
Adhesion Properties, Biofilm Forming Potential, and Susceptibility to Disinfectants of Contaminant Wine Yeasts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Samples
2.2. Yeasts Identification
2.3. Adhesion Properties of Yeasts
2.4. MAT Formation
2.5. Determination of the Minimum Inhibitory Concentrations (MICs)
2.6. Statistical Analysis
3. Results and Discussion
3.1. Yeast Identification
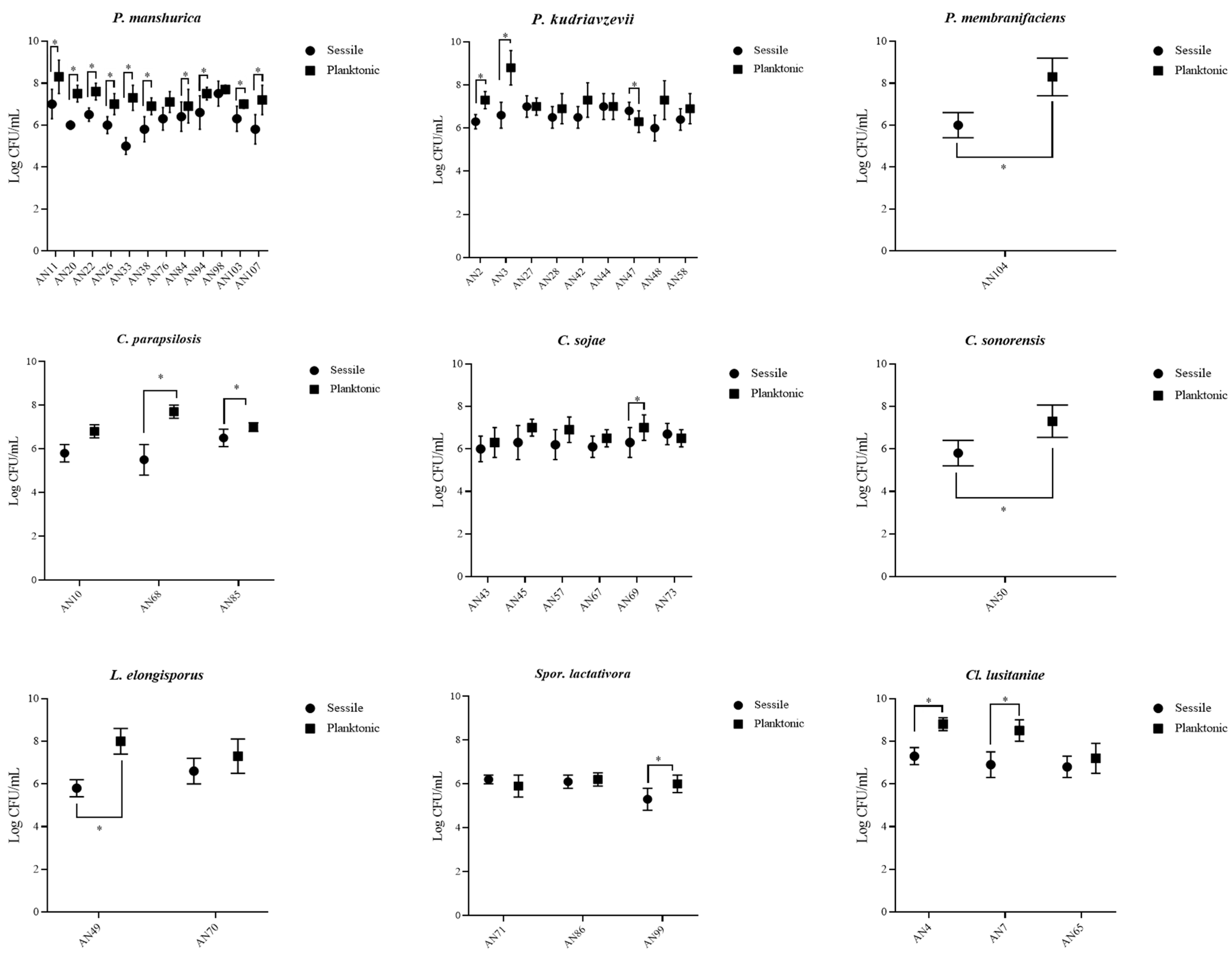
3.2. Yeast Adhesion to Abiotic Surfaces
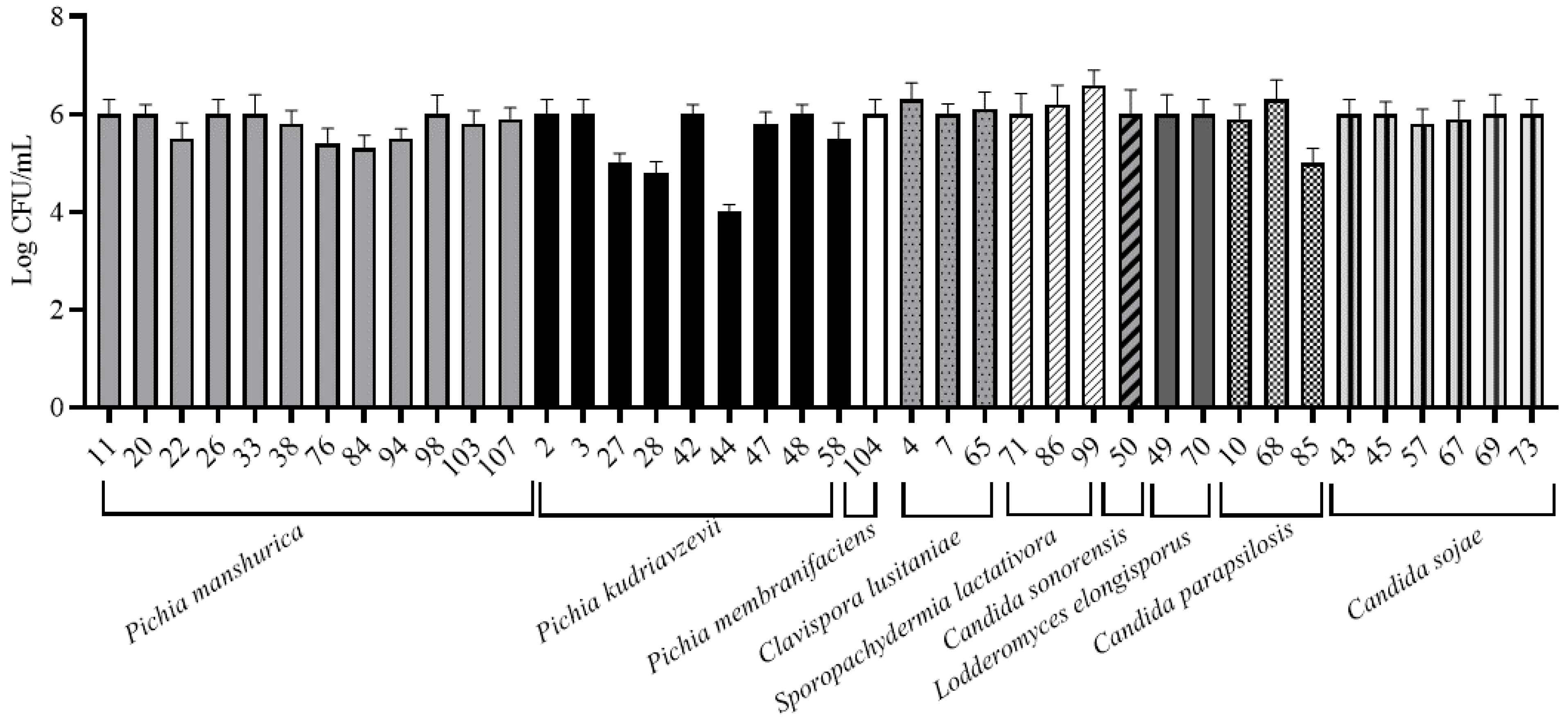
3.3. MATs Formation
3.4. Determination of the Minimum Inhibitory Concentrations (MICs)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial Terroir for Wine Grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I. Yeast and Bacterial Modulation of Wine Aroma and Flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Fleet, G.H.; Lafon-Lafourcade, S.; Ribéreau-Gayon, P. Evolution of Yeasts and Lactic Acid Bacteria during Fermentation and Storage of Bordeaux Wines. Appl. Environ. Microbiol. 1984, 48, 1034–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S. The Soil Microbiome Influences Grapevine-Associated Microbiota. MBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Morrison-Whittle, P.; Goddard, M.R. From Vineyard to Winery: A Source Map of Microbial Diversity Driving Wine Fermentation. Environ. Microbiol. 2018, 20, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A Distinct Population of Saccharomyces cerevisiae in New Zealand: Evidence for Local Dispersal by Insects and Human-Aided Global Dispersal in Oak Barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef]
- Stefanini, I.; Dapporto, L.; Legras, J.-L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S. Role of Social Wasps in Saccharomyces cerevisiae Ecology and Evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [Green Version]
- Lam, S.S.; Howell, K.S. Drosophila-Associated Yeast Species in Vineyard Ecosystems. FEMS Microbiol. Lett. 2015, 362, fnv170. [Google Scholar] [CrossRef] [Green Version]
- Knight, S.; Goddard, M.R. Quantifying Separation and Similarity in a Saccharomyces cerevisiae Metapopulation. ISME J. 2015, 9, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perpetuini, G.; Tittarelli, F.; Battistelli, N.; Suzzi, G.; Tofalo, R. Contribution of Pichia manshurica Strains to Aroma Profile of Organic Wines. Eur. Food Res. Technol. 2020, 246, 1405–1417. [Google Scholar] [CrossRef]
- Hernández, A.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Serradilla, M.J.; Villalobos, M.C.; Martín, A.; Córdoba, M.G. Spoilage Yeasts: What Are the Sources of Contamination of Foods and Beverages? Int. J. Food Microbiol. 2018, 286, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Enrique, M.; Marcos, J.F.; Yuste, M.; Martínez, M.; Vallés, S.; Manzanares, P. Antimicrobial Action of Synthetic Peptides towards Wine Spoilage Yeasts. Int. J. Food Microbiol. 2007, 118, 318–325. [Google Scholar] [CrossRef]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage Yeasts in the Wine Industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Dimopoulou, M.; Renault, M.; Dols-Lafargue, M.; Albertin, W.; Herry, J.-M.; Bellon-Fontaine, M.-N.; Masneuf-Pomarede, I. Microbiological, Biochemical, Physicochemical Surface Properties and Biofilm Forming Ability of Brettanomyces bruxellensis. Ann. Microbiol. 2019, 69, 1217–1225. [Google Scholar] [CrossRef]
- Perpetuini, G.; Tittarelli, F.; Schirone, M.; Di Gianvito, P.; Corsetti, A.; Arfelli, G.; Suzzi, G.; Tofalo, R. Adhesion Properties and Surface Hydrophobicity of Pichia manshurica Strains Isolated from Organic Wines. LWT 2018, 87, 385–392. [Google Scholar] [CrossRef]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging Technologies to Control Brettanomyces spp. in Wine: Recent Advances and Future Trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Frisón, L.N.; Chiericatti, C.A.; Aríngoli, E.E.; Basílico, J.C.; Basílico, M.Z. Effect of Different Sanitizers against Zygosaccharomyces rouxii. J. Food Sci. Technol. 2015, 52, 4619–4624. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, Y.; Yang, H. Efficacy of Low Concentration Neutralised Electrolysed Water and Ultrasound Combination for Inactivating Escherichia coli ATCC 25922, Pichia pastoris GS115 and Aureobasidium pullulans 2012 on Stainless Steel Coupons. Food Control 2017, 73, 889–899. [Google Scholar] [CrossRef]
- Reynolds, T.B.; Fink, G.R. Bakers’ Yeast, a Model for Fungal Biofilm Formation. Science 2001, 291, 878–881. [Google Scholar] [CrossRef]
- Tek, E.L.; Sundstrom, J.F.; Gardner, J.M.; Oliver, S.G.; Jiranek, V. Evaluation of the Ability of Commercial Wine Yeasts to Form Biofilms (Mats) and Adhere to Plastic: Implications for the Microbiota of the Winery Environment. FEMS Microbiol. Ecol. 2018, 94, fix188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofalo, R.; Chaves-López, C.; Di Fabio, F.; Schirone, M.; Felis, G.E.; Torriani, S.; Paparella, A.; Suzzi, G. Molecular Identification and Osmotolerant Profile of Wine Yeasts That Ferment a High Sugar Grape Must. Int. J. Food Microbiol. 2009, 130, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subunit (26S) Ribosomal DNA Partial Sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Tomičić, R.; Raspor, P. Influence of Growth Conditions on Adhesion of Yeast Candida spp. and Pichia spp. to Stainless Steel Surfaces. Food Microbiol. 2017, 65, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tristezza, M.; Lourenço, A.; Barata, A.; Brito, L.; Malfeito-Ferreira, M.; Loureiro, V. Susceptibility of Wine Spoilage Yeasts and Bacteria in the Planktonic State and in Biofilms to Disinfectants. Ann. Microbiol. 2010, 60, 549–556. [Google Scholar] [CrossRef]
- Mikata, K.; Ueda-Nishimura, K. Reclassification of Pichia membranifaciens Sensu Kurtzman. Antonie Van Leeuwenhoek 2000, 77, 159–171. [Google Scholar] [CrossRef]
- Saez, J.S.; Lopes, C.A.; Kirs, V.E.; Sangorrín, M. Production of Volatile Phenols by Pichia manshurica and Pichia membranifaciens Isolated from Spoiled Wines and Cellar Environment in Patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and Safety Aspects of Yeast Strains Characterized from Vineyards and Spontaneous Fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef]
- Del Mónaco, S.M.; Barda, N.B.; Rubio, N.C.; Caballero, A.C. Selection and Characterization of a Patagonian Pichia kudriavzevii for Wine Deacidification. J. Appl. Microbiol. 2014, 117, 451–464. [Google Scholar] [CrossRef]
- Dandi, N.D.; Dandi, B.N.; Chaudhari, A.B. Bioprospecting of Thermo- and Osmo-Tolerant Fungi from Mango Pulp–Peel Compost for Bioethanol Production. Antonie Van Leeuwenhoek 2013, 103, 723–736. [Google Scholar] [CrossRef]
- Sun, W.; Vila-Santa, A.; Liu, N.; Prozorov, T.; Xie, D.; Faria, N.T.; Ferreira, F.C.; Mira, N.P.; Shao, Z. Metabolic Engineering of an Acid-Tolerant Yeast Strain Pichia kudriavzevii for Itaconic Acid Production. Metab. Eng. Commun. 2020, 10, e00124. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Moon, S.H.; Chang, M.; Kim, H.Y.; Chang, H.C. Pichia kudriavzevii Is the Major Yeast Involved in Film-Formation, off-Odor Production, and Texture-Softening in over-Ripened Kimchi. Food Sci. Biotechnol. 2014, 23, 489–497. [Google Scholar] [CrossRef]
- Choi, D.-H.; Park, E.-H.; Kim, M.-D. Isolation of Thermotolerant Yeast Pichia kudriavzevii from Nuruk. Food Sci. Biotechnol. 2017, 26, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Zara, G.; Budroni, M.; Mannazzu, I.; Fancello, F.; Zara, S. Yeast Biofilm in Food Realms: Occurrence and Control. World J. Microbiol. Biotechnol. 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Storgårds, E.; Tapani, K.; Hartwall, P.; Saleva, R.; Suihko, M.L. Microbial Attachment and Biofilm Formation in Brewery Bottling Plants. J. Am. Soc. Brew. Chem. 2006, 64, 8–15. [Google Scholar] [CrossRef]
- Nakase, T.; Suzuki, M.; Takashima, M.; Miyakawa, Y.; Kagaya, K.; Fukazawa, Y.; Komagata, K. Candida sojae, a New Species of Yeast Isolated from an Extraction Process of Water-Soluble Substances of Defatted Soybean Flakes. J. Gen. Appl. Microbiol. 1994, 40, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast Interactions and Wine Flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- González, S.S.; Barrio, E.; Querol, A. Molecular Identification and Characterization of Wine Yeasts Isolated from Tenerife (Canary Island, Spain). J. Appl. Microbiol. 2007, 102, 1018–1025. [Google Scholar] [CrossRef]
- Ganter, P.F.; Cardinali, G.; Giammaria, M.; Quarles, B. Correlations among Measures of Phenotypic and Genetic Variation within an Oligotrophic Asexual Yeast, Candida sonorensis, Collected from Opuntia. FEMS Yeast Res. 2004, 4, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Ilmén, M.; Koivuranta, K.; Ruohonen, L.; Rajgarhia, V.; Suominen, P.; Penttilä, M. Production of L-Lactic Acid by the Yeast Candida sonorensis Expressing Heterologous Bacterial and Fungal Lactate Dehydrogenases. Microb. Cell Factories 2013, 12, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitzilaiou, E.; Stoica, I.M.; Knøchel, S. Microbial Biofilm Communities on Reverse Osmosis Membranes in Whey Water Processing before and after Cleaning. J. Membr. Sci. 2019, 587, 117174. [Google Scholar] [CrossRef]
- Pietrowski, G.A.M.; Bittencourt, J.V.M.; Brandão, L.R.; Rosa, C.A.; Alberti, A.; Nogueira, A. Identification and Selection of Non-Saccharomyces Strains Isolate from Brazilian Apple Must. Ciênc. Rural 2018, 48. [Google Scholar] [CrossRef]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of Non-Saccharomyces Yeasts to Be Used in Grape Musts with High Alcoholic Potential: A Strategy to Obtain Wines with Reduced Ethanol Content. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Pereyra, M.M.; Díaz, M.A.; Meinhardt, F.; Dib, J.R. Effect of Stress Factors Associated with Postharvest Citrus Conditions on the Viability and Biocontrol Activity of Clavispora lusitaniae Strain 146. PLoS ONE 2020, 15, e0239432. [Google Scholar] [CrossRef] [PubMed]
- Mingorance-Cazorla, L.; Clemente-Jiménez, J.M.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Contribution of Different Natural Yeasts to the Aroma of Two Alcoholic Beverages. World J. Microbiol. Biotechnol. 2003, 19, 297–304. [Google Scholar] [CrossRef]
- Coetzee, G.; Malandra, L.; Wolfaardt, G.M.; Viljoen-Bloom, M. Dynamics of a Microbial Biofilm in a Rotating Biological Contactor for the Treatment of Winery Effluent. Water SA 2004, 30, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Tomičić, R.; Tomičić, Z.; Thaler, N.; Humar, M.; Raspor, P. Factors Influencing Adhesion of Bacteria Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus and Yeast Pichia membranifaciens to Wooden Surfaces. Wood Sci. Technol. 2020, 54, 1663–1676. [Google Scholar] [CrossRef]
- Tomičić, R.; Tomičić, Z.; Raspor, P. Adhesion of Candida spp. and Pichia spp. to Wooden Surfaces. Food Technol. Biotechnol. 2017, 55, 138–142. [Google Scholar] [CrossRef]
- Alberti-Segui, C.; Morales, A.J.; Xing, H.; Kessler, M.M.; Willins, D.A.; Weinstock, K.G.; Cottarel, G.; Fechtel, K.; Rogers, B. Identification of Potential Cell-Surface Proteins in Candida albicans and Investigation of the Role of a Putative Cell-Surface Glycosidase in Adhesion and Virulence. Yeast 2004, 21, 285–302. [Google Scholar] [CrossRef]
- Malfeito-Ferreira, M. Yeasts and Wine Off-Flavours: A Technological Perspective. Ann. Microbiol. 2011, 61, 95–102. [Google Scholar] [CrossRef]
- Biniarz, P.; Baranowska, G.; Feder-Kubis, J.; Krasowska, A. The Lipopeptides Pseudofactin II and Surfactin Effectively Decrease Candida albicans Adhesion and Hydrophobicity. Antonie Van Leeuwenhoek 2015, 108, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Valotteau, C.; Prystopiuk, V.; Cormack, B.P.; Dufrêne, Y.F. Atomic Force Microscopy Demonstrates That Candida glabrata Uses Three Epa Proteins To Mediate Adhesion to Abiotic Surfaces. Msphere 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recht, J.; Martínez, A.; Torello, S.; Kolter, R. Genetic Analysis of Sliding Motility in Mycobacterium smegmatis. J. Bacteriol. 2000, 182, 4348–4351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gori, K.; Knudsen, P.B.; Nielsen, K.F.; Arneborg, N.; Jespersen, L. Alcohol-Based Quorum Sensing Plays a Role in Adhesion and Sliding Motility of the Yeast Debaryomyces hansenii. FEMS Yeast Res. 2011, 11, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perpetuini, G.; Tittarelli, F.; Suzzi, G.; Tofalo, R. Cell Wall Surface Properties of Kluyveromyces marxianus Strains from Dairy-Products. Front. Microbiol. 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofalo, R.; Perpetuini, G.; Fasoli, G.; Schirone, M.; Corsetti, A.; Suzzi, G. Biodiversity Study of Wine Yeasts Belonging to the “Terroir” of Montepulciano d’Abruzzo “Colline Teramane” Revealed Saccharomyces cerevisiae Strains Exhibiting Atypical and Unique 5.8 S-ITS Restriction Patterns. Food Microbiol. 2014, 39, 7–12. [Google Scholar] [CrossRef]
- Sarode, N.; Davis, S.E.; Tams, R.N.; Reynolds, T.B. The Wsc1p Cell Wall Signaling Protein Controls Biofilm (Mat) Formation Independently of Flo11p in Saccharomyces cerevisiae. G3 Genes Genomes Genet. 2014, 4, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Goossens, K.; Willaert, R. Flocculation Protein Structure and Cell–Cell Adhesion Mechanism in Saccharomyces cerevisiae. Biotechnol. Lett. 2010, 32, 1571–1585. [Google Scholar] [CrossRef] [Green Version]
- Lo, W.-S.; Dranginis, A.M. FLO11, a Yeast Gene Related to the STA Genes, Encodes a Novel Cell Surface Flocculin. J. Bacteriol. 1996, 178, 7144–7151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willaert, R.G. Adhesins of Yeasts: Protein Structure and Interactions. J. Fungi 2018, 4, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengeler, K.B.; Davidson, R.C.; D’souza, C.; Harashima, T.; Shen, W.-C.; Wang, P.; Pan, X.; Waugh, M.; Heitman, J. Signal Transduction Cascades Regulating Fungal Development and Virulence. Microbiol. Mol. Biol. Rev. 2000, 64, 746–785. [Google Scholar] [CrossRef] [Green Version]
- Liu, H. Transcriptional Control of Dimorphism in Candida albicans. Curr. Opin. Microbiol. 2001, 4, 728–735. [Google Scholar] [CrossRef]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A Yeast by Any Other Name: Candida glabrata and Its Interaction with the Host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef]
- Maidan, M.M.; De Rop, L.; Serneels, J.; Exler, S.; Rupp, S.; Tournu, H.; Thevelein, J.M.; Van Dijck, P. The G Protein-Coupled Receptor Gpr1 and the Gα Protein Gpa2 Act through the CAMP-Protein Kinase A Pathway to Induce Morphogenesis in Candida albicans. Mol. Biol. Cell 2005, 16, 1971–1986. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Fox, E.P.; Nett, J.E.; Sorrells, T.R.; Mitrovich, Q.M.; Hernday, A.D.; Tuch, B.B.; Andes, D.R.; Johnson, A.D. A Recently Evolved Transcriptional Network Controls Biofilm Development in Candida albicans. Cell 2012, 148, 126–138. [Google Scholar] [CrossRef] [Green Version]
- Cormack, B.P.; Ghori, N.; Falkow, S. An Adhesin of the Yeast Pathogen Candida glabrata Mediating Adherence to Human Epithelial Cells. Science 1999, 285, 578–582. [Google Scholar] [CrossRef]
- Sarode, N.; Miracle, B.; Peng, X.; Ryan, O.; Reynolds, T.B. Vacuolar Protein Sorting Genes Regulate Mat Formation in Saccharomyces cerevisiae by Flo11p-Dependent and-Independent Mechanisms. Eukaryot. Cell 2011, 10, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Mills, D.A. Facility-Specific “House” Microbiome Drives Microbial Landscapes of Artisan Cheesemaking Plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef] [Green Version]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Deniel, F.; Coton, E.; Mounier, J. Diversity of Spoilage Fungi Associated with Various French Dairy Products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Resa, C.P.O.; Jagus, R.J.; Gerschenson, L.N. Natamycin Efficiency for Controlling Yeast Growth in Models Systems and on Cheese Surfaces. Food Control 2014, 35, 101–108. [Google Scholar] [CrossRef]
- Romero-Gil, V.; García-García, P.; Garrido-Fernández, A.; Arroyo-López, F.N. Susceptibility and Resistance of Lactic Acid Bacteria and Yeasts against Preservatives with Potential Application in Table Olives. Food Microbiol. 2016, 54, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Winniczuk, P.P.; Parish, M.E. Minimum Inhibitory Concentrations of Antimicrobials against Micro-Organisms Related to Citrus Juice. Food Microbiol. 1997, 14, 373–381. [Google Scholar] [CrossRef]
- Divol, B.; du Toit, M.; Duckitt, E. Surviving in the Presence of Sulphur Dioxide: Strategies Developed by Wine Yeasts. Appl. Microbiol. Biotechnol. 2012, 95, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Erten, H. Relations between Elevated Temperatures and Fermentation Behaviour of Kloeckera apiculata and Saccharomyces cerevisiae Associated with Winemaking in Mixed Cultures. World J. Microbiol. Biotechnol. 2002, 18, 377–382. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strains | Species | PA (0.05–10%) | SH (0.5–5%) | Potassium Metabisulphite (5–1600 ppm) |
|---|---|---|---|---|
| AN11 | P. manshurica | 0.2 | 1 | 140 |
| AN20, AN22, AN38 | 0.2 | 1.2 | 160 | |
| AN26 | 0.2 | 1.2 | 140 | |
| AN33, AN76 | 0.4 | 1.2 | 160 | |
| AN84, AN103 | 0.2 | 1.2 | 150 | |
| AN94 | 0.2 | 1 | 160 | |
| AN98, AN107 | 0.2 | 1.2 | 160 | |
| PED 141-1 | 0.25 | 5 | 150 | |
| AN2, AN3 | P. kudriavzevii | 1 | 2 | 160 |
| AN27, AN28 | 0.8 | 1.2 | 120 | |
| AN42 | 0.6 | 1.2 | 140 | |
| AN44 | 0.6 | 1.2 | 80 | |
| AN47, AN48, AN58 | 1 | 2 | 160 | |
| AN4 | Cl. lusitaniae | 1 | 2 | 120 |
| AN7 | 0.5 | 1.2 | 100 | |
| AN65 | 1 | 2 | 130 | |
| AN71 | Spor. lactativora | 0.2 | 0.8 | 120 |
| AN86 | 0.5 | 0.4 | 80 | |
| AN99 | 0.1 | 0.6 | 100 | |
| AN10, AN68, AN85 | C. parapsilosis | 0.08 | 1 | 120 |
| AN43 | 0.1 | 2 | 130 | |
| AN45 | 0.1 | 2 | 120 | |
| AN57 | C. sojae | 0.2 | 2 | 110 |
| AN67, AN73 | 0.8 | 2 | 80 | |
| AN69 | 1 | 2 | 120 | |
| AN104 | P. membranifaciens | 0.6 | 1.5 | 140 |
| AN50 | C. sonorensis | 1 | 2 | 130 |
| AN49, AN70 | L. elongisporus | 0.6 | 1.5 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perpetuini, G.; Rossetti, A.P.; Battistelli, N.; Arfelli, G.; Tofalo, R. Adhesion Properties, Biofilm Forming Potential, and Susceptibility to Disinfectants of Contaminant Wine Yeasts. Microorganisms 2021, 9, 654. https://doi.org/10.3390/microorganisms9030654
Perpetuini G, Rossetti AP, Battistelli N, Arfelli G, Tofalo R. Adhesion Properties, Biofilm Forming Potential, and Susceptibility to Disinfectants of Contaminant Wine Yeasts. Microorganisms. 2021; 9(3):654. https://doi.org/10.3390/microorganisms9030654
Chicago/Turabian StylePerpetuini, Giorgia, Alessio Pio Rossetti, Noemi Battistelli, Giuseppe Arfelli, and Rosanna Tofalo. 2021. "Adhesion Properties, Biofilm Forming Potential, and Susceptibility to Disinfectants of Contaminant Wine Yeasts" Microorganisms 9, no. 3: 654. https://doi.org/10.3390/microorganisms9030654