
Epidemiology and Characterization of CTX-M-55-Type Extended-Spectrum β-Lactamase-Producing Salmonella enterica Serovar Enteritidis Isolated from Patients in Shanghai, China

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
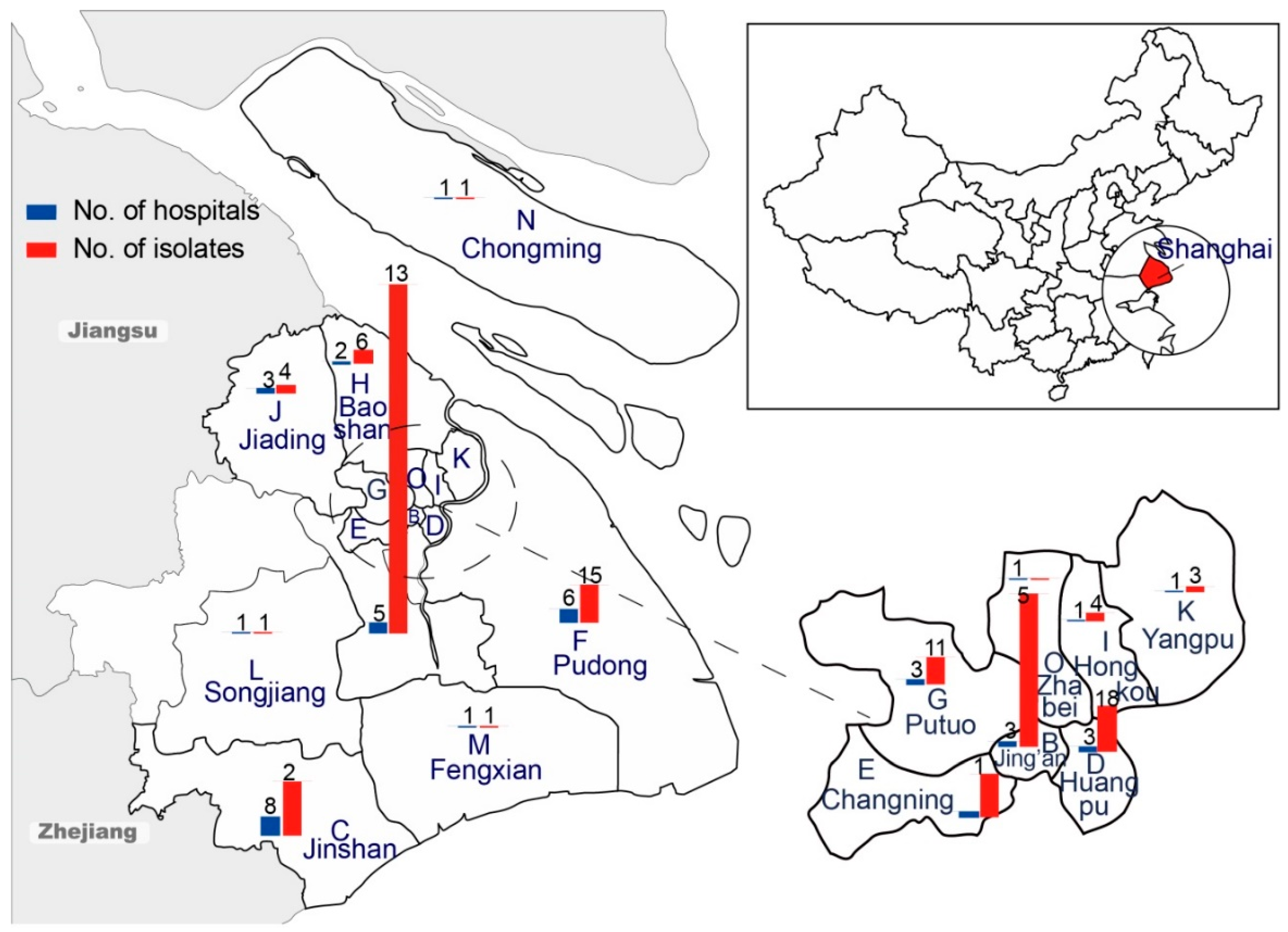
2.1. Bacterial Isolates
2.2. Antibiotic Susceptibility Testing
2.3. Detection of ESBL Phenotypes and Sequencing of ESBL-Encoding Genes
2.4. Pulsed-Field Gel Electrophoresis (PFGE)
2.5. Conjugation Experiment
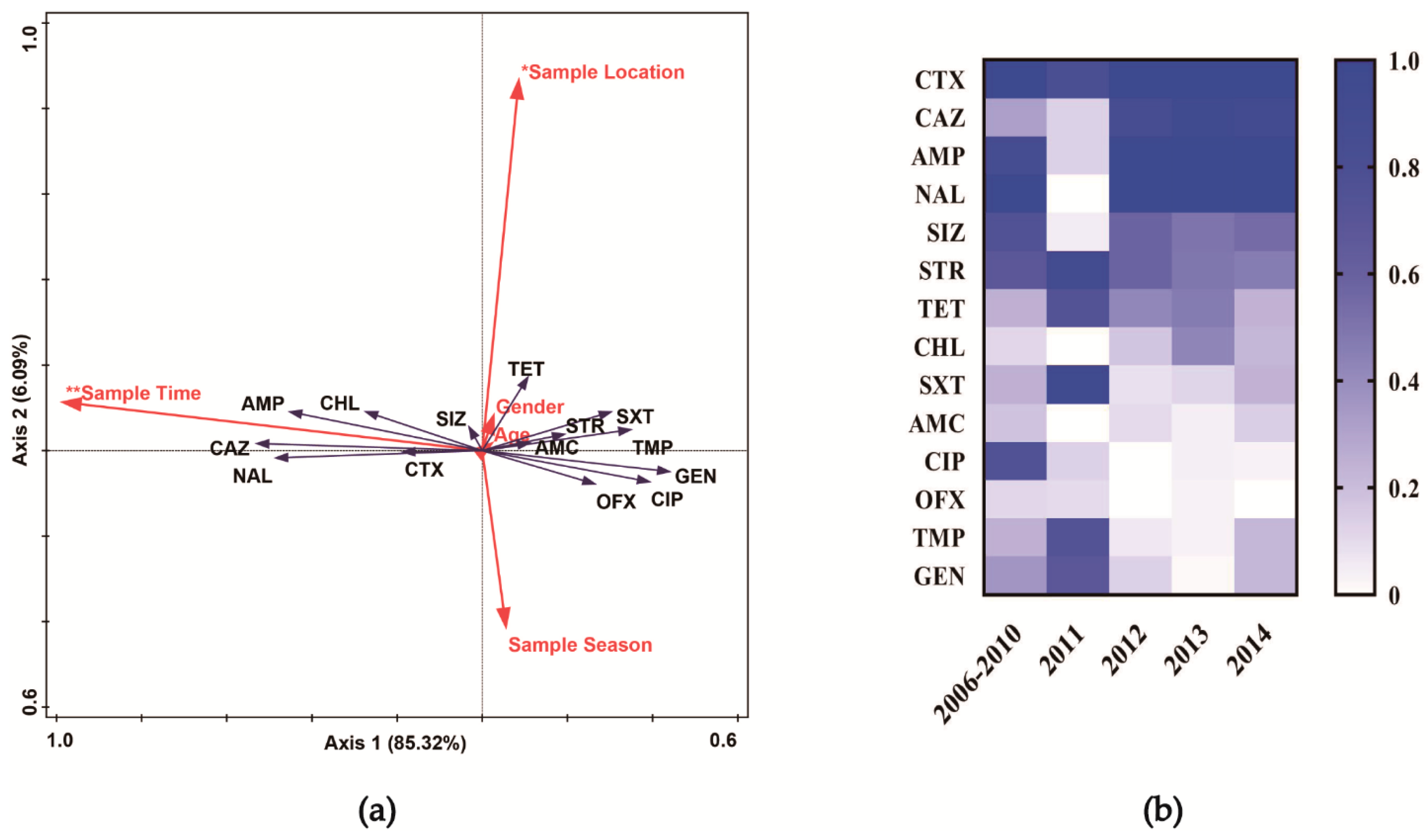
2.6. Statistical Analysis
3. Results
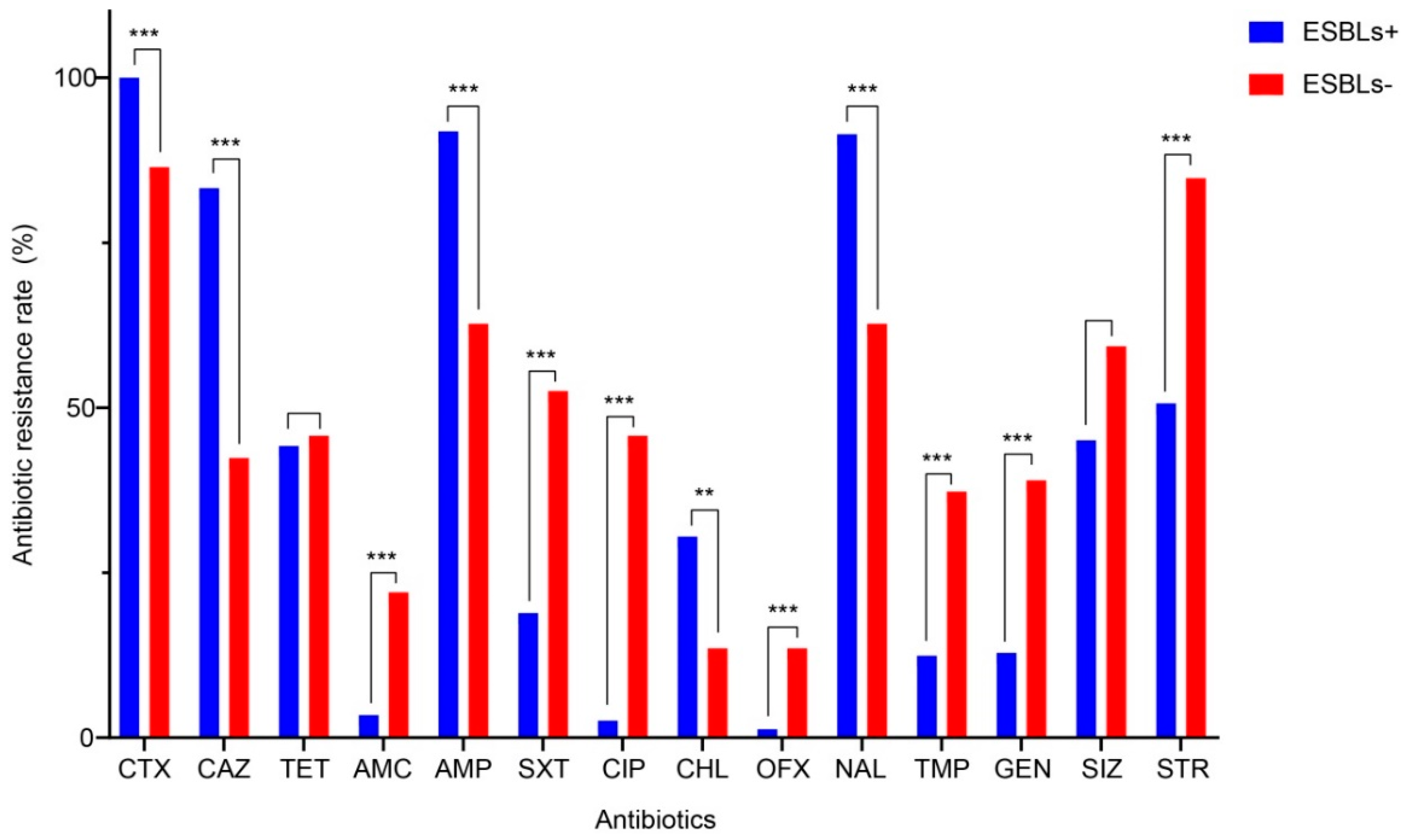
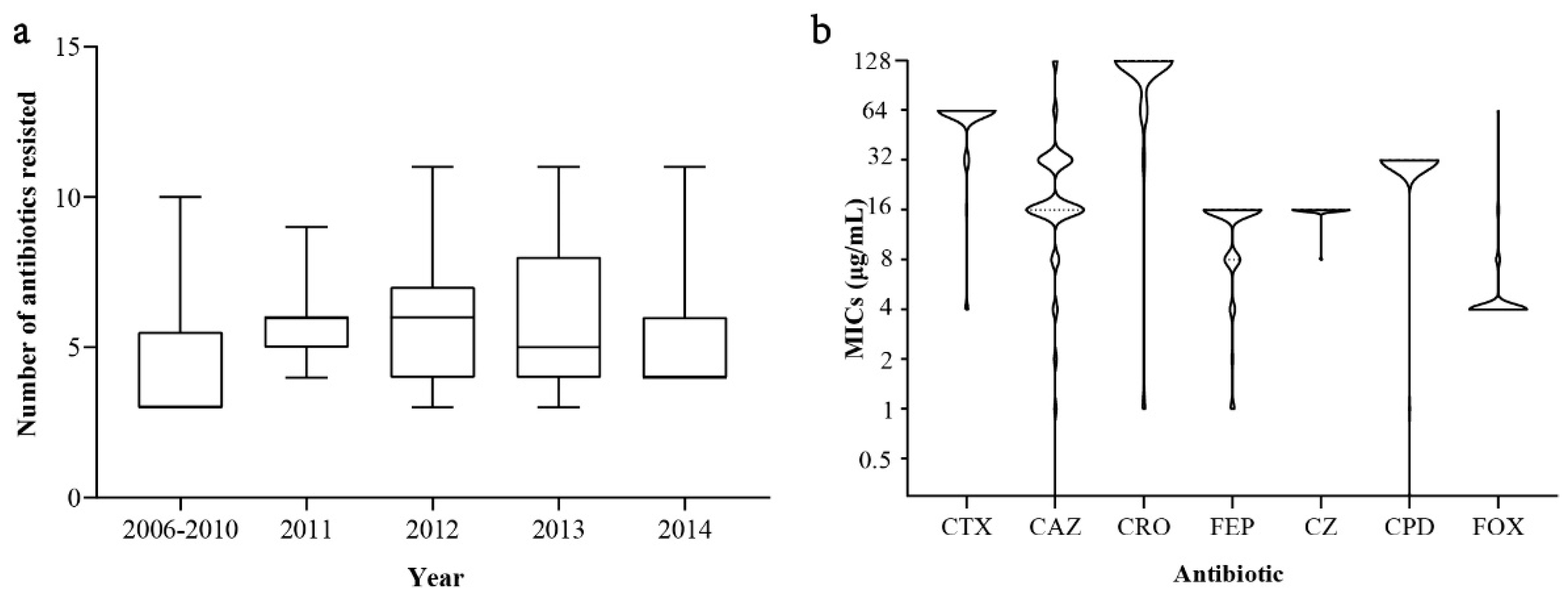
3.1. Antibiotic Susceptibility
3.2. Characterization of ESBL-SE
3.3. Prevalence of ESBL-Encoding Genes and Description of blaCTX-M-55-Positive ESBL-SE
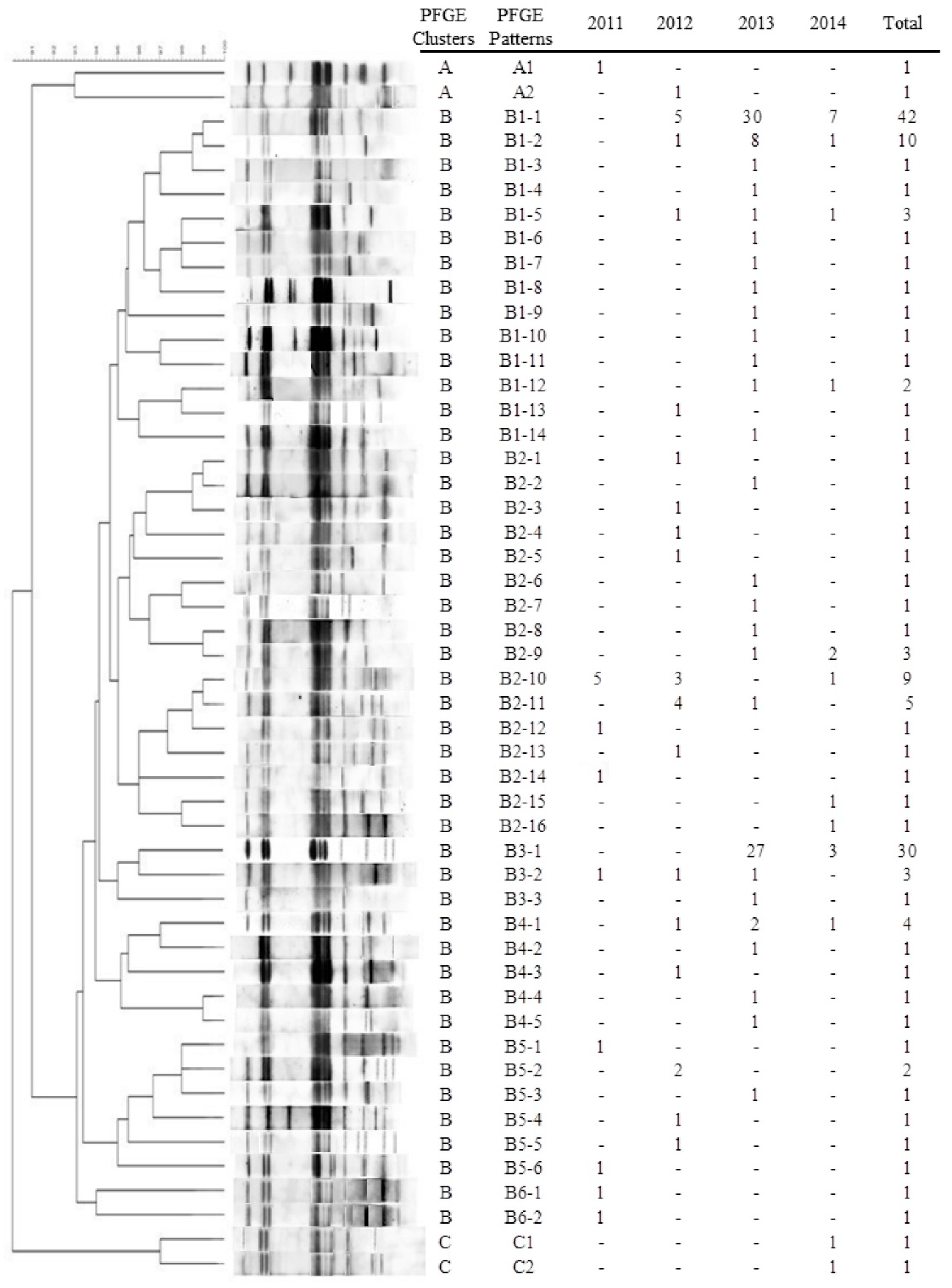
3.4. PFGE Patterns of blaCTX-M-55-Positive ESBL-SE
3.5. Conjugation of Three blaCTX-M-55-Postitive ESBL-SE
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yin, Y.; Zhou, D. Organoid and Enteroid Modeling of Salmonella Infection. Front. Cell Infect. Microbiol. 2018, 8, 102. [Google Scholar] [CrossRef]
- Gal-Mor, O. Persistent Infection and Long-Term Carriage of Typhoidal and Nontyphoidal Salmonellae. Clin. Microbiol. Rev. 2019, 32, e00088-00018. [Google Scholar] [CrossRef] [Green Version]
- Keestra-Gounder, A.M.; Tsolis, R.M.; Baumler, A.J. Now you see me, now you don’t: The interaction of Salmonella with innate immune receptors. Nat. Rev. Microbiol. 2015, 13, 206–216. [Google Scholar] [CrossRef]
- Shi, C.; Singh, P.; Ranieri, M.L.; Wiedmann, M.; Moreno Switt, A.I. Molecular methods for serovar determination of Salmonella. Crit. Rev. Microbiol. 2015, 41, 309–325. [Google Scholar] [CrossRef]
- Betancor, L.; Pereira, M.; Martinez, A.; Giossa, G.; Fookes, M.; Flores, K.; Barrios, P.; Repiso, V.; Vignoli, R.; Cordeiro, N.; et al. Prevalence of Salmonella enterica in poultry and eggs in Uruguay during an epidemic due to Salmonella enterica serovar Enteritidis. J. Clin. Microbiol. 2010, 48, 2413–2423. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.J.; Lappi, V.; Wolfgang, W.J.; Lapierre, P.; Palumbo, M.J.; Medus, C.; Boxrud, D. Characterization of Foodborne Outbreaks of Salmonella enterica Serovar Enteritidis with Whole-Genome Sequencing Single Nucleotide Polymorphism-Based Analysis for Surveillance and Outbreak Detection. J. Clin. Microbiol. 2015, 53, 3334–3340. [Google Scholar] [CrossRef] [Green Version]
- Kariuki, S.; Okoro, C.; Kiiru, J.; Njoroge, S.; Omuse, G.; Langridge, G.; Kingsley, R.A.; Dougan, G.; Revathi, G. Ceftriaxone-resistant Salmonella enterica serotype typhimurium sequence type 313 from Kenyan patients is associated with the blaCTX-M-15 gene on a novel IncHI2 plasmid. Antimicrob. Agents Chemother. 2015, 59, 3133–3139. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Li, Y.; Zhao, J.; Ma, H.; Wang, J.; Liang, B.; Du, X.; Wu, F.; Xia, S.; Yang, X.; et al. The phenotypic and molecular characteristics of antimicrobial resistance of Salmonella enterica subsp. enterica serovar Typhimurium in Henan Province, China. BMC Infect. Dis. 2020, 20, 511. [Google Scholar] [CrossRef]
- Katiyo, S.; Muller-Pebody, B.; Minaji, M.; Powell, D.; Johnson, A.P.; De Pinna, E.; Day, M.; Harris, R.; Godbole, G. Epidemiology and Outcomes of Nontyphoidal Salmonella Bacteremias from England, 2004 to 2015. J. Clin. Microbiol. 2019, 57, e01189-01118. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Chen, H.; Ou, Y.; Huang, T.; Chen, S.; Zhou, L.; Zhang, J.; Hu, Q.; Zhou, Y.; Ma, W. Prevalence, serotypes, and antimicrobial resistance of Salmonella isolates from patients with diarrhea in Shenzhen, China. BMC Microbiol. 2020, 20, 197. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly Prevalent Multidrug-Resistant Salmonella From Chicken and Pork Meat at Retail Markets in Guangdong, China. Front. Microbiol. 2018, 9, 2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monogue, M.L.; Tsuji, M.; Yamano, Y.; Echols, R.; Nicolau, D.P. Efficacy of Humanized Exposures of Cefiderocol (S-649266) against a Diverse Population of Gram-Negative Bacteria in a Murine Thigh Infection Model. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum beta-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Chen, B.; Hulth, A.; Schwarz, S.; Ji, X.; Nilsson, L.E.; Ma, S.; Sun, Q.; Bi, Z.; Wang, Y.; et al. Genomic analysis of Staphylococcus aureus along a pork production chain and in the community, Shandong Province, China. Int. J. Antimicrob. Agents 2019, 54, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xu, X.; Zhang, L.; Xiong, Z.; Ma, Y.; Wei, Y.; Chen, Z.; Bai, J.; Liao, M.; Zhang, J. Fourth Generation Cephalosporin Resistance Among Salmonella enterica Serovar Enteritidis Isolates in Shanghai, China Conferred by bla CTX-M-55 Harboring Plasmids. Front. Microbiol. 2020, 11, 910. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Ke, B.; Deng, X.; Liang, J.; Ran, L.; Lu, L.; He, D.; Huang, Q.; Ke, C.; Li, Z.; et al. Serotypes, seasonal trends, and antibiotic resistance of non-typhoidal Salmonella from human patients in Guangdong Province, China, 2009-2012. BMC Infect. Dis. 2015, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Z.; Zhang, Y.; Zhang, Z.; Lei, L.; Xia, Z. Increasing Prevalence of ESBL-Producing Multidrug Resistance Escherichia coli From Diseased Pets in Beijing, China From 2012 to 2017. Front. Microbiol. 2019, 10, 2852. [Google Scholar] [CrossRef]
- Brown, A.C.; Chen, J.C.; Watkins, L.K.F.; Campbell, D.; Folster, J.P.; Tate, H.; Wasilenko, J.; Van Tubbergen, C.; Friedman, C.R. CTX-M-65 Extended-Spectrum beta-Lactamase-Producing Salmonella enterica Serotype Infantis, United States. Emerg. Infect. Dis. 2018, 24, 2284–2291. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Li, M.; Xu, X.; Fu, Y.; Xiong, Z.; Zhang, L.; Qu, X.; Zhang, H.; Wei, Y.; Zhan, Z.; et al. High-levels of resistance to quinolone and cephalosporin antibiotics in MDR-ACSSuT Salmonella enterica serovar Enteritidis mainly isolated from patients and foods in Shanghai, China. Int. J. Food Microbiol. 2018, 286, 190–196. [Google Scholar] [CrossRef]
- Qiao, J.; Zhang, Q.; Alali, W.Q.; Wang, J.; Meng, L.; Xiao, Y.; Yang, H.; Chen, S.; Cui, S.; Yang, B. Characterization of extended-spectrum beta-lactamases (ESBLs)-producing Salmonella in retail raw chicken carcasses. Int. J. Food Microbiol. 2017, 248, 72–81. [Google Scholar] [CrossRef]
- Usha, G.; Chunderika, M.; Prashini, M.; Willem, S.A.; Yusuf, E.S. Characterization of extended-spectrum beta-lactamases in Salmonella spp. at a tertiary hospital in Durban, South Africa. Diagn. Microbiol. Infect. Dis. 2008, 62, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008-2014. Emerg. Microbes Infect. 2018, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrois, D.; Breurec, S.; Seck, A.; Delaune, A.; Le Hello, S.; Pardos de la Gandara, M.; Sontag, L.; Perrier-Gros-Claude, J.D.; Sire, J.M.; Garin, B.; et al. Prevalence and characterization of extended-spectrum beta-lactamase-producing clinical Salmonella enterica isolates in Dakar, Senegal, from 1999 to 2009. Clin. Microbiol. Infect. 2014, 20, O109–O116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigemura, H.; Sakatsume, E.; Sekizuka, T.; Yokoyama, H.; Hamada, K.; Etoh, Y.; Carle, Y.; Mizumoto, S.; Hirai, S.; Matsui, M.; et al. Food Workers as a Reservoir of Extended-Spectrum-Cephalosporin-Resistant Salmonella Strains in Japan. Appl. Environ. Microbiol. 2020, 86, e00072-00020. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Z.L.; Huang, X.Y.; Ma, Z.B.; Guo, Z.W.; Lv, L.C.; Xia, Y.B.; Zeng, L.; Song, Q.H.; Liu, J.H. Evolution and Comparative Genomics of F33:A-:B- Plasmids Carrying blaCTX-M-55 or blaCTX-M-65 in Escherichia coli and Klebsiella pneumoniae Isolated from Animals, Food Products, and Humans in China. mSphere 2018, 3, e00137-00118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, J.; Wang, X.; Bai, X.; Ma, J.; Dang, R.; Xiong, Y.; Fanning, S.; Bai, L.; Yang, Z. Characterization of Five Escherichia coli Isolates Co-expressing ESBL and MCR-1 Resistance Mechanisms From Different Origins in China. Front. Microbiol. 2019, 10, 1994. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement; CLSI Document M100-S25; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Yang, B.; Wang, Q.; Cui, S.; Wang, Y.; Shi, C.; Xia, X.; Xi, M.; Wang, X.; Shi, X.; Wang, D.; et al. Characterization of extended-spectrum beta-lactamases-producing Salmonella strains isolated from retail foods in Shaanxi and Henan Province, China. Food Microbiol. 2014, 42, 14–18. [Google Scholar] [CrossRef]
- Archambault, M.; Petrov, P.; Hendriksen, R.S.; Asseva, G.; Bangtrakulnonth, A.; Hasman, H.; Aarestrup, F.M. Molecular characterization and occurrence of extended-spectrum beta-lactamase resistance genes among Salmonella enterica serovar Corvallis from Thailand, Bulgaria, and Denmark. Microb. Drug Resist. 2006, 12, 192–198. [Google Scholar] [CrossRef]
- Kiratisin, P.; Apisarnthanarak, A.; Laesripa, C.; Saifon, P. Molecular characterization and epidemiology of extended-spectrum-beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae isolates causing health care-associated infection in Thailand, where the CTX-M family is endemic. Antimicrob. Agents Chemother. 2008, 52, 2818–2824. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Alali, W.Q.; Liu, J.; Wang, Y.; Chen, S.; Cui, S.; Yang, B. Prevalence of Virulence Genes in Extended-Spectrum beta-lactamases (ESBLs)-Producing Salmonella in Retail Raw Chicken in China. J. Food Sci. 2018, 83, 1048–1052. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sheng, H.; Xu, W.; Huang, J.; Meng, L.; Cao, C.; Zeng, J.; Meng, J.; Yang, B. Diversity of Serotype, Genotype, and Antibiotic Susceptibility of Salmonella Prevalent in Pickled Ready-to-Eat Meat. Front. Microbiol. 2019, 10, 2577. [Google Scholar] [CrossRef]
- Wang, X.; Chen, G.; Wu, X.; Wang, L.; Cai, J.; Chan, E.W.; Chen, S.; Zhang, R. Increased prevalence of carbapenem resistant Enterobacteriaceae in hospital setting due to cross-species transmission of the bla NDM-1 element and clonal spread of progenitor resistant strains. Front. Microbiol. 2015, 6, 595. [Google Scholar] [CrossRef] [Green Version]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease ‘Burden of Illness Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Lupo, A.; Saras, E.; Madec, J.Y.; Haenni, M. Emergence of blaCTX-M-55 associated with fosA, rmtB and mcr gene variants in Escherichia coli from various animal species in France. J. Antimicrob. Chemother. 2018, 73, 867–872. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Jin, H.; Hu, J.; Yuan, Z.; Shi, W.; Yang, X.; Meng, J.; Xu, X. Laboratory monitoring of bacterial gastroenteric pathogens Salmonella and Shigella in Shanghai, China 2006–2012. Epidemiol. Infect. 2015, 143, 478–485. [Google Scholar] [CrossRef] [Green Version]
- China, National Health Commission. Antimicrobial Management and Bacterial Resistance in China. Available online: http://www.nhc.gov.cn/yzygj/s3594/201904/1b5a42f0e326487295b260c813da9b0e/files/c4328389c1b2462983fa94da9093cd05.pdf (accessed on 6 June 2020).
- Khosravani, M.; Soltan Dallal, M.M.; Norouzi, M. Phytochemical Composition and Anti-Efflux Pump Activity of Hydroalcoholic, Aqueous, and Hexane Extracts of Artemisia tournefortiana in Ciprofloxacin-Resistant Strains of Salmonella enterica Serotype Enteritidis. Iran. J. Public Health 2020, 49, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Usai, D.; Donadu, M.; Bua, A.; Molicotti, P.; Zanetti, S.; Piras, S.; Corona, P.; Ibba, R.; Carta, A. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J. Infect. Dev. Ctries 2019, 13, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, A.; Nakano, S.; Yamasaki, S.; Nishino, K. An efflux inhibitor of the MacAB pump in Salmonella enterica serovar Typhimurium. Microbiol. Immunol. 2020, 64, 182–188. [Google Scholar] [CrossRef]
- Donadu, M.G.; Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, M.; Marchetti, M.; Sanna, G.; Madeddu, S.; et al. Phytochemical Compositions and Biological Activities of Essential Oils from the Leaves, Rhizomes and Whole Plant of Hornstedtia bella Skornick. Antibiotics 2020, 9, 334. [Google Scholar] [CrossRef]
- Solarte, A.L.; Astorga, R.J.; de Aguiar, F.C.; De Frutos, C.; Barrero-Dominguez, B.; Huerta, B. Susceptibility Distribution to Essential Oils of Salmonella enterica Strains Involved in Animal and Public Health and Comparison of the Typhimurium and Enteritidis Serotypes. J. Med. Food 2018, 21, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, D.; Madeddu, S.; Marchetti, M.; Usai, M.; Rappelli, P.; et al. In vitro Antimicrobial Activity of Essential Oil Extracted from Leaves of Leoheo domatiophorus Chaowasku, D.T. Ngo and H.T. Le in Vietnam. Plants 2020, 9, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhao, L.; Hu, Y.; Dottorini, T.; Fanning, S.; Xu, J.; Li, F. Epidemiological Study on Prevalence, Serovar Diversity, Multidrug Resistance, and CTX-M-Type Extended-Spectrum beta-Lactamases of Salmonella spp. from Patients with Diarrhea, Food of Animal Origin, and Pets in Several Provinces of China. Antimicrob. Agents Chemother. 2020, 64, e00092-00020. [Google Scholar] [CrossRef] [PubMed]
- Nadimpalli, M.; Fabre, L.; Yith, V.; Sem, N.; Gouali, M.; Delarocque-Astagneau, E.; Sreng, N.; Le Hello, S.; The BIRDY Study Group. CTX-M-55-type ESBL-producing Salmonella enterica are emerging among retail meats in Phnom Penh, Cambodia. J. Antimicrob. Chemother. 2019, 74, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Chen, Q.; Yu, X.; Li, Q.; Ding, B.; Yang, L.; Chen, C.; Qin, Z.; Parsons, C.; Zhang, X.; et al. High prevalence of extended-spectrum beta lactamases among Salmonella enterica Typhimurium isolates from pediatric patients with diarrhea in China. PLoS ONE 2011, 6, e16801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Chang, Y.J.; Fang, S.H.; Su, L.H.; Li, H.C.; Yang, H.P.; Yu, M.J.; Chiu, C.H. Emergence and Evolution of High-Level Cephalosporin-Resistant Salmonella Goldcoast in Northern Taiwan. Open Forum. Infect. Dis. 2019, 6, ofz447. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Ding, X.M.; Lin, X.L.; Sun, R.Y.; Lu, Y.W.; Cai, R.M.; Webber, M.A.; Ding, H.Z.; Jiang, H.X. The Emergence of Chromosomally Located bla CTX-M-55 in Salmonella from Foodborne Animals in China. Front. Microbiol. 2019, 10, 1268. [Google Scholar] [CrossRef]
- Adator, E.H.; Walker, M.; Narvaez-Bravo, C.; Zaheer, R.; Goji, N.; Cook, S.R.; Tymensen, L.; Hannon, S.J.; Church, D.; Booker, C.W.; et al. Whole Genome Sequencing Differentiates Presumptive Extended Spectrum Beta-Lactamase Producing Escherichia coli along Segments of the One Health Continuum. Microorganisms 2020, 8, 448. [Google Scholar] [CrossRef] [Green Version]
- Sanjit Singh, A.; Lekshmi, M.; Prakasan, S.; Nayak, B.B.; Kumar, S. Multiple Antibiotic-Resistant, Extended Spectrum-beta-Lactamase (ESBL)-Producing Enterobacteria in Fresh Seafood. Microorganisms 2017, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, I.; Magnet, S.; Hawser, S.; Shapiro, S.; Knechtle, P. In Vitro Activity of Cefepime-Enmetazobactam against Gram-Negative Isolates Collected from U.S. and European Hospitals during 2014-2015. Antimicrob. Agents Chemother. 2019, 63, e00514-00519. [Google Scholar] [CrossRef] [Green Version]
- Luk-In, S.; Chatsuwan, T.; Pulsrikarn, C.; Bangtrakulnonth, A.; Rirerm, U.; Kulwichit, W. High prevalence of ceftriaxone resistance among invasive Salmonella enterica serotype Choleraesuis isolates in Thailand: The emergence and increase of CTX-M-55 in ciprofloxacin-resistant S. Choleraesuis isolates. Int. J. Med. Microbiol. 2018, 308, 447–453. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. (%) of ESBL-SE Isolates (n = 233) | No. (%) of Non-ESBL-SE Isolates (n = 59) | Unadjusted OR (95% CI) 1 |
|---|---|---|---|
| Year | |||
| 2006–2010 | 6 (2.6) | 22 (37.3) | 1.0 (ref) |
| 2011 | 19 (8.2) | 21 (35.6) | 3.3 (1.1–9.9) |
| 2012 | 46 (19.7) | 2 (3.4) | 84.3 (15.7–452.1) |
| 2013 | 131 (56.2) | 8 (13.6) | 60.0 (19.0–189.8) |
| 2014 | 31 (13.3) | 6 (10.2) | 18.9 (5.4–66.6) |
| Source | |||
| General outpatient | 33 (14.2) | 4 (6.8) | 1.0 (ref) |
| Intestinal outpatient | 132 (56.7) | 47 (79.7) | 0.3 (0.1–1.0) |
| Other outpatient | 45 (19.3) | 8 (13.6) | 0.7 (0.2–2.5) |
| Others | 23 (9.9) | 0 (0.0) | |
| District | |||
| Minhang district | 109 (46.8) | 22 (37.3) | 1.0 (ref) |
| Jing’an district | 53 (22.7) | 5 (8.5) | 2.1 (0.8–6.0) |
| Jinshan district | 14 (6.0) | 7 (11.9) | 0.4 (0.2–1.1) |
| Huangpu district | 8 (3.4) | 10 (16.9) | 0.2 (0.06–0.5) |
| Changning district | 9 (3.9) | 8 (13.6) | 0.2 (0.08–0.7) |
| Others | 40 (17.2) | 7 (11.9) | 1.2 (0.5–2.9) |
| Hospital | |||
| Public hospital | 79 (33.9) | 33 (55.9) | 1.0 (ref) |
| Pediatric hospital | 126 (54.1) | 25 (42.4) | 2.1 (1.2–3.9) |
| Community hospital | 14 (6.0) | 1 (1.7) | 5.9 (0.8–46.9) |
| Others | 14 (6.0) | 0 (0.0) | |
| ESBL-Encoding Gene | No. (%) of Isolates by Year | Total No. (%) (n = 233) | ||||
|---|---|---|---|---|---|---|
| 2006–2010 (n = 6) | 2011 (n = 19) | 2012 (n = 46) | 2013 (n = 131) | 2014 (n = 31) | ||
| blaCTX-M | ||||||
| blaCTX-M-3 | 0 (0.0) | 1 (5.3) | 2 (4.3) | 0 (0.0) | 0 (0.0) | 3 (1.3) |
| blaCTX-M-15 | 0 (0.0) | 0 (0.0) | 1 (2.2) | 0 (0.0) | 0 (0.0) | 1 (0.4) |
| blaCTX-M-55 | 0 (0.0) | 13 (68.4) | 28 (60.9) | 90 (68.7) | 21 (67.7) | 152 (65.2) |
| blaCTX-M-64 | 0 (0.0) | 0 (0.0) | 4 (8.7) | 0 (0.0) | 0 (0.0) | 4 (1.7) |
| blaCTX-M-79 | 0 (0.0) | 0 (0.0) | 1 (2.2) | 1 (0.8) | 0 (0.0) | 2 (0.9) |
| blaCTX-M-123 | 1 (16.7) | 1 (5.3) | 1 (2.2) | 0 (0.0) | 1 (3.2) | 4 (1.7) |
| Total | 1 (16.7) | 15 (78.9) | 37 (80.4) | 91 (69.5) | 22 (71.0) | 166 (71.2) |
| blaTEM | ||||||
| blaTEM-1 | 1 (16.7) | 12 (63.2) | 26 (56.5) | 14 (10.7) | 3 (9.7) | 56 (24.0) |
| blaTEM-214 | 0 (0.0) | 3 (15.8) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 3 (1.3) |
| Total | 1 (16.7) | 15 (78.9) | 26 (56.5) | 14 (10.7) | 3 (9.7) | 59 (25.3) |
| Cephalosporin | blaCTX-M-55-Positive ESBL-SE (n = 152) | blaCTX-M-55-Negative ESBL-SE (n = 81) | p Value 1 | ||||
|---|---|---|---|---|---|---|---|
| MIC50 (μg/mL) | MIC90 (μg/mL) | Resistance (%) | MIC50 (μg/mL) | MIC90 (μg/mL) | Resistance (%) | ||
| Cefotaxime | 64 | 64 | 100.0 | 64 | 64 | 100.0 | - |
| Ceftazidime | 16 | 32 | 91.5 | 16 | 32 | 67.9 | 0.0 ** |
| Ceftriaxone | 128 | 128 | 98.0 | 128 | 128 | 92.6 | 0.04 * |
| Cefepime | 16 | 16 | 75.0 | 16 | 16 | 58.0 | 0.008 ** |
| Cefazolin | 16 | 16 | 98.0 | 16 | 16 | 92.6 | 0.04 * |
| Cefpodoxime | 32 | 32 | 98.0 | 32 | 32 | 92.6 | 0.04 * |
| Cefoxitin | 4 | 4 | 0.7 | 4 | 8 | 2.5 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, C.; Niu, Q.; Chen, J.; Xu, X.; Sheng, H.; Cui, S.; Liu, B.; Yang, B. Epidemiology and Characterization of CTX-M-55-Type Extended-Spectrum β-Lactamase-Producing Salmonella enterica Serovar Enteritidis Isolated from Patients in Shanghai, China. Microorganisms 2021, 9, 260. https://doi.org/10.3390/microorganisms9020260
Cao C, Niu Q, Chen J, Xu X, Sheng H, Cui S, Liu B, Yang B. Epidemiology and Characterization of CTX-M-55-Type Extended-Spectrum β-Lactamase-Producing Salmonella enterica Serovar Enteritidis Isolated from Patients in Shanghai, China. Microorganisms. 2021; 9(2):260. https://doi.org/10.3390/microorganisms9020260
Chicago/Turabian StyleCao, Chenyang, Qinya Niu, Jia Chen, Xuebin Xu, Huanjing Sheng, Shenghui Cui, Bin Liu, and Baowei Yang. 2021. "Epidemiology and Characterization of CTX-M-55-Type Extended-Spectrum β-Lactamase-Producing Salmonella enterica Serovar Enteritidis Isolated from Patients in Shanghai, China" Microorganisms 9, no. 2: 260. https://doi.org/10.3390/microorganisms9020260