
Molecular Epidemiology and Characterization of Carbapenem-Resistant Klebsiella pneumoniae Isolated from Urine at a Teaching Hospital in Taiwan


Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Isolates of K. Pneumoniae
2.2. Antimicrobial Susceptibility Test
2.3. Genomic DNA Extraction
2.4. Multilocus Sequence Typing and Relatedness Analysis
2.5. Detection of Extended-Spectrum β-Lactamase Genes and Carbapenemases
2.6. Sequence Analysis of the Outer Membrane Proteins OmpK35 and OmpK36
2.7. Measurement of Gene Expression
2.8. Fluorescent Dye Accumulation Assay
3. Results
3.1. Carbapenem-Resistant K. pneumoniae from Urine
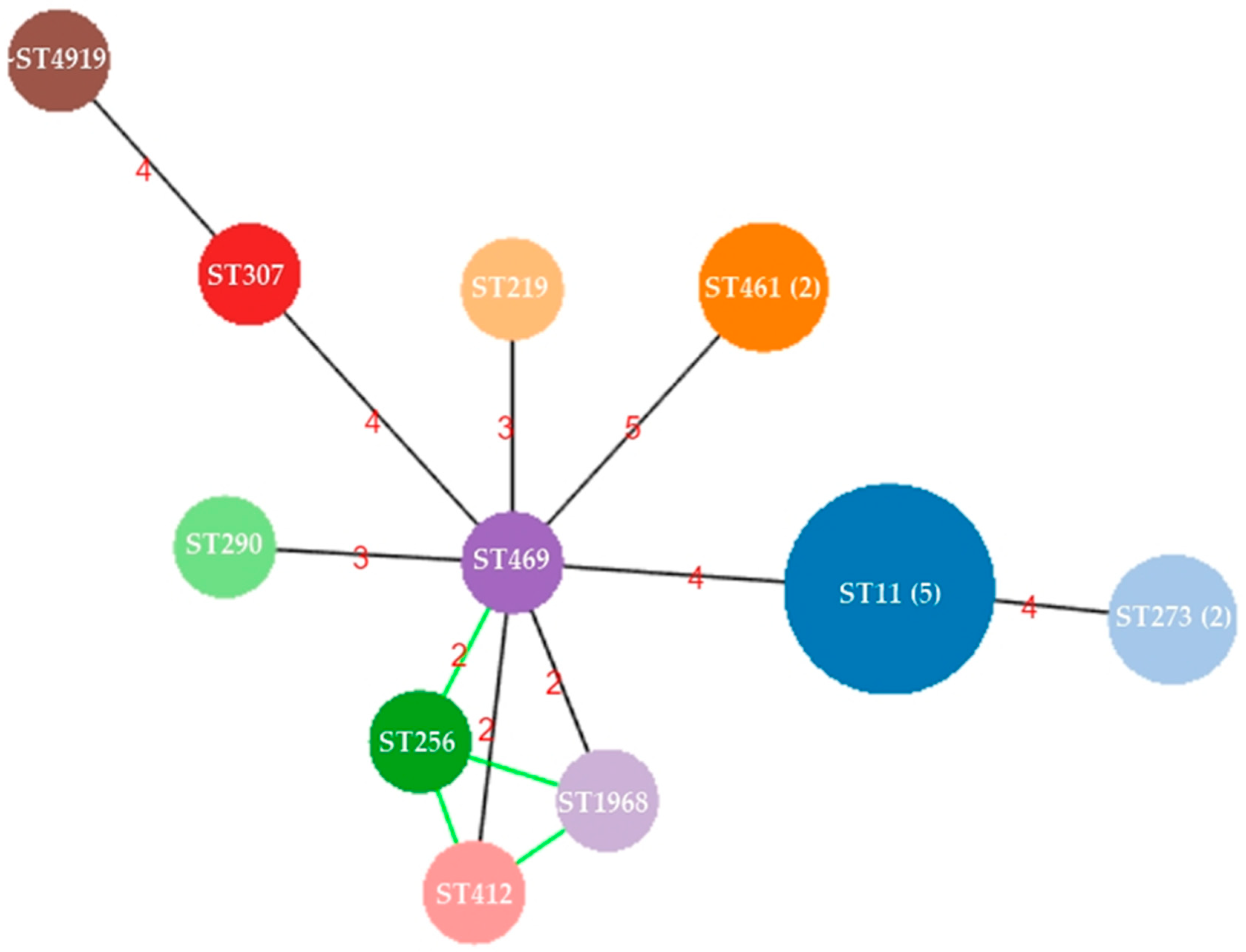
3.2. Capsule Types and Population Structure of CRKP from Urine
3.3. Detection of Acquired β-Lactamases and Mutation Analysis of OmpK35 and OmpK36
3.4. Role of Efflux Pumps in Carbapenem Resistance
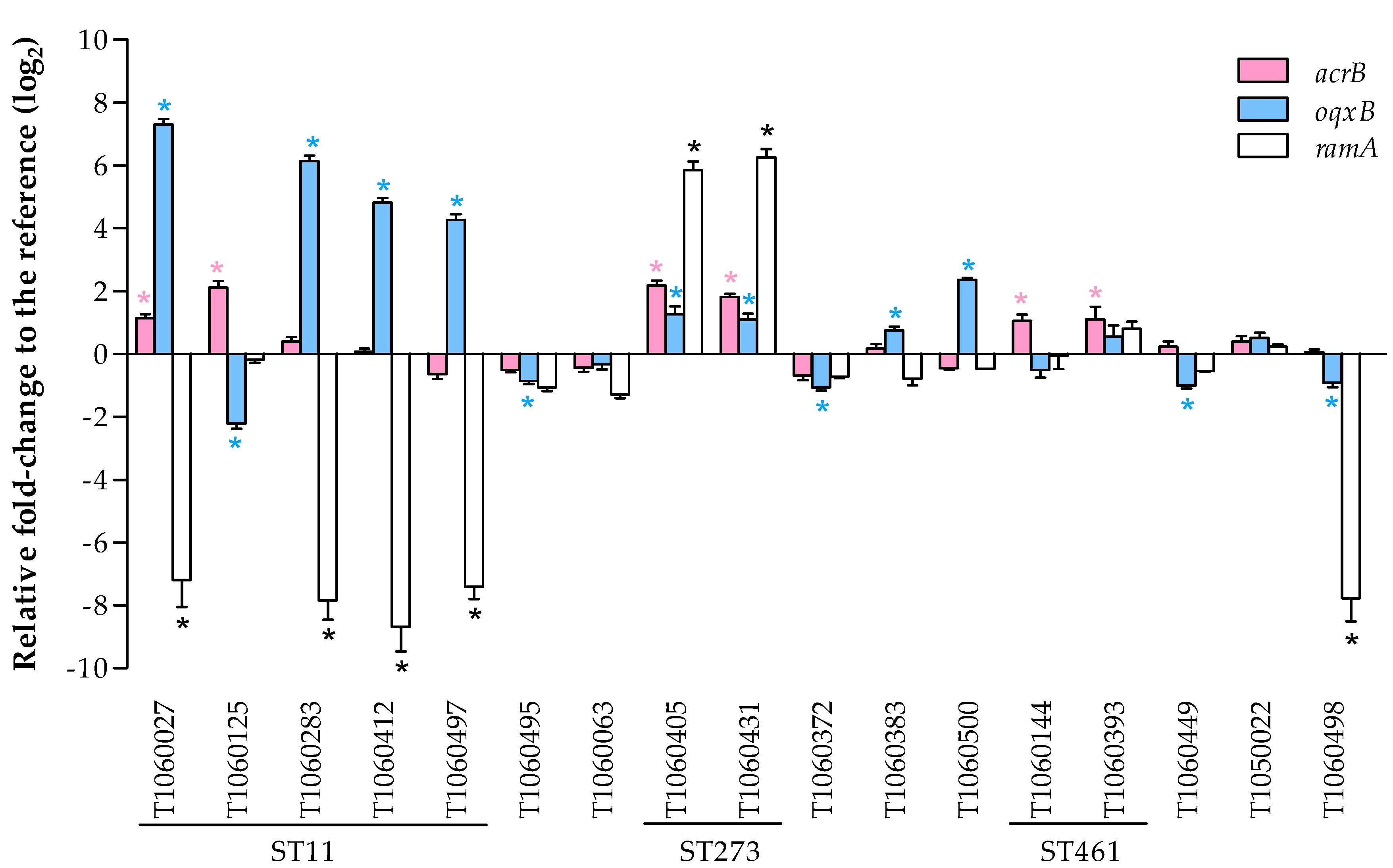
3.4.1. Expression of Efflux Pumps and the Related Regulator RamA
3.4.2. Membrane Permeability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, R.; Murt, A. Epidemiology of urological infections: A global burden. World J. Urol. 2020, 38, 2669–2679. [Google Scholar] [CrossRef] [PubMed]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef]
- Tandogdu, Z.; Wagenlehner, F.M.E. Global epidemiology of urinary tract infections. Curr. Opin. Infect. Dis. 2016, 29, 73–79. [Google Scholar] [CrossRef]
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1. [Google Scholar] [CrossRef]
- Li, Y.; Shen, H.; Zhu, C.; Yu, Y. Carbapenem-resistant Klebsiella pneumoniae infections among ICU admission patients in central China: Prevalence and prediction model. BioMed Res. Int. 2019, 2019, 9767313. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria-To Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 30 April 2020).
- Logan, L.K.; Weinstein, R.A. The Epidemiology of carbapenem-resistant Enterobacteriaceae: The impact and evolution of a global menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.; Laupland, K.B. Extended-spectrum beta-lactamase-producing Enterobacteriaceae: An emerging public-health concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.T.; Wu, U.I.; Lauderdale, T.L.; Chen, M.C.; Li, S.Y.; Hsu, L.Y.; Chang, S.C. Carbapenem-nonsusceptible Enterobacteriaceae in Taiwan. PLoS ONE 2015, 10, e0121668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.P.; Wang, J.T.; Chang, S.C.; Chang, F.Y.; Fung, C.P.; Chuang, Y.C.; Chen, Y.S.; Shiau, Y.R.; Tan, M.C.; Wang, H.Y.; et al. The antimicrobial susceptibility of Klebsiella pneumoniae from community settings in Taiwan, a trend analysis. Sci. Rep. 2016, 6, 36280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, S.K.; Chan, M.C.; Huang, L.Y.; Lin, Y.T.; Lin, J.C.; Lu, P.L.; Siu, L.K.; Chang, F.Y.; Yeh, K.M. Tigecycline resistance among carbapenem-resistant Klebsiella pneumoniae: Clinical characteristics and expression levels of efflux pump genes. PLoS ONE 2017, 12, e0175140. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [Green Version]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decre, D. wzi Gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-Goncalves, B.; Francisco, A.P.; Vaz, C.; Ramirez, M.; Carrico, J.A. PHYLOViZ Online: Web-based tool for visualization, phylogenetic inference, analysis and sharing of minimum spanning trees. Nucleic Acids Res. 2016, 44, W246–W251. [Google Scholar] [CrossRef]
- Yan, J.J.; Wang, M.C.; Zheng, P.X.; Tsai, L.H.; Wu, J.J. Associations of the major international high-risk resistant clones and virulent clones with specific ompK36 allele groups in Klebsiella pneumoniae in Taiwan. New Microbes New Infect. 2015, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Jimenez-Castellanos, J.C.; Wan Ahmad Kamil, W.N.; Cheung, C.H.; Tobin, M.S.; Brown, J.; Isaac, S.G.; Heesom, K.J.; Schneiders, T.; Avison, M.B. Comparative effects of overproducing the AraC-type transcriptional regulators MarA, SoxS, RarA and RamA on antimicrobial drug susceptibility in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2016, 71, 1820–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialek-Davenet, S.; Lavigne, J.P.; Guyot, K.; Mayer, N.; Tournebize, R.; Brisse, S.; Leflon-Guibout, V.; Nicolas-Chanoine, M.H. Differential contribution of AcrAB and OqxAB efflux pumps to multidrug resistance and virulence in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2015, 70, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Liu, L.; Zhou, H.; Chan, E.W.; Li, J.; Fang, Y.; Li, Y.; Liao, K.; Chen, S. Nationwide surveillance of clinical carbapenem-resistant Enterobacteriaceae (CRE) strains in China. EBioMedicine 2017, 19, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.J.; Lin, T.L.; Lin, Y.T.; Su, P.A.; Chen, C.T.; Hsieh, P.F.; Hsu, C.R.; Chen, C.C.; Hsieh, Y.C.; Wang, J.T. Identification of capsular types in carbapenem-resistant Klebsiella pneumoniae strains by wzc sequencing and implications for capsule depolymerase treatment. Antimicrob. Agents Chemother. 2015, 59, 1038–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Xiao, T.; David, S.; Wang, Q.; Zhou, Y.; Guo, L.; Aanensen, D.; Holt, K.E.; Thomson, N.R.; Grundmann, H.; et al. Novel subclone of carbapenem-resistant Klebsiella pneumoniae sequence type 11 with enhanced virulence and transmissibility, China. Emerg. Infect. Dis. 2020, 26, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biehl, L.M.; Schmidt-Hieber, M.; Liss, B.; Cornely, O.A.; Vehreschild, M.J. Colonization and infection with extended spectrum beta-lactamase producing Enterobacteriaceae in high-risk patients—Review of the literature from a clinical perspective. Crit. Rev. Microbiol. 2016, 42, 1–16. [Google Scholar] [CrossRef]
- Coudeyras, S.; Nakusi, L.; Charbonnel, N.; Forestier, C. A tripartite efflux pump involved in gastrointestinal colonization by Klebsiella pneumoniae confers a tolerance response to inorganic acid. Infect. Immun. 2008, 76, 4633–4641. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, W.; Onishi, M.; Ni, R.; Tsuchiya, T.; Kuroda, T. Functional study of the novel multidrug efflux pump KexD from Klebsiella pneumoniae. Gene 2012, 498, 177–182. [Google Scholar] [CrossRef]
- Ni, R.T.; Onishi, M.; Mizusawa, M.; Kitagawa, R.; Kishino, T.; Matsubara, F.; Tsuchiya, T.; Kuroda, T.; Ogawa, W. The role of RND-type efflux pumps in multidrug-resistant mutants of Klebsiella pneumoniae. Sci. Rep. 2020, 10, 10876. [Google Scholar] [CrossRef]
- Webber, M.A.; Piddock, L.J. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Huang, T.W.; Juan, C.H.; Chou, S.H.; Tseng, Y.Y.; Chen, T.W.; Yang, T.C.; Lin, Y.T. Tigecycline-non-susceptible hypervirulent Klebsiella pneumoniae strains in Taiwan. J. Antimicrob. Chemother. 2020, 75, 309–317. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Fu, Y.; Chen, Q.; Ruan, Z.; Hua, X.; Zhou, H.; Yu, Y. Tigecycline susceptibility and the role of efflux pumps in tigecycline resistance in KPC-producing Klebsiella pneumoniae. PLoS ONE 2015, 10, e0119064. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, J.; Zhu, Z.; Xu, T.; Sheng, Z.K.; Ye, M.; Xu, X.; Wang, M. Efflux pumps AcrAB and OqxAB contribute to nitrofurantoin resistance in an uropathogenic Klebsiella pneumoniae isolate. Int. J. Antimicrob. Agents 2019, 54, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Spengler, G.; Kincses, A.; Gajdács, M.; Amaral, L. New roads leading to old destinations: Efflux pumps as targets to reverse multidrug resistance in bacteria. Molecules 2017, 22, 468. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Sanz-García, F.; Hernando-Amado, S.; Martínez, J.L.; Alcalde-Rico, M. The development of efflux pump inhibitors to treat Gram-negative infections. Expert Opin. Drug Discov. 2018, 13, 919–931. [Google Scholar] [CrossRef]
- Usai, D.; Donadu, M.; Bua, A.; Molicotti, P.; Zanetti, S.; Piras, S.; Corona, P.; Ibba, R.; Carta, A. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J. Infect. Dev. Ctries. 2019, 13, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Hsieh, Y.H.; Powers, Z.M.; Kao, C.Y. Defeating antibiotic-resistant bacteria: Exploring alternative therapies for a post-antibiotic era. Int. J. Mol. Sci. 2020, 21, 1061. [Google Scholar] [CrossRef] [Green Version]
- Owen, L.; Laird, K. Synchronous application of antibiotics and essential oils: Dual mechanisms of action as a potential solution to antibiotic resistance. Crit. Rev. Microbiol. 2018, 44, 414–435. [Google Scholar] [CrossRef]
- Donadu, M.G.; Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, M.; Marchetti, M.; Sanna, G.; Madeddu, S.; et al. Phytochemical compositions and biological activities of essential oils from the leaves, rhizomes and whole plant of Hornstedtia bella Škorničk. Antibiotics 2020, 9, 334. [Google Scholar] [CrossRef]
- Le, N.T.; Donadu, M.G.; Ho, D.V.; Doan, T.Q.; Le, A.T.; Raal, A.; Usai, D.; Sanna, G.; Marchetti, M.; Usai, M.; et al. Biological activities of essential oil extracted from leaves of Atalantia sessiflora Guillauminin Vietnam. J. Infect. Dev. Ctries. 2020, 14, 1054–1064. [Google Scholar] [CrossRef]
- Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, D.; Madeddu, S.; Marchetti, M.; Usai, M.; Rappelli, P.; et al. In vitro antimicrobial activity of essential oil extracted from leaves of Leoheo domatiophorus Chaowasku, D.T. Ngo and H.T. Le in Vietnam. Plants 2020, 9, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, D.; Sanna, G.; Carta, A.; Rappelli, P.; Diaz, N.; et al. Biological activities of essential oils from leaves of Paramignya trimera (Oliv.) Guillaum and Limnocitrus littoralis (Miq.) Swingle. Antibiotics 2020, 9, 207. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Ward 4 | Age | Isolated Date | CMZ 1 | CAZ 1 | CRO 1 | FEP1 | IMP 1 | MEM 1 | ETP 1 | GEN 1 | AMK 1 | CIP 1 | LVX 1 | SXT 1 | TZP 1 | COL 1 | TGC 2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1050022 | Ward 7 | 68 | 3 December 2016 | R (≥32) | R (>16) | I (32) | S (≤2) | R (4) | S (≤1) | R (>4) | R (>8) | S (≤8) | S (≤0.5) | S (≤1) | R (>2/38) | R (>64/4) | WT (1) | S (0.5) |
| T1060027 | ER | 82 | 4 January 2017 | R (≥32) | I (8) | R (>32) | R (>16) | I 3 | S (≤1) | R (>4) | R (>8) | S (≤8) | R (>2) | R (>4) | S 3 | R (>64/4) | WT (1) | S (1) |
| T1060063 | ER | 88 | 13 February 2017 | R (≥32) | R (>16) | I (16) | S (≤2) | R (8) | I (2) | R (>4) | R (>8) | S (≤8) | S (1) | S (≤1) | R (>2/38) | R (>64/4) | WT (1) | S (0.19) |
| T1060125 | Ward 10B | 65 | 21 March 2017 | R (≥32) | R (>16) | R (>32) | R (>16) | R (> 8) | R (>8) | R (>4) | R (>8) | R (>32) | R (>2) | R (>4) | S (≤0.5/9.5) | R (>64/4) | WT (1) | S (0.5) |
| T1060144 | Ward 8B | 89 | 4 April 2017 | I (32) | R (>16) | R (>32) | R (>16) | R (8) | R (4) | R (4) | R (>8) | S (≤8) | S (1) | S (≤1) | R (>2/38) | R (>64/4) | WT (1) | S (0.75) |
| T1060283 | Ward 8A | 95 | 20 June 2017 | R (≥32) | R (>16) | R (>32) | R (>16) | I3 | S (≤1) | I (1) | R (>8) | R (>32) | R (>2) | R (>4) | R (>2/38) | R (>64/4) | WT (1) | S (0.75) |
| T1060372 | OPD | 54 | 2 August 2017 | R (≥32) | R (>16) | S (≤1) | S (≤1) | R (4) | S (≤0.25) | S (0.5) | R (>8) | S (≤8) | S (1) | S (≤1) | R (>2/38) | S (≤4/4) | WT (1) | S (1) |
| T1060383 | ER | 88 | 6 August 2017 | R (≥32) | R (>16) | S (≤1) | S (≤1) | I (2) | S (≤0.25) | S (≤0.25) | R (>8) | S (≤8) | R (>2) | R (>4) | S (≤0.5/9.5) | S (≤4/4) | WT (1) | S (0.38) |
| T1060393 | Ward 8B | 79 | 14 August 2017 | I (32) | R (>16) | R (>32) | R (>16) | R (> 4) | R (>4) | R (2) | R (>8) | R (>32) | S (1) | S (≤1) | R (>2/38) | R (≥64/4) | WT (≤0.25) | S (0.5) |
| T1060405 5 | ER | 81 | 17 August 2017 | R (≥32) | R (>16) | R (16) | S (≤1) | R (4) | S (≤0.25) | I (1) | S (≤2) | S (≤8) | R (>2) | R (>4) | R (>2/38) | I (64/4) | NWT (16) | R (2) |
| T1060412 | OPD | 93 | 15 August 2017 | R (≥32) | R (>16) | I (2) | S (2) | R (> 4) | R (4) | R (>4) | S (≤2) | S (16) | R (>2) | R (>4) | S (≤0.5/9.5) | R (≥64/4) | WT (≤0.25) | S (0.25) |
| T1060431 5 | ER | 81 | 26 August 2017 | R (≥32) | R (>16) | R (32) | S (2) | R (> 4) | I (2) | R (>4) | S (≤2) | S (≤8) | R (>2) | R (>4) | R (>2/38) | R (≥64/4) | NWT (8) | R (2) |
| T1060449 | Ward 8 | 55 | 5 September 2017 | R (≥32) | S (4) | S (≤1) | S (≤1) | R (4) | S (≤0.25) | S (0.5) | R (>8) | S (≤8) | S (1) | S (≤1) | R (>2/38) | S (≤4/4) | WT (0.5) | I (1.5) |
| T1060495 | OPD | 47 | 28 September 2017 | R (≥ 32) | R (>16) | R (>32) | I (8) | I (2) | S (≤0.25) | S (0.5) | R (>8) | S (≤8) | S (1) | S (≤1) | R (>2/38) | I (64/4) | WT (0.5) | S (0.75) |
| T1060497 | ER | 73 | 25 September 2017 | R (≥32) | I (8) | R (>32) | I (8) | I3 | S (≤0.25) | R (2) | S (≤2) | S (≤8) | R (>2) | R (>4) | S (≤0.5/9.5) | S (≤4/4) | WT (0.5) | S (0.75) |
| T1060498 | OPD | 86 | 20 September 2017 | R (≥32) | R (>16) | R (32) | S (≤1) | I 3 | R 3 | R (≥2) | R (>8) | S (≤8) | I (2) | S (≤1) | R (>2/38) | I (64/4) | WT (0.5) | S (0.25) |
| T1060500 | OPD | 64 | 22 September 2017 | R (≥32) | S (2) | S (≤1) | S (≤1) | R (4) | S (≤0.25) | S (≤0.25) | S (≤2) | S (≤8) | S (1) | S (≤1) | R (>2/38) | S (≤4/4) | WT (0.5) | I (1.5) |
| Susceptibility Frequency (%) | 0% (0/17) | 11.8% (2/17) | 23.5% (4/17) | 58.8% (10/17) | 0% (0/17) | 58.8% (10/17) | 29.4% (5/17) | 29.4% (5/17) | 82.4% (14/17) | 47.1% (8/17) | 52.9% (9/17) | 29.4% (5/17) | 29.4% (5/17) | 88.2% (15/17) | 76.5% (13/17) |
| Isolate Name | Sequence Types | Capsule Type | β-Lactamase 1 | OmpK35 2 | OmpK36 2 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| TEM | SHV | AmpC | CTX-M | Carbapenemase | Mutations | Type | Description | |||
| T1060027 | ST11 | KL64 | + | + | DHA | Group 9 | - | Ser233fsX | C | 135 amino acid deletion and frameshift |
| T1060125 | ST11 | KL47 | + | + | - | Group 9 | KPC | Asn29fsX | D | 2 amino acid deletion |
| T1060283 | ST11 | KL64 | + | + | DHA | Group 9 | - | Ser233fsX | A | WT |
| T1060412 | ST11 | KL64 | + | + | DHA | Group 9 | - | Ser233fsX | A | WT |
| T1060497 | ST11 | KL64 | + | + | DHA | Group 9 | - | Ser233fsX | A | WT |
| T1060495 | ST219 | KL114/125 | + | + | DHA | Group 1 | - | WT | C | Frameshift |
| T1060063 | ST256 | KL47 | + | + | DHA | - | - | WT | D | Frameshift |
| T1060405 | ST273 | KL15/17/51/52 | + | + | DHA | - | - | WT | A | 1 missense mutation (Val178Pro) |
| T1060431 | ST273 | KL15/17/51/52 | - | + | DHA | - | - | WT | A | 1 missense mutation (Val178Pro) |
| T1060372 | ST290 | KL21 | - | + | DHA | - | - | WT | A | 7 missense mutations and 1 amino acid insertion |
| T1060383 | ST307 | ~KL102/149/155 3 | - | + | DHA | - | - | Trp79X | D | WT |
| T1060500 | ST412 | KL57 | - | + | DHA | Group 9 | - | WT | C | Frameshift |
| T1060144 | ST461 | KL51 | - | + | - | Group 9 | VIM | WT | D | WT |
| T1060393 | ST461 | KL51 | + | + | - | Group 9 | VIM | WT | D | WT |
| T1060449 | ST469 | KL12 | + | + | DHA | - | - | WT | A | WT |
| T1050022 | ST1968 | KL102/149/155 | + | + | DHA | - | - | WT | D | WT |
| T1060498 | ~ST4919 /ST5154 3 | nd 4 | + | + | DHA | - | - | WT | C | IS insertion at N-terminal region |
| Prevalence (Positive/Total) | 70.6% (12/17) | 100% (17/17) | 82.3% (14/17) | 52.9% (9/17) | 17.6% (3/17) | 35.3% (6/17) | A: 7 C: 4 D: 6 | A: WT (4); a.a. substitutes (2) and insertion (1) C: frameshift (3); IS insertion (1) D: WT (4); frameshift (1); deletion (1) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-J.; Huang, C.-H.; Ilsan, N.A.; Lee, I.-H.; Huang, T.-W. Molecular Epidemiology and Characterization of Carbapenem-Resistant Klebsiella pneumoniae Isolated from Urine at a Teaching Hospital in Taiwan. Microorganisms 2021, 9, 271. https://doi.org/10.3390/microorganisms9020271
Lee Y-J, Huang C-H, Ilsan NA, Lee I-H, Huang T-W. Molecular Epidemiology and Characterization of Carbapenem-Resistant Klebsiella pneumoniae Isolated from Urine at a Teaching Hospital in Taiwan. Microorganisms. 2021; 9(2):271. https://doi.org/10.3390/microorganisms9020271
Chicago/Turabian StyleLee, Yuarn-Jang, Chih-Hung Huang, Noor Andryan Ilsan, I-Hui Lee, and Tzu-Wen Huang. 2021. "Molecular Epidemiology and Characterization of Carbapenem-Resistant Klebsiella pneumoniae Isolated from Urine at a Teaching Hospital in Taiwan" Microorganisms 9, no. 2: 271. https://doi.org/10.3390/microorganisms9020271