int_LTR_ERV1 | ERV1 | 22.23 | 50.74 | 2.28 |
| HERV1_LTRd_LTR_ERV1 | ERV1 | 6.46 | 14.7 | 2.28 |
| LTR3_LTR_ERVK | ERV2 | 65.17 | 145.43 | 2.23 |
| MER9B_LTR_ERVK | ERV2 | 16.12 | 35.17 | 2.18 |
| LTR23_LTR_ERV1 | ERV1 | 61.05 | 132.71 | 2.17 |
| MER92B_LTR_ERV1 | ERV1 | 96.38 | 209 | 2.17 |
| LTR30_LTR_ERV1 | ERV1 | 29.1 | 62.58 | 2.15 |
| MER51D_LTR_ERV1 | ERV1 | 11.86 | 25.37 | 2.14 |
| LTR35_LTR_ERV1 | ERV1 | 25.63 | 54.69 | 2.13 |
| HERVFH19-int_LTR_ERV1 | ERV1 | 10.32 | 21.85 | 2.12 |
| HERVS71-int_LTR_ERV1 | ERV1 | 122.04 | 256.2 | 2.1 |
| MER9a3_LTR_ERVK | ERV2 | 100.84 | 210.7 | 2.09 |
| MER4B-int_LTR_ERV1 | ERV1 | 105.81 | 221.07 | 2.09 |
| MER4-int_LTR_ERV1 | ERV1 | 317.48 | 656.31 | 2.07 |
| HERV15-int_LTR_ERV1 | ERV1 | 24.28 | 49.96 | 2.06 |
| MER57E3_LTR_ERV1 | ERV1 | 31.62 | 64.73 | 2.05 |
| LTR108a_Mam_LTR_ERVL | ERV3 | 25.92 | 52.95 | 2.04 |
| MER34A1_LTR_ERV1 | ERV1 | 77.33 | 157.29 | 2.03 |
| HERVK11D-int_LTR_ERVK | ERV2 | 6.01 | 12.2 | 2.03 |
| MLT1F1-int_LTR_ERVL-MaLR | ERV3 | 40.73 | 82.56 | 2.03 |
| LTR70_LTR_ERV1 | ERV1 | 37.51 | 75.05 | 2 |
| LTR1E_LTR_ERV1 | ERV1 | 45.06 | 21.22 | −2.12 |
| HERV-Fc1_LTR2_LTR_ERV1 | ERV1 | 0.34 | 0.16 | −2.16 |
| MER11B_LTR_ERVK | ERV2 | 25,765.36 | 3311.14 | −7.78 |
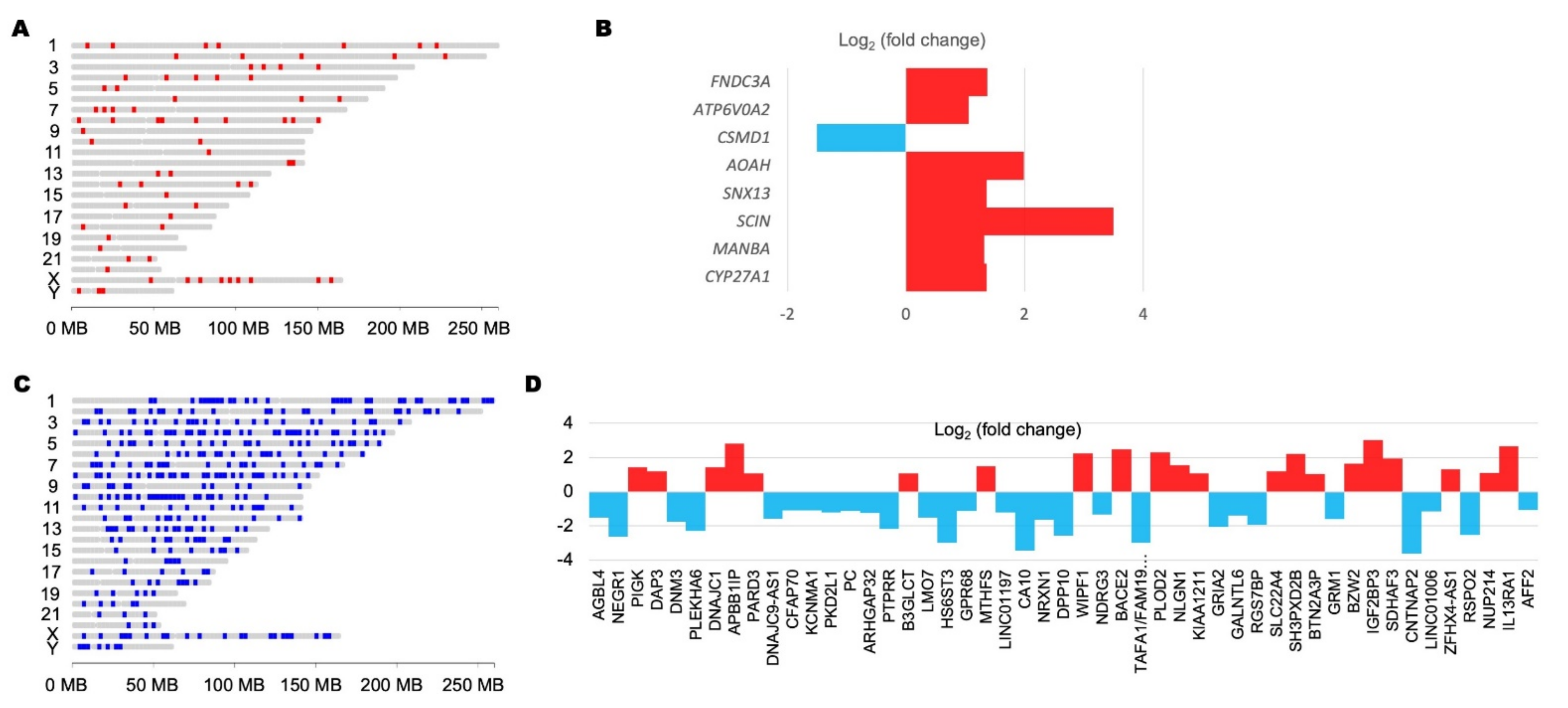
| Gene ID | Gene Name | DrugBank ID | Drug Name |
|---|---|---|---|
| SLC22A4 | solute carrier family 22 member 4 | DB00122 | Choline |
| DB00575 | Clonidine | ||
| DB01151 | Desipramine | ||
| DB00458 | Imipramine | ||
| DB01043 | Memantine | ||
| DB06691 | Mepyramine | ||
| DB00468 | Quinine | ||
| DB14754 | Solriamfetol | ||
| CA10 | carbonic anhydrase 10 | DB00909 | Zonisamide |
| GRIA2 | glutamate ionotropic receptor AMPA type subunit 2 | DB01351 | Amobarbital |
| DB00312 | Pentobarbital | ||
| DB00237 | Butabarbital | ||
| DB00241 | Butalbital | ||
| DB00306 | Talbutal | ||
| DB00418 | Secobarbital | ||
| DB00599 | Thiopental | ||
| DB00794 | Primidone | ||
| DB00849 | Methylphenobarbital | ||
| DB01174 | Phenobarbital | ||
| DB01353 | Butobarbital |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Yang, Y.; Zhang, N.; Soto, C.; Jiang, X.; An, Z.; Zheng, W.J. Human Endogenous Retroviruses in Glioblastoma Multiforme. Microorganisms 2021, 9, 764. https://doi.org/10.3390/microorganisms9040764
Yuan Z, Yang Y, Zhang N, Soto C, Jiang X, An Z, Zheng WJ. Human Endogenous Retroviruses in Glioblastoma Multiforme. Microorganisms. 2021; 9(4):764. https://doi.org/10.3390/microorganisms9040764
Chicago/Turabian StyleYuan, Zihao, Yuntao Yang, Ningyan Zhang, Claudio Soto, Xiaoqian Jiang, Zhiqiang An, and Wenjin Jim Zheng. 2021. "Human Endogenous Retroviruses in Glioblastoma Multiforme" Microorganisms 9, no. 4: 764. https://doi.org/10.3390/microorganisms9040764