Effect of Co-Inoculation of Candida zemplinina, Saccharomyces cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy)

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains
2.2. Microbial Population Analyses
2.3. Lab-Scale Tests
2.4. Pilot-Scale Vinification
2.5. Industrial-Scale Vinification
2.6. Wines Chemical Analyses
2.7. Statistical Analysis
3. Results and Discussion
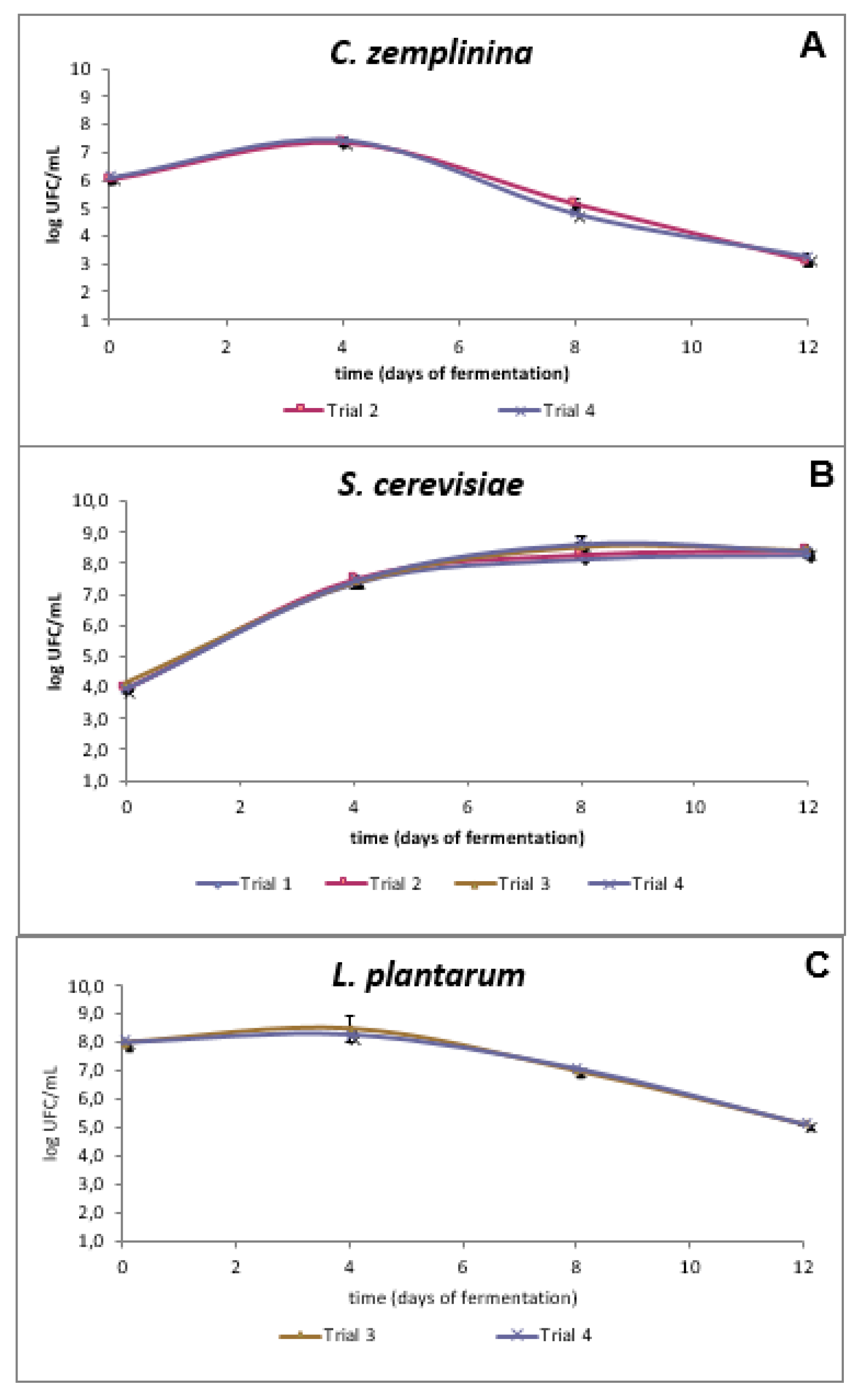
3.1. Lab-Scale Trials
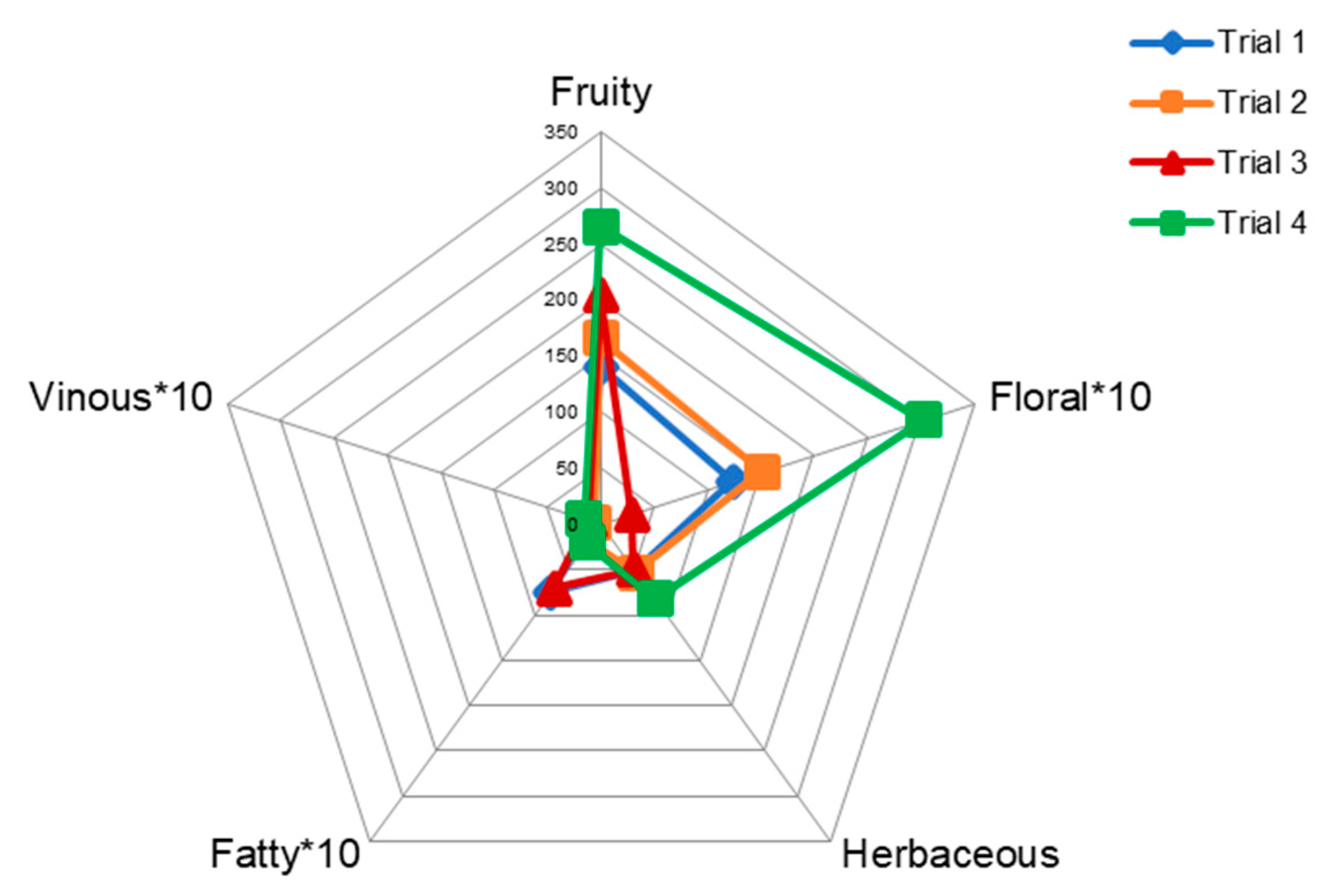
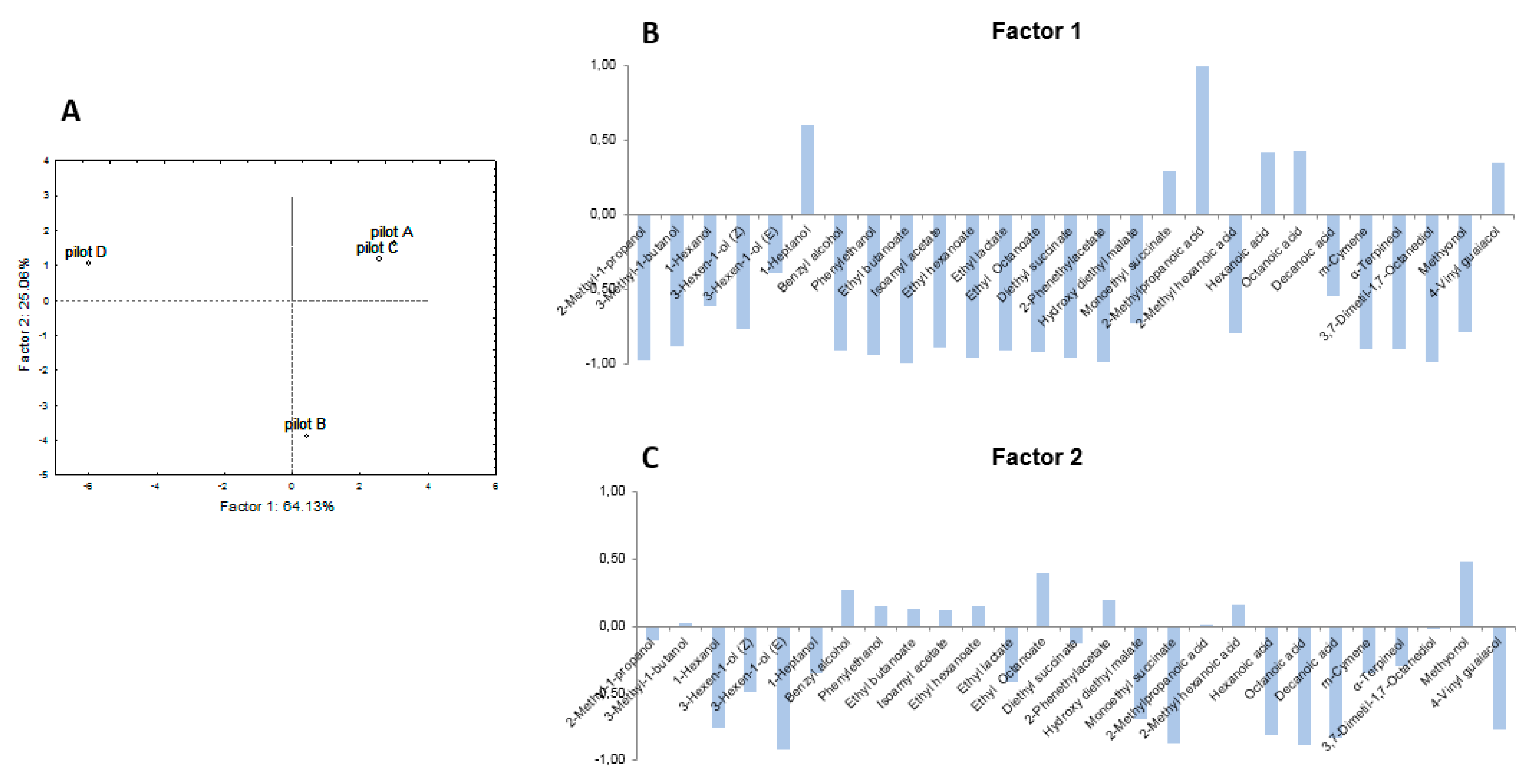
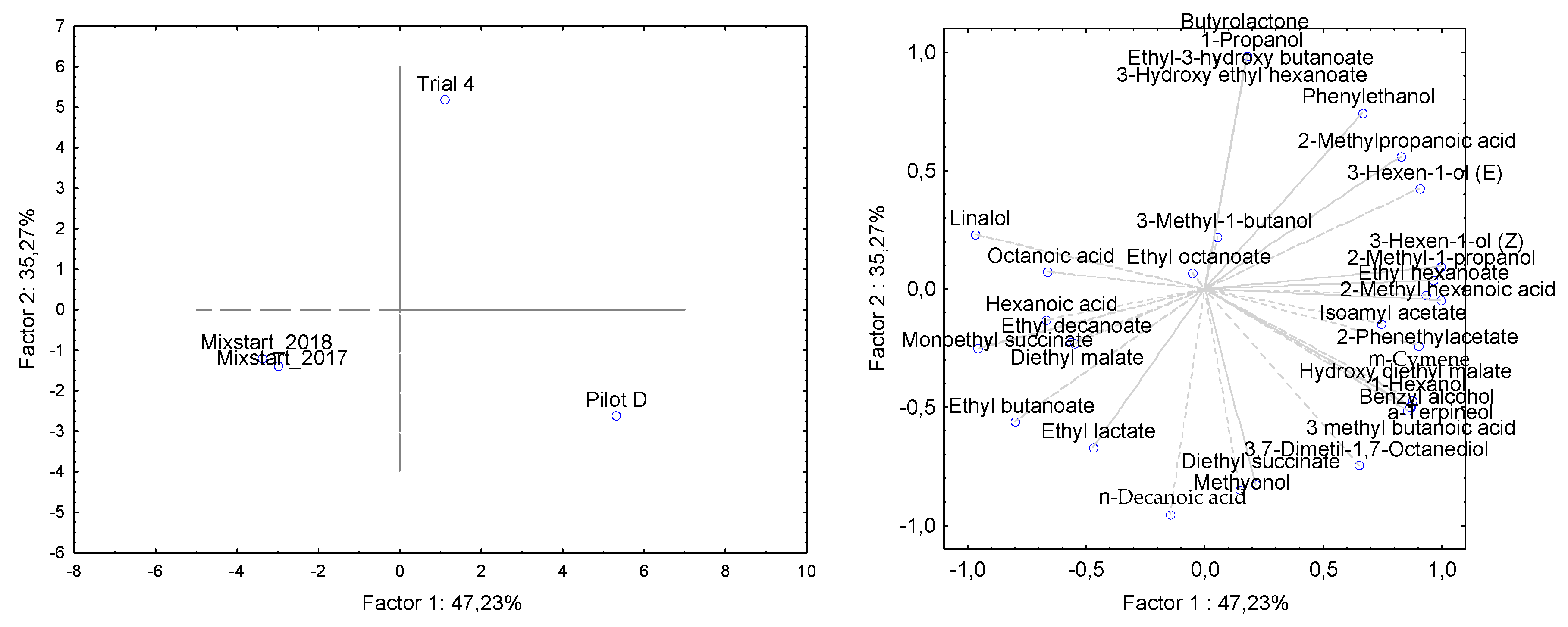
3.2. Pilot-Scale Trials
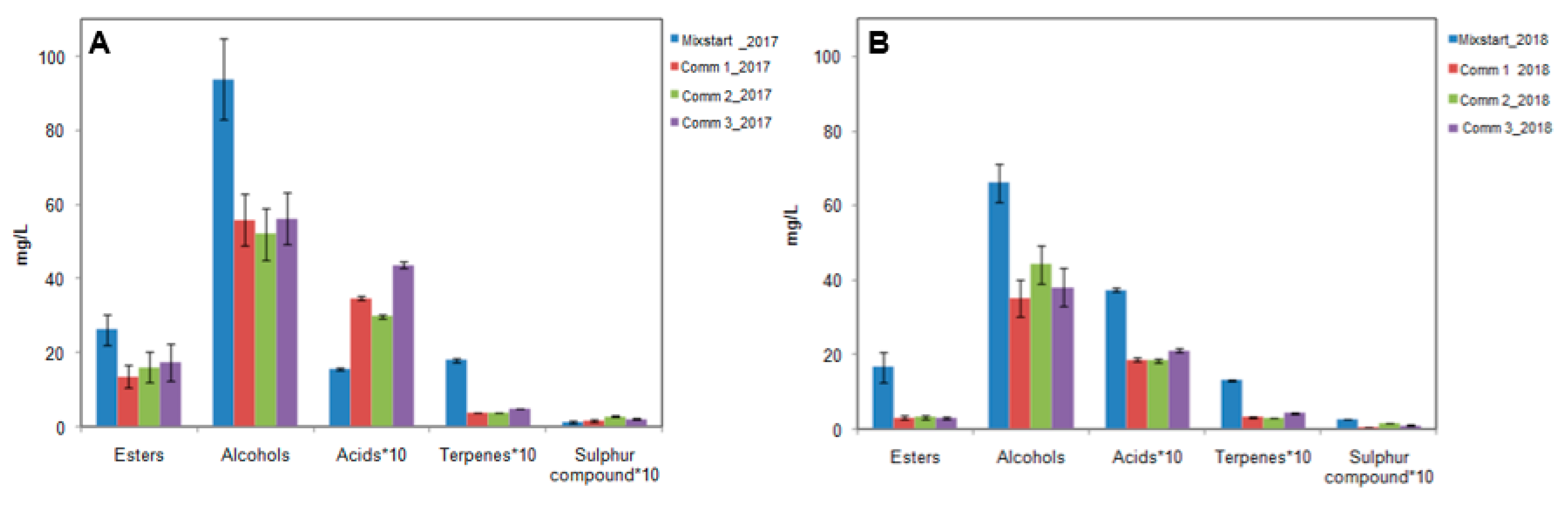
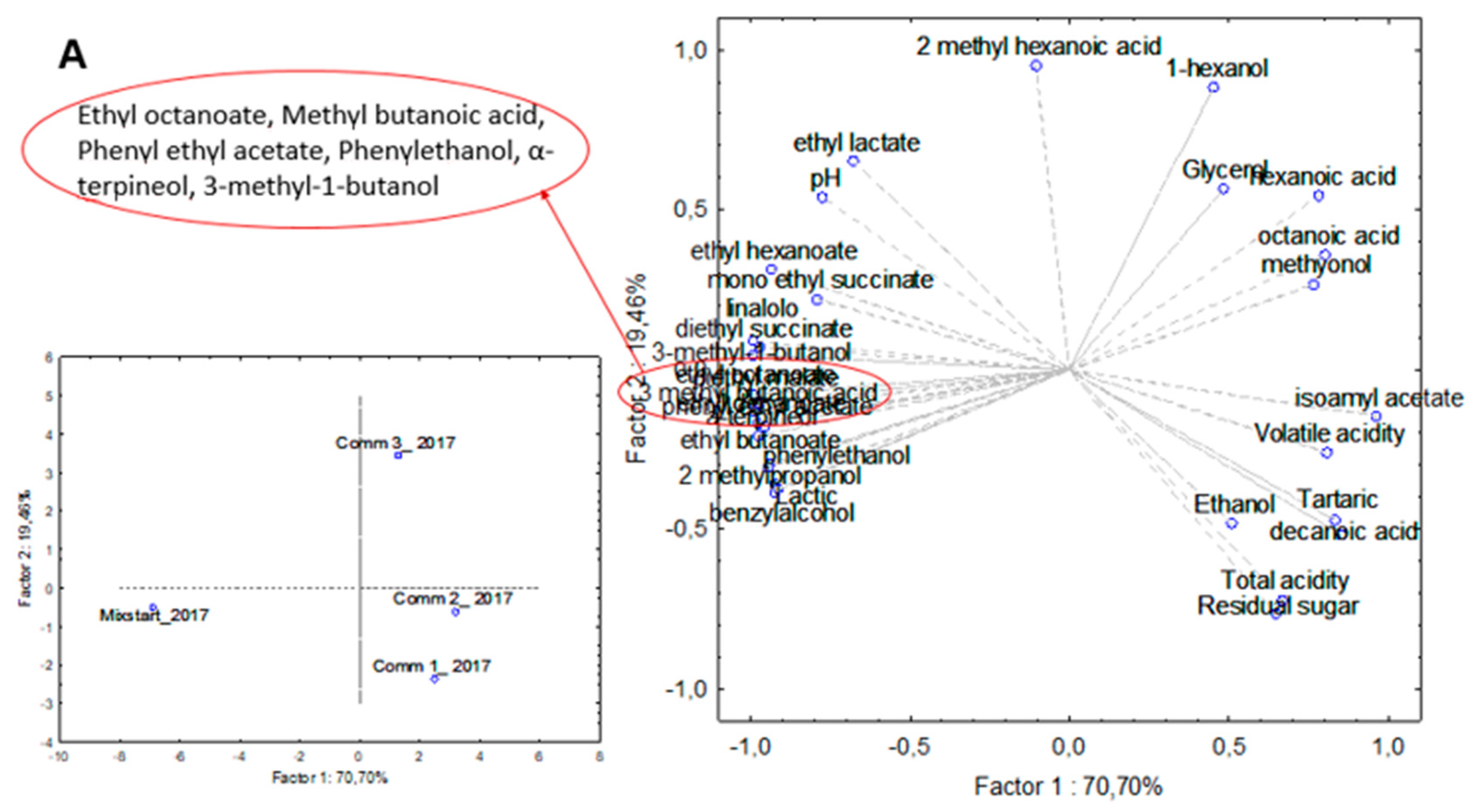
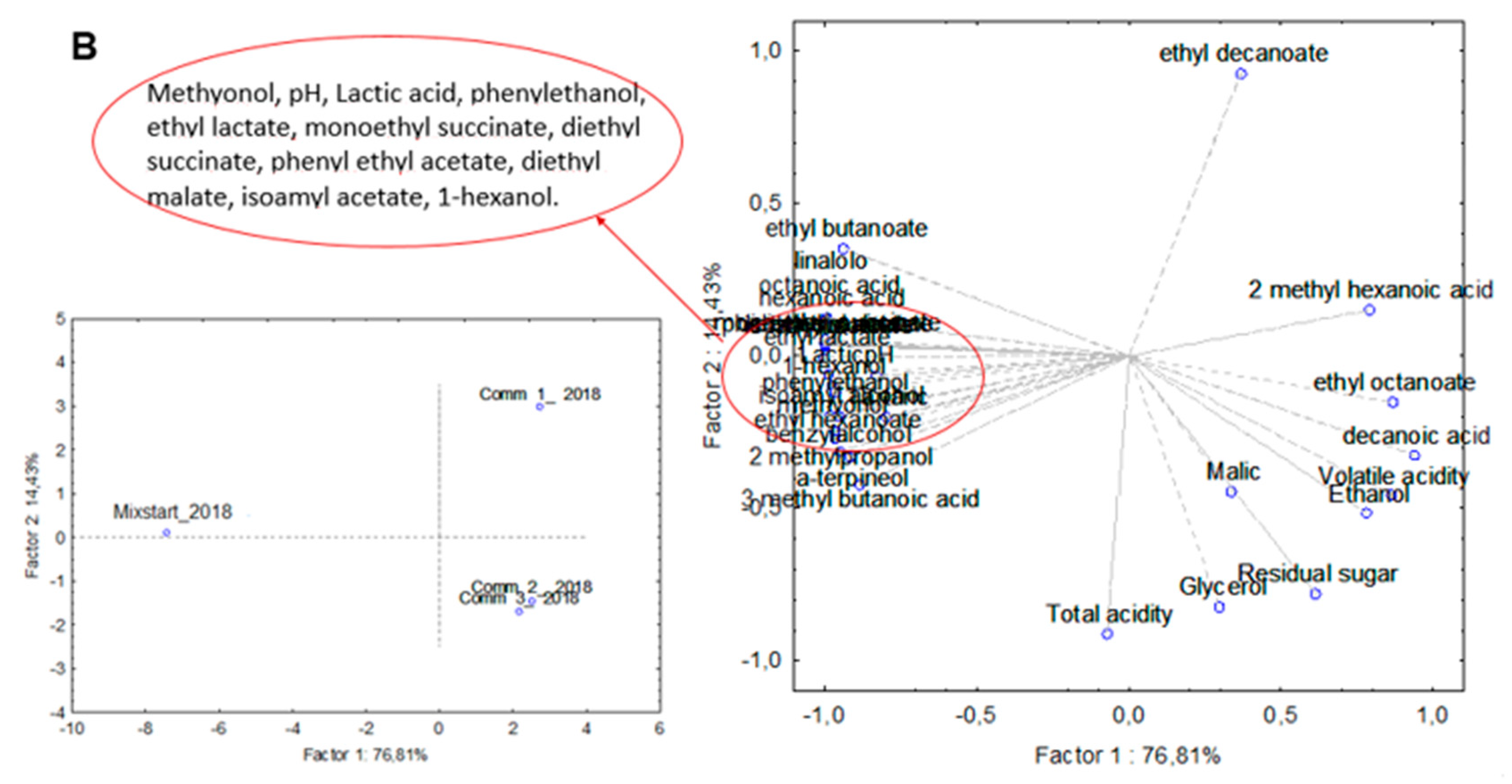
3.3. Industrial-Scale Vinifications
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Grangeteau, C.; Roullier-Gall, C.; Rousseaux, S.; Gougeon, R.D.; Schmitt-Kopplin, P.; Alexandre, H.; Guilloux-Benatier, M. Wine microbiology is driven by vineyard and winery anthropogenic factors. Microb. Biotechnol. 2017, 10, 354–370. [Google Scholar] [CrossRef] [PubMed]
- Tempère, S.; Marchal, A.; Barbe, J.-C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Sills, H.; Khoury, M.E.; Gammacurta, M.; Miot-Sertier, C.; Dutilh, L.; Vestner, J.; Capozzi, V.; Sherman, D.; Hubert, C.; Claisse, O.; et al. Two different Oenococcus oeni lineages are associated to either red or white wines in Burgundy: Genomics and metabolomics insights. OENO One 2017, 51, 309. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; du Toit, M. Selection and Characterisation of Oenococcus oeni and Lactobacillus plantarum South African Wine Isolates for Use as Malolactic Fermentation Starter Cultures. S. Afr. J. Enol. Vitic. 2011, 32, 280–295. [Google Scholar] [CrossRef] [Green Version]
- Lucio, O.; Pardo, I.; Heras, J.M.; Krieger-Weber, S.; Ferrer, S. Use of starter cultures of Lactobacillus to induce malolactic fermentation in wine. Aust. J. Grape Wine Res. 2017, 23, 15–21. [Google Scholar] [CrossRef]
- Capozzi, V.; Russo, P.; Ladero, V.; Fernández, M.; Fiocco, D.; Alvarez, M.A.; Grieco, F.; Spano, G. Biogenic Amines Degradation by Lactobacillus plantarum: Toward a Potential Application in Wine. Front. Microbiol. 2012, 3, 122. [Google Scholar] [CrossRef] [Green Version]
- Bartowsky, E.J.; Costello, P.J.; Chambers, P.J. Emerging trends in the application of malolactic fermentation. Aust. J. Grape Wine Res. 2015, 21, 663–669. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; La Hens, D.V.; Hollmann, A.; Delfederico, L.; Caballero, A.; Tymczyszyn, E.E.; Semorile, L. Comparative vinification assays with selected Patagonian strains of Oenococcus oeni and Lactobacillus plantarum. LWT 2017, 77, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Russo, P.; Capozzi, V.; Spano, G.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Metabolites of Microbial Origin with an Impact on Health: Ochratoxin A and Biogenic Amines. Front. Microbiol. 2016, 7, 482. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Tristezza, M.; Grieco, F.; Capozzi, V. Microbial Resources and Innovation in the Wine Production Sector. S. Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- De Benedictis, M.; Bleve, G.; Grieco, F.; Tristezza, M.; Tufariello, M.; Grieco, F. An optimized procedure for the enological selection of non-Saccharomyces starter cultures. Antonie Van Leeuwenhoek 2011, 99, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef] [PubMed]
- Masneuf-Pomarede, I.; Juquin, E.; Miot-Sertier, C.; Renault, P.; Laizet, Y.; Salin, F.; Alexandre, H.; Capozzi, V.; Cocolin, L.; Colonna-Ceccaldi, B.; et al. The yeast Starmerella bacillaris (synonym Candida zemplinina) shows high genetic diversity in winemaking environments. FEMS Yeast Res. 2015, 15, fov045. [Google Scholar] [CrossRef] [Green Version]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2019. [Google Scholar] [CrossRef]
- Magyar, I.; Tóth, T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 94–100. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef]
- Rantsiou, K.; Dolci, P.; Giacosa, S.; Torchio, F.; Tofalo, R.; Torriani, S.; Suzzi, G.; Rolle, L.; Cocolin, L. Candida zemplinina can reduce acetic acid produced by Saccharomyces cerevisiae in sweet wine fermentations. Appl. Environ. Microbiol. 2012, 78, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma Profile of Montepulciano d’Abruzzo Wine Fermented by Single and Co-culture Starters of Autochthonous Saccharomyces and Non-saccharomyces Yeasts. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Zara, G.; Mannazzu, I.; Caro, A.D.; Budroni, M.; Pinna, M.B.; Murru, M.; Farris, G.A.; Zara, S. Wine quality improvement through the combined utilisation of yeast hulls and Candida zemplinina/Saccharomyces cerevisiae mixed starter cultures. Aust. J. Grape Wine Res. 2014, 20, 199–207. [Google Scholar] [CrossRef]
- Englezos, V.; Torchio, F.; Cravero, F.; Marengo, F.; Giacosa, S.; Gerbi, V.; Rantsiou, K.; Rolle, L.; Cocolin, L. Aroma profile and composition of Barbera wines obtained by mixed fermentations of Starmerella bacillaris (synonym Candida zemplinina) and Saccharomyces cerevisiae. LWT 2016, 73, 567–575. [Google Scholar] [CrossRef]
- Russo, P.; Tufariello, M.; Renna, R.; Tristezza, M.; Taurino, M.; Palombi, L.; Capozzi, V.; Rizzello, C.G.; Grieco, F. New Insights into the Oenological Significance of Candida zemplinina: Impact of Selected Autochthonous Strains on the Volatile Profile of Apulian Wines. Microorganisms 2020, 8, 628. [Google Scholar] [CrossRef] [PubMed]
- Abrahamse, C.E.; Bartowsky, E.J. Timing of malolactic fermentation inoculation in Shiraz grape must and wine: Influence on chemical composition. World J. Microbiol. Biotechnol. 2012, 28, 255–265. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The Oenological Potential of Hanseniaspora uvarum in Simultaneous and Sequential Co-fermentation with Saccharomyces cerevisiae for Industrial Wine Production. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; El Khoury, M.; Lucas, P.; Bely, M.; Russo, P.; Spano, G.; Capozzi, V. Autochthonous starter cultures and indigenous grape variety for regional wine production. J. Appl. Microbiol. 2015, 118, 1395–1408. [Google Scholar] [CrossRef]
- Berbegal, C.; Borruso, L.; Fragasso, M.; Tufariello, M.; Russo, P.; Brusetti, L.; Spano, G.; Capozzi, V. A Metagenomic-Based Approach for the Characterization of Bacterial Diversity Associated with Spontaneous Malolactic Fermentations in Wine. Int. J. Mol. Sci. 2019, 20, 3980. [Google Scholar] [CrossRef] [Green Version]
- Nardi, T.; Panero, L.; Petrozziello, M.; Guaita, M.; Tsolakis, C.; Cassino, C.; Vagnoli, P.; Bosso, A. Managing wine quality using Torulaspora delbrueckii and Oenococcus oeni starters in mixed fermentations of a red Barbera wine. Eur. Food Res. Technol. 2019, 245, 293–307. [Google Scholar] [CrossRef]
- Ivit, N.N.; Kemp, B. The Impact of non-Saccharomyces Yeast on Traditional Method Sparkling Wine. Fermentation 2018, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Minnaar, P.P.; du Plessis, H.W.; Jolly, N.P.; van der Rijst, M.; du Toit, M. Non-Saccharomyces yeast and lactic acid bacteria in Co-inoculated fermentations with two Saccharomyces cerevisiae yeast strains: A strategy to improve the phenolic content of Syrah wine. Food Chem. X 2019, 4, 100070. [Google Scholar] [CrossRef]
- Lorentzen, M.P.G.; Lucas, P.M. Distribution of Oenococcus oeni populations in natural habitats. Appl. Microbiol. Biotechnol. 2019, 103, 2937–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tufariello, M.; Chiriatti, M.A.; Grieco, F.; Perrotta, C.; Capone, S.; Rampino, P.; Tristezza, M.; Mita, G.; Grieco, F. Influence of autochthonous Saccharomyces cerevisiae strains on volatile profile of Negroamaro wines. LWT Food Sci. Technol. 2014, 58, 35–48. [Google Scholar] [CrossRef]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological properties of Lactobacillus plantarum strains isolated from grape must fermentation. Food Microbiol. 2016, 57, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Bleve, G.; Lezzi, C.; Chiriatti, M.A.; D’Ostuni, I.; Tristezza, M.; Di Venere, D.; Sergio, L.; Mita, G.; Grieco, F. Selection of non-conventional yeasts and their use in immobilized form for the bioremediation of olive oil mill wastewaters. Bioresour. Technol. 2011, 102, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Gerardi, C.; Logrieco, A.; Grieco, F. An optimized protocol for the production of interdelta markers in Saccharomyces cerevisiae by using capillary electrophoresis. J. Microbiol. Methods 2009, 78, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Grieco, F.; Tufariello, M.; Quarta, A.; Mita, G.; Spano, G.; Grieco, F. Autochthonous fermentation starters for the industrial production of Negroamaro wines. J. Ind. Microbiol. Biotechnol. 2012, 39, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Grieco, F.; Tristezza, M.; Vetrano, C.; Bleve, G.; Panico, E.; Grieco, F.; Mita, G.; Logrieco, A. Exploitation of autochthonous micro-organism potential to enhance the quality of Apulian wines. Ann. Microbiol. 2011, 61, 67–73. [Google Scholar] [CrossRef]
- Fragasso, M.; Antonacci, D.; Pati, S.; Tufariello, M.; Baiano, A.; Forleo, L.R.; Caputo, A.R.; Notte, E.L. Influence of Training System on Volatile and Sensory Profiles of Primitivo Grapes and Wines. Am. J. Enol. Vitic. 2012, 63, 477–486. [Google Scholar] [CrossRef]
- Balmaseda, A.; Bordons, A.; Reguant, C.; Bautista-Gallego, J. Non-Saccharomyces in Wine: Effect Upon Oenococcus oeni and Malolactic Fermentation. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van der Rijst, M.; Kidd, M.; Jolly, N. Effect of Saccharomyces, Non-Saccharomyces Yeasts and Malolactic Fermentation Strategies on Fermentation Kinetics and Flavor of Shiraz Wines. Fermentation 2017, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Cachón, D.C.; Rantsiou, K.; Blanco, P.; Petrozziello, M.; Pollon, M.; Giacosa, S.; Río Segade, S.; Rolle, L.; Cocolin, L. Effect of mixed species alcoholic fermentation on growth and malolactic activity of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2019, 103, 7687–7702. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Englezos, V.; Capozzi, V.; Pollon, M.; Río Segade, S.; Rantsiou, K.; Spano, G.; Cocolin, L. Effect of mixed fermentations with Starmerella bacillaris and Saccharomyces cerevisiae on management of malolactic fermentation. Food Res. Int. 2020, 134, 109246. [Google Scholar] [CrossRef]
- Tufariello, M.; Capone, S.; Siciliano, P. Volatile components of Negroamaro red wines produced in Apulian Salento area. Food Chem. 2012, 132, 2155–2164. [Google Scholar] [CrossRef]
- Bordet, F.; Joran, A.; Klein, G.; Roullier-Gall, C.; Alexandre, H. Yeast–Yeast Interactions: Mechanisms, Methodologies and Impact on Composition. Microorganisms 2020, 8, 600. [Google Scholar] [CrossRef] [Green Version]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of Pure and Mixed Cultures of the Main Wine Yeast Species on Grape Must Fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Pielech-Przybylska, K.; Balcerek, M.; Dziekońska-Kubczak, U.; Pacholczyk-Sienicka, B.; Ciepielowski, G.; Albrecht, Ł.; Patelski, P. The Role of Saccharomyces cerevisiae Yeast and Lactic Acid Bacteria in the Formation of 2-Propanol from Acetone during Fermentation of Rye Mashes Obtained Using Thermal-Pressure Method of Starch Liberation. Molecules 2019, 24, 610. [Google Scholar] [CrossRef] [Green Version]
- Kłosowski, G.; Mikulski, D.; Macko, D.; Miklaszewska, B.; Kotarska, K.; Czupryński, B. Influence of various yeast strains and selected starchy raw materials on production of higher alcohols during the alcoholic fermentation process. Eur. Food Res. Technol. 2015, 240, 233–242. [Google Scholar] [CrossRef]
- Antalick, G.; Perello, M.C.; Revel, G. de Co-inoculation with Yeast and LAB Under Winery Conditions: Modification of the Aromatic Profile of Merlot Wines. South Afr. J. Enol. Vitic. 2013, 34, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Antalick, G.; Perello, M.-C.; de Revel, G. Characterization of fruity aroma modifications in red wines during malolactic fermentation. J. Agric. Food Chem. 2012, 60, 12371–12383. [Google Scholar] [CrossRef]
- Izquierdo Cañas, P.M.; Pérez-Martín, F.; García Romero, E.; Seseña Prieto, S.; Palop Herreros, M. de los L. Influence of inoculation time of an autochthonous selected malolactic bacterium on volatile and sensory profile of Tempranillo and Merlot wines. Int. J. Food Microbiol. 2012, 156, 245–254. [Google Scholar] [CrossRef]
- Ugliano, M.; Moio, L. Changes in the Concentration of Yeast-Derived Volatile Compounds of Red Wine during Malolactic Fermentation with Four Commercial Starter Cultures of Oenococcus oeni. J. Agric. Food Chem. 2005, 53, 10134–10139. [Google Scholar] [CrossRef] [PubMed]
- Maicas, S.; Gil, J.-V.; Pardo, I.; Ferrer, S. Improvement of volatile composition of wines by controlled addition of malolactic bacteria. Food Res. Int. 1999, 32, 491–496. [Google Scholar] [CrossRef]
- Knoll, C.; Fritsch, S.; Schnell, S.; Grossmann, M.; Rauhut, D.; du Toit, M. Influence of pH and ethanol on malolactic fermentation and volatile aroma compound composition in white wines. LWT Food Sci. Technol. 2011, 44, 2077–2086. [Google Scholar] [CrossRef]
- Perestrelo, R.; Fernandes, A.; Albuquerque, F.F.; Marques, J.C.; Câmara, J.S. Analytical characterization of the aroma of Tinta Negra Mole red wine: Identification of the main odorants compounds. Anal. Chim. Acta 2006, 563, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef]
- García-Carpintero, E.G.; Sánchez-Palomo, E.; Gallego, M.A.G.; González-Viñas, M.A. Volatile and sensory characterization of red wines from cv. Moravia Agria minority grape variety cultivated in La Mancha region over five consecutive vintages. Food Res. Int. 2011, 44, 1549–1560. [Google Scholar] [CrossRef]
- Francis, I.L.; Newton, J.L. Determining wine aroma from compositional data. Aust. J. Grape Wine Res. 2005, 11, 114–126. [Google Scholar] [CrossRef]
- Franco, M.; Peinado, R.A.; Medina, M.; Moreno, J. Off-Vine Grape Drying Effect on Volatile Compounds and Aromatic Series in Must from Pedro Ximénez Grape Variety. J. Agric. Food Chem. 2004, 52, 3905–3910. [Google Scholar] [CrossRef]
- Capone, S.; Tufariello, M.; Siciliano, P. Analytical characterisation of Negroamaro red wines by “Aroma Wheels”. Food Chem. 2013, 141, 2906–2915. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.-C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, C.; Spano, G.; Fragasso, M.; Grieco, F.; Russo, P.; Capozzi, V. Starter cultures as biocontrol strategy to prevent Brettanomyces bruxellensis proliferation in wine. Appl. Microbiol. Biotechnol. 2018, 102, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate Changes and Food Quality: The Potential of Microbial Activities as Mitigating Strategies in the Wine Sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Berbegal, C.; Garofalo, C.; Russo, P.; Pati, S.; Capozzi, V.; Spano, G. Use of Autochthonous Yeasts and Bacteria in Order to Control Brettanomyces bruxellensis in Wine. Fermentation 2017, 3, 65. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef]
- Tofalo, R.; Chaves-López, C.; Di Fabio, F.; Schirone, M.; Felis, G.E.; Torriani, S.; Paparella, A.; Suzzi, G. Molecular identification and osmotolerant profile of wine yeasts that ferment a high sugar grape must. Int. J. Food Microbiol. 2009, 130, 179–187. [Google Scholar] [CrossRef]
- Rankine, B.C.; Bridson, D.A. Glycerol in Australian Wines and Factors Influencing Its Formation. Am. J. Enol. Vitic. 1971, 22, 6–12. [Google Scholar]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. South Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking wine lactic acid bacteria diversity with wine aroma and flavour. Int. J. Food Microbiol. 2017, 243, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Russo, P. Microbiological Safety and the Management of Microbial Resources in Artisanal Foods and Beverages: The Need for a Transdisciplinary Assessment to Conciliate Actual Trends and Risks Avoidance. Microorganisms 2020, 8, 306. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Spano, G. Food microbial biodiversity and “microbes of protected origin”. Front. Microbiol. 2011, 2, 237. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous Food Fermentations and Potential Risks for Human Health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Nardi, T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [Green Version]
- Rossouw, D.; Jolly, N.; Jacobson, D.; Bauer, F.F. The effect of scale on gene expression: Commercial versus laboratory wine fermentations. Appl. Microbiol. Biotechnol. 2012, 93, 1207–1219. [Google Scholar] [CrossRef]
- Sampaio, T.L.; Kennedy, J.A.; Vasconcelos, M.C. Use of Microscale Fermentations in Grape and Wine Research. Am. J. Enol. Vitic. 2007, 58, 534–539. [Google Scholar]
- Lopes, C.A.; Van Broock, M.; Querol, A.; Caballero, A.C. Saccharomyces cerevisiae wine yeast populations in a cold region in Argentinean Patagonia. A study at different fermentation scales. J. Appl. Microbiol. 2002, 93, 608–615. [Google Scholar] [CrossRef]
- Schmid, F.; Schadt, J.; Jiranek, V.; Block, D.E. Formation of temperature gradients in large- and small-scale red wine fermentations during cap management. Aust. J. Grape Wine Res. 2009, 15, 249–255. [Google Scholar] [CrossRef]
- Capozzi, V.; Russo, P.; Beneduce, L.; Weidmann, S.; Grieco, F.; Guzzo, J.; Spano, G. Technological properties of Oenococcus oeni strains isolated from typical southern Italian wines. Lett. Appl. Microbiol. 2010, 50, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Berbegal, C.; Grieco, F.; Tufariello, M.; Spano, G.; Capozzi, V. Selection of indigenous yeast strains for the production of sparkling wines from native Apulian grape varieties. Int. J. Food Microbiol. 2018, 285, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Arena, M.P.; Laddomada, B.; Cappello, M.S.; Bleve, G.; Grieco, F.; Beneduce, L.; Berbegal, C.; Spano, G.; Capozzi, V. Starter Cultures for Sparkling Wine. Fermentation 2016, 2, 21. [Google Scholar] [CrossRef]
- Garofalo, C.; Tristezza, M.; Grieco, F.; Spano, G.; Capozzi, V. From grape berries to wine: Population dynamics of cultivable yeasts associated to “Nero di Troia” autochthonous grape cultivar. World J. Microbiol. Biotechnol. 2016, 32, 59. [Google Scholar] [CrossRef]
- Capozzi, V.; Berbegal, C.; Tufariello, M.; Grieco, F.; Spano, G.; Grieco, F. Impact of co-inoculation of Saccharomyces cerevisiae, Hanseniaspora uvarum and Oenococcus oeni autochthonous strains in controlled multi starter grape must fermentations. LWT 2019, 109, 241–249. [Google Scholar] [CrossRef]
- Di Toro, M.R.; Capozzi, V.; Beneduce, L.; Alexandre, H.; Tristezza, M.; Durante, M.; Tufariello, M.; Grieco, F.; Spano, G. Intraspecific biodiversity and ‘spoilage potential’ of Brettanomyces bruxellensis in Apulian wines. LWT Food Sci. Technol. 2015, 60, 102–108. [Google Scholar] [CrossRef]
- Tufariello, M.; Maiorano, G.; Rampino, P.; Spano, G.; Grieco, F.; Perrotta, C.; Capozzi, V.; Grieco, F. Selection of an autochthonous yeast starter culture for industrial production of Primitivo “Gioia del Colle” PDO/DOC in Apulia (Southern Italy). LWT 2019, 99, 188–196. [Google Scholar] [CrossRef]
- Grieco, F.; Carluccio, M.A.; Giovinazzo, G. Autochthonous Saccharomyces cerevisiae Starter Cultures Enhance Polyphenols Content, Antioxidant Activity, and Anti-Inflammatory Response of Apulian Red Wines. Foods 2019, 8, 453. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Ethanol | Sugars | TA | VA | pH | Malic | Lactic | Glycerol |
|---|---|---|---|---|---|---|---|---|
| 1 | 12.14 ± 1.12 | 1.13 ± 0.20 | 9.38 ± 1.24 | 0.22 ± 0.02 | 3.00 ± 0.15 | 2.17 b ± 0.61 | 0.01 a ± 0.008 | 8.17 ± 1.40 |
| 2 | 12.23 ± 1.20 | 1.50 ± 0.49 | 9.14 ± 1.11 | 0.30 ± 0.01 | 3.08 ± 0.05 | 2.00 b ± 0.04 | nd | 9.86 ± 1.42 |
| 3 | 12.65 ± 1.44 | 1.36 ± 0.16 | 9.36 ± 1.17 | 0.23 ± 0.07 | 3.11 ± 0.21 | 0.02 a ± 0.00 | 2.64 b ± 0.33 | 8.55 ± 1.42 |
| 4 | 12.00 ± 1.25 | 1.66 ± 0.21 | 9.29 ± 1.33 | 0.29 ± 0.06 | 3.09 ± 0.15 | 0.02 a ± 0.01 | 2.00 b ± 0.25 | 9.13 ± 1.34 |
| Compounds | Trial 1 | Trial 2 | Trial 3 | Trial 4 | Odour Thresh Old | Sensory Notes | Odorant Series |
|---|---|---|---|---|---|---|---|
| Alcohols | |||||||
| 1-Propanol | 0.20 a ± 0.04 | 0.15 a ± 0.05 | nd | 0.31 a ± 0.07 | 306 | Ripe fruit, alcohol | |
| 2-Methyl-1-propanol | 1.85 a ± 0.30 | 1.56 a ± 0.35 | 1.47 a ± 0.24 | 2.84 a ± 0.65 | 0.2 | Bitter, green, harsh | 1 |
| 1-Butanol | 0.07 a ± 0.02 | nd | 0.06 a ± 0.02 | 0.07 a ± 0.02 | 150 | ||
| 3-Methyl-1-butanol | 26.82 a ± 5.48 | 18.43 a ± 4.90 | 32.23 a ± 4.67 | 45.51 a ± 8.97 | 30 | Vinous fusel alcohol | 2 |
| 3-Methyl-pentanol | 0.55 a ± 0.12 | 0.76 a ± 0.13 | 0.11 a ± 0.03 | 0.95 a ± 0.23 | 1.1 | Pungent, cocoa, wine-like | |
| 1-Hexanol | 0.47 a ± 0.07 | 0.34 a ± 0.06 | 0.41 a ± 0.06 | 0.56 a ± 0.11 | 8 | Green | |
| 3-Hexen-1-ol (Z) | 0.02 a ± 0.005 | nd | nd | 0.03 a ± 0.005 | 1 | Herbaceous, green | |
| 3-Hexen-1-ol (E) | 0.07 a ± 0.005 | 0.07 a ± 0.02 | 0.07 a ± 0.03 | 0.10 a ± 0.02 | 15 | Green | |
| Benzyl alcohol | 0.07 a ± 0.02 | 0.07 a ± 0.03 | 0.08 a ± 0.02 | 0.10a ± 0.02 | 900 | Burning taste | |
| Phenylethanol | 39.07 a ± 7.11 | 33.23 a ± 4.58 | 40.57 a ± 7.21 | 59.50 b ± 9.55 | 14 | Rose floral | 3 |
| Esters | |||||||
| Ethyl butanoate | 0.25 a ± 0.02 | 0.38 a ± 0.08 | 0.55 a ± 0.11 | 0.72 a ± 0.13 | 0.02 | Fruity apple | 4 |
| Isoamyl acetate | 0.26 a ± 0.06 | 0.28 a ± 0.02 | 0.55 a ± 0.03 | 0.73a ± 0.13 | 0.03 | Banana | 4 |
| Ethyl hexanoate | 0.59 a ± 0.02 | 0.64 a ± 0.02 | 0.60 a ± 0.03 | 0.95 a ± 0.12 | 0.014 | Green apple, anise | 1,4 |
| Ethyl lactate | 0.14 a ± 0.03 | 0.08 a ± 0.02 | 1.13 b ± 0.03 | 1.16 b ± 0.03 | 150 | ||
| Ethyl octanoate | 0.38 a ± 0.12 | 0.46 a ± 0.14 | 0.57 a ± 0.13 | 0.67 a ± 0.14 | 0.005 | Sweet, fruity, fresh | 4 |
| Ethyl-3-hydroxy butanoate | 0.04 a ± 0.005 | 0.03 a ± 0.006 | 0.03 a ± 0.004 | 0.032 a ± 0.005 | 1 | Fruity, grape | 4 |
| Ethyl decanoate | 0.67 a ± 0.12 | 0.55 a ± 0.10 | 0.76 a ± 0.25 | 1.10 a ± 0.22 | 0.2 | Fruity, sweet, grape | 4 |
| Diethyl succinate | 0.31 a ± 0.04 | 0.11 a ± 0.05 | 0.22 a ± 0.006 | 0.13 a ± 0.03 | 6 | Wine | |
| 3-Hydroxy ethyl hexanoate | nd | 0.017 a ± 0.005 | nd | 0.028 a ± 0.04 | NA | ||
| 1,3-propandiol acetate | 0.07 a ± 0.02 | nd | 0.09 a ± 0.02 | nd | NA | ||
| 2-Phenethylacetate | 0.13 a ± 0.05 | 0.33 a ± 0.02 | 0.45 a ± 0.06 | 0.95a ± 0.15 | 0.25 | Fruity | 4 |
| Diethyl malate | 0.06 a ± 0.02 | 0.06 a ± 0.02 | 0.07 a ± 0.02 | 0.10 a ± 0.03 | 10 | Fruity | |
| Monoethyl succinate | 1.93 b ± 0.45 | 0.66 a ± 0.07 | 1.44 b ± 0.54 | 1.61 b ± 0.44 | NA | ||
| Terpenes | |||||||
| Linalol | 0.24 a ± 0.06 | 0.32 a ± 0.10 | nd | 0.65 a ± 0.20 | 0.025 | Floreal | 3 |
| Volatile acids | |||||||
| 2-Methylpropanoic acid | 0.20 a ± 0.05 | 0.13 a ± 0.04 | 0.24 a ± 0.05 | 0.25 a ± 0.06 | NA | ||
| Butanoic acid | nd | nd | 0.09 ± 0.02 | nd | 2.2 | Cheesy | |
| 2-Methyl hexanoic acid | 0.92 a ± 0.11 | 0.24 a ± 0.06 | 0.90 a ± 0.16 | 0.34 a ± 0.05 | |||
| Hexanoic acid | 1.23 b ± 0.06 | 0.35 a ± 0.08 | 1.15 b ± 0.35 | 0.40 a ± 0.06 | 0.42 | Fatty acid, cheese | 5 |
| Octanoic acid | 2.28 b ± 0.55 | 0.55 a ± 0.16 | 2.14 b ± 0.07 | 0.54 a ± 0.10 | 0.5 | Fatty acid, cheese | 5 |
| n-Decanoic acid | 0.62 a ± 0.07 | nd | 0.41 a ± 0.07 | nd | 1 | ||
| 9-Decenoic acid | 0.69 a ± 0.18 | nd | 0.50 a ± 0.06 | nd | NA | ||
| Lactones | |||||||
| Butyrolactone | 0.25 a ± 0.03 | 0.17 a ± 0.03 | nd | 0.33 a ± 0.07 | NA |
| Trials | Ethanol | Sugars | TA | VA | pH | Malic | Lactic | Glycerol |
|---|---|---|---|---|---|---|---|---|
| Pilot A | 12.13 a ± 1.61 | 0.27 a ± 0.09 | 6.65 b ± 0.34 | 0.27 a ± 0.01 | 3.38 a ± 0.12 | 1.74 a ± 0.027 | 0.13 a ± 0.09 | 6.21 a ± 1.41 |
| Pilot B | 12.22 a ± 1.23 | 1.12 b ± 0.06 | 4.92 a ± 0.62 | 0.38 a ± 0.02 | 3.53 a ± 0.14 | nd | 1.38 b ± 0.16 | 6.58 a ± 1.42 |
| Pilot C | 12.34 a ± 1.11 | 0.71 a ± 0.09 | 5.20 a ± 0.21 | 0.30 a ± 0.02 | 3.52 a ± 0.11 | 1.75 a ± 0.027 | 0.14 a ± 0.04 | 8.32 b ± 1.25 |
| Pilot D | 13.50 b ± 1.80 | 0.11 a ± 0.04 | 5.12 a ± 0.16 | 0.33 a ± 0.01 | 3.55 a ± 0.13 | nd | 1.78 b ± 0.12 | 8.20 b ± 1.45 |
| Compounds | Pilot A | Pilot B | Pilot C | Pilot D |
|---|---|---|---|---|
| Alcohols | ||||
| 2-Methyl-1-propanol | 3.17 a ± 0.04 | 3.70 a ± 0.05 | 2.90 a ± 0.025 | 4.80 a ± 0.34 |
| 3-Methyl-1-butanol | 12.34 a ± 3.55 | 33.67 a ± 4.76 | 26.54 a ± 4.55 | 40.28 a ± 6.11 |
| 1-Hexanol | 0.90 a ± 0.11 | 2.73 b ± 0.44 | 0.48 a ± 0.07 | 2.20 b ± 0.47 |
| 3-Hexen-1-ol (Z) | 0.020 a ± 0.007 | 0.045 a ± 0.005 | nd | 0.05 a ± 0.02 |
| 3-Hexen-1-ol (E) | nd | 0.22 a ± 0.06 | nd | 0.10 a ± 0.03 |
| 1-Heptanol | 0.04 a ± 0.02 | 0.15 b ± 0.04 | 0.21 b ± 0.07 | nd |
| Benzyl alcohol | 8.21 a ± 2.10 | 21.67 b ± 3.93 | 17.45 b ± 4.10 | 33.23 b ± 7.36 |
| Phenylethanol | 16.20 a ± 4.43 | 30.70 b ± 5.33 | 21.20 a ± 4.38 | 48.11 b ± 4.17 |
| Esters | ||||
| Ethyl b utanoate | 0.45 a ± 0.11 | 0.52 a ± 0.16 | 0.45 a ± 0.12 | 0.95 a ± 0.14 |
| Isoamyl acetate | 0.18 a ± 0.05 | 0.50 a ± 0.06 | 0.67 a ± 0.13 | 1.20 a ± 0.26 |
| Ethyl hexanoate | 0.19 a ± 0.05 | 0.49 a ± 0.05 | 0.58 a ± 0.11 | 1.50 a ± 0.12 |
| Ethyl lactate | nd | 1.54 a ± 0.15 | nd | 2.22 a ± 0.58 |
| Ethyl Octanoate | 0.27 a ± 0.06 | 0.22 a ± 0.06 | 0.25 a ± 0.06 | 0.52 a ± 0.15 |
| Diethyl succinate | 2.38 ab ± 0.06 | 4.40 b ± 0.55 | 0.62 a ± 0.06 | 8.68 c ± 2.58 |
| 2-Phenethylacetate | 0.06 a ± 0.02 | 0.22 a ± 0.06 | 0.05 a ± 0.02 | 2.15 b ± 0.56 |
| Hydroxy diethyl malate | nd | 1.56 a ± 0.21 | nd | 1.40 a ± 0.60 |
| Volatile acids | ||||
| 2-Methylpropanoic acid | 0.45 a ± 0.11 | 0.37 a ± 0.05 | 0.41 a ± 0.11 | 0.20 a ± 0.05 |
| 2-Methyl hexanoic acid | nd | 0.22 a ± 0.06 | 0.43 a ± 0.01 | 0.68 a ± 0.11 |
| Hexanoic acid | 0.48 a ± 0.07 | 0.73 a ± 0.45 | 0.27 a ± 0.06 | 0.20 a ± 0.05 |
| Octanoic acid | 1.09 a ± 0.34 | 1.42 a ± 0.74 | 0.72 a ± 0.12 | 0.17 a ± 0.04 |
| Decanoic acid | 0.12 a ± 0.03 | 0.82 a ± 0.12 | 0.11 a ± 0.03 | 0.56 a ± 0.11 |
| Terpenes | ||||
| m-Cymene | 0.12 a ± 0.05 | 0.42 a ± 0.05 | 0.28 a ± 0.10 | 0.56 a ± 0.06 |
| α-Terpineol | 0.11 a ± 0.04 | 0.34 a ± 0.05 | 0.25 a ± 0.04 | 0.47 a ± 0.02 |
| 3,7-Dimetil-1,7-Octanediol | 1.11 a ± 0.03 | 1.37 a ± 0.11 | 0.95 a ± 0.11 | 2.20 a ± 0.65 |
| Sulphur compound | ||||
| Methyonol | 0.13 a ± 0.01 | nd | nd | 0.28 a ± 0.06 |
| Volatile phenol | ||||
| 4-Vinyl guaiacol | nd | 0.31 a ± 0.06 | 0.22 a ± 0.06 | nd |
| Ethanol | Sugar | TA | VA | pH | Malic | Lactic | Glycerol | |
|---|---|---|---|---|---|---|---|---|
| Mixstart_2017 | 11.97 a ± 0.55 | 0.04 a ± 0.01 | 6.32 a ± 0.20 | 0.34 a ± 0.05 | 3.64 a ± 0.23 | nd | 2.83 b ± 0.05 | 9.79 a ± 0.56 |
| Comm 1_ 2017 | 12.06 a ± 1.54 | 0.86c ± 0.14 | 6.86 a ± 0.13 | 0.41 a ± 0.06 | 3.37 a ± 0.21 | nd | 1.70 a ± 0.21 | 9.66 a ± 0.54 |
| Comm 2_ 2017 | 12.30 a ± 2.10 | 0.65 b ± 0.11 | 6.64 a ± 1.10 | 0.49 a ± 0.05 | 3.49 a ± 0.43 | nd | 1.41 a ± 0.05 | 10.66 a ± 1.10 |
| Comm 3_ 2017 | 11.92 a ± 1.11 | nd | 6.35 a ± 0.54 | 0.38 a ± 0.04 | 3.56 a ± 0.23 | nd | 1.22 a ± 0.04 | 10.44 a ± 0.17 |
| Mixstart_2018 | 13.69 a ± 2.13 | 0.20 a ± 0.02 | 6.51 a ± 0.23 | 0.23 a ± 0.06 | 3.68 a ± 0.16 | nd | 2.65 b ± 0.21 | 10.48 a ± 0.22 |
| Comm 1_ 2018 | 13.40 a ± 0.23 | 0.99 b ± 0.20 | 6.17 a ± 1.55 | 0.31 a ± 0.05 | 3.62 a ± 0.05 | nd | 1.75 a ± 0.12 | 10.18 a ± 1.45 |
| Comm 2_ 2018 | 13.80 a ± 0.33 | 0.52 c ± 0.05 | 6.75 a ± 1.30 | 0.35 a ± 0.05 | 3.60 a ± 0.30 | 0.26 ± 0.07 | 1.73 a ± 0.11 | 10.92 a ± 0.55 |
| Comm 3_ 2018 | 13.44 a ± 0.55 | 0.67 c ± 0.11 | 6.54 a ± 0.54 | 0.38 a ± 0.05 | 3.65 a ± 0.43 | nd | 1.94 a ± 0.14 | 11.80 a ± 0.50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tufariello, M.; Capozzi, V.; Spano, G.; Cantele, G.; Venerito, P.; Mita, G.; Grieco, F. Effect of Co-Inoculation of Candida zemplinina, Saccharomyces cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms 2020, 8, 726. https://doi.org/10.3390/microorganisms8050726
Tufariello M, Capozzi V, Spano G, Cantele G, Venerito P, Mita G, Grieco F. Effect of Co-Inoculation of Candida zemplinina, Saccharomyces cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms. 2020; 8(5):726. https://doi.org/10.3390/microorganisms8050726
Chicago/Turabian StyleTufariello, Maria, Vittorio Capozzi, Giuseppe Spano, Giovanni Cantele, Pasquale Venerito, Giovanni Mita, and Francesco Grieco. 2020. "Effect of Co-Inoculation of Candida zemplinina, Saccharomyces cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy)" Microorganisms 8, no. 5: 726. https://doi.org/10.3390/microorganisms8050726