
Effect of 17β-Estradiol, Progesterone, and Tamoxifen on Neurons Infected with Toxoplasma gondii In Vitro

 ,
, 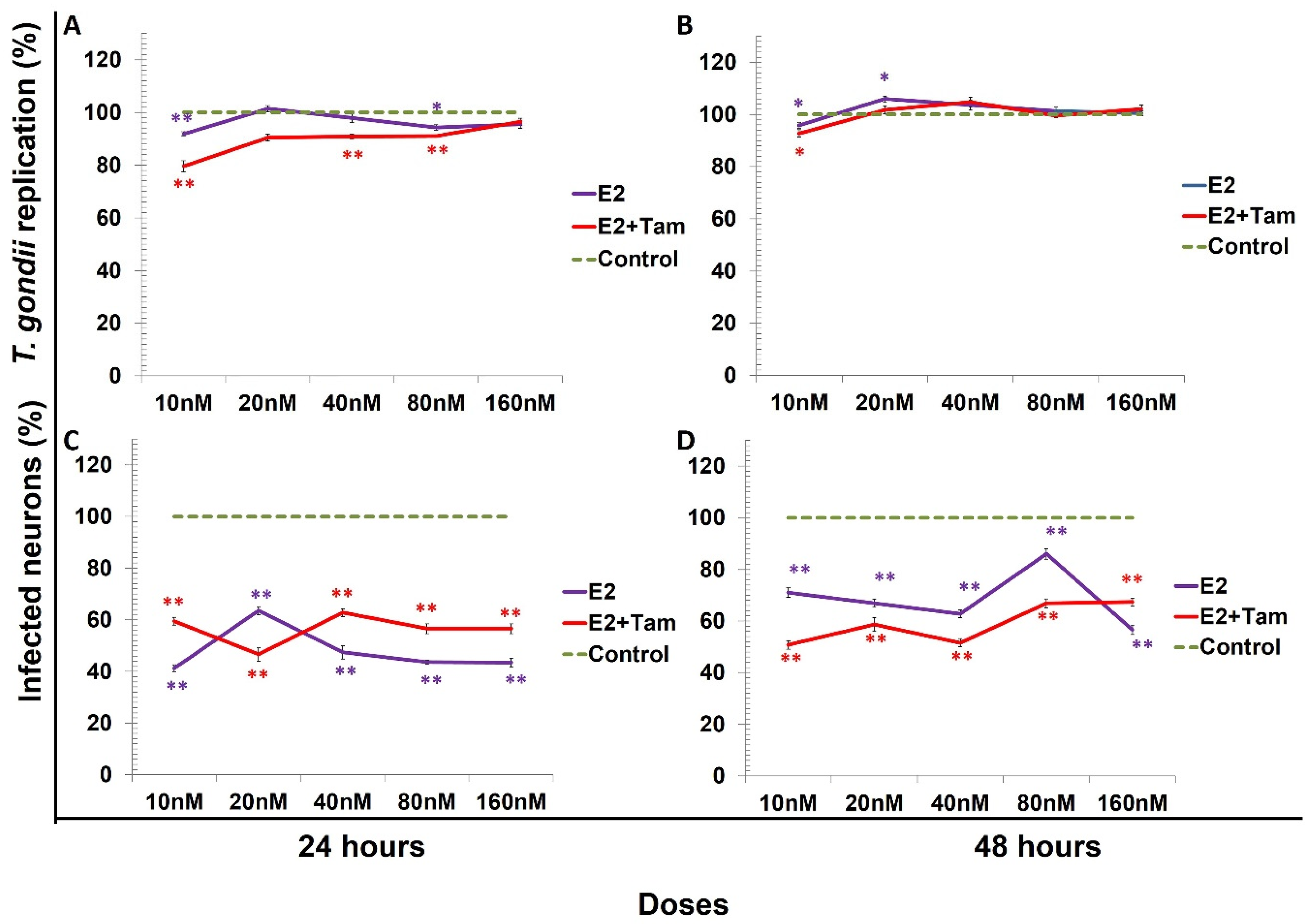{kind=link}
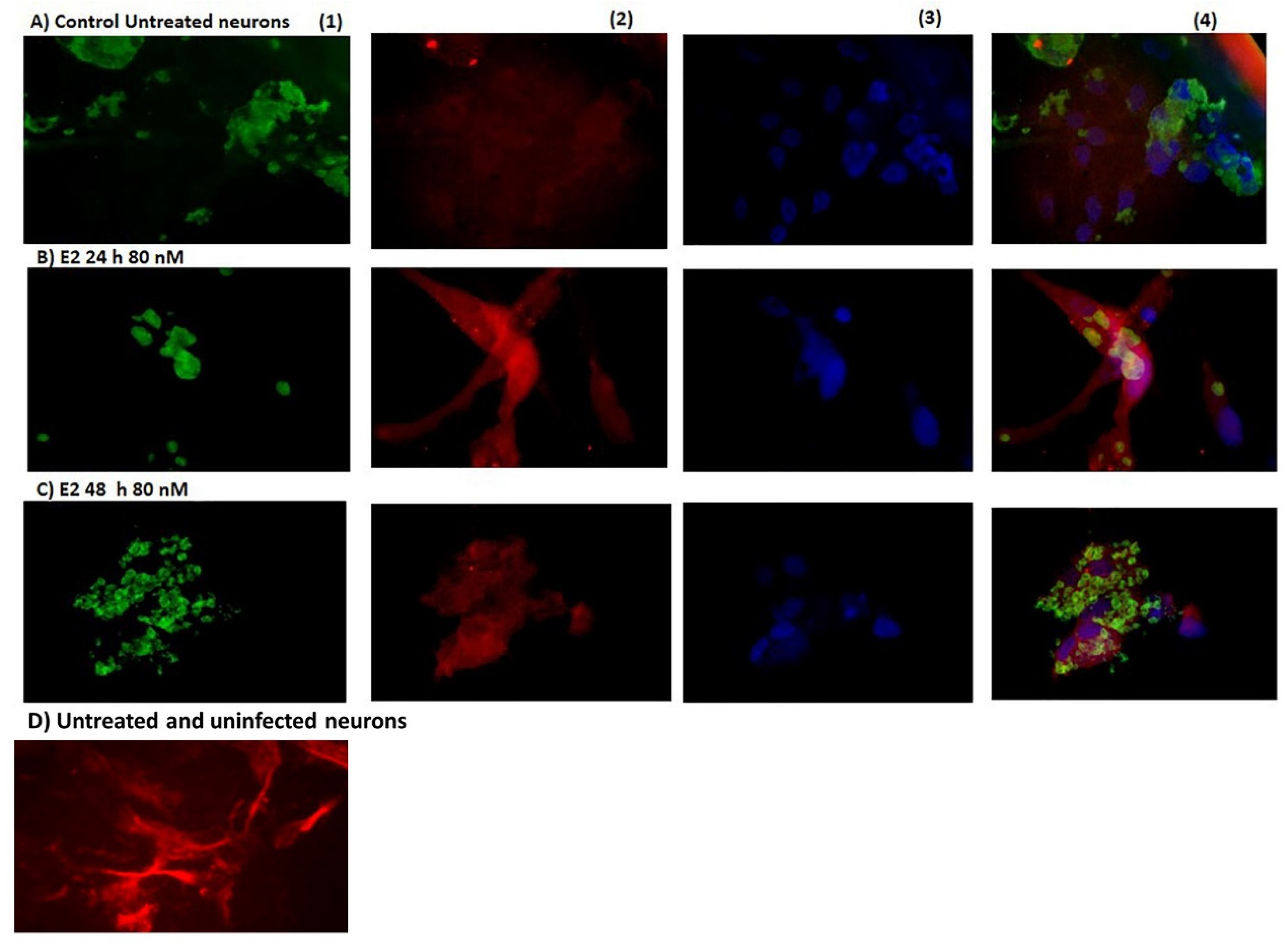{kind=link}
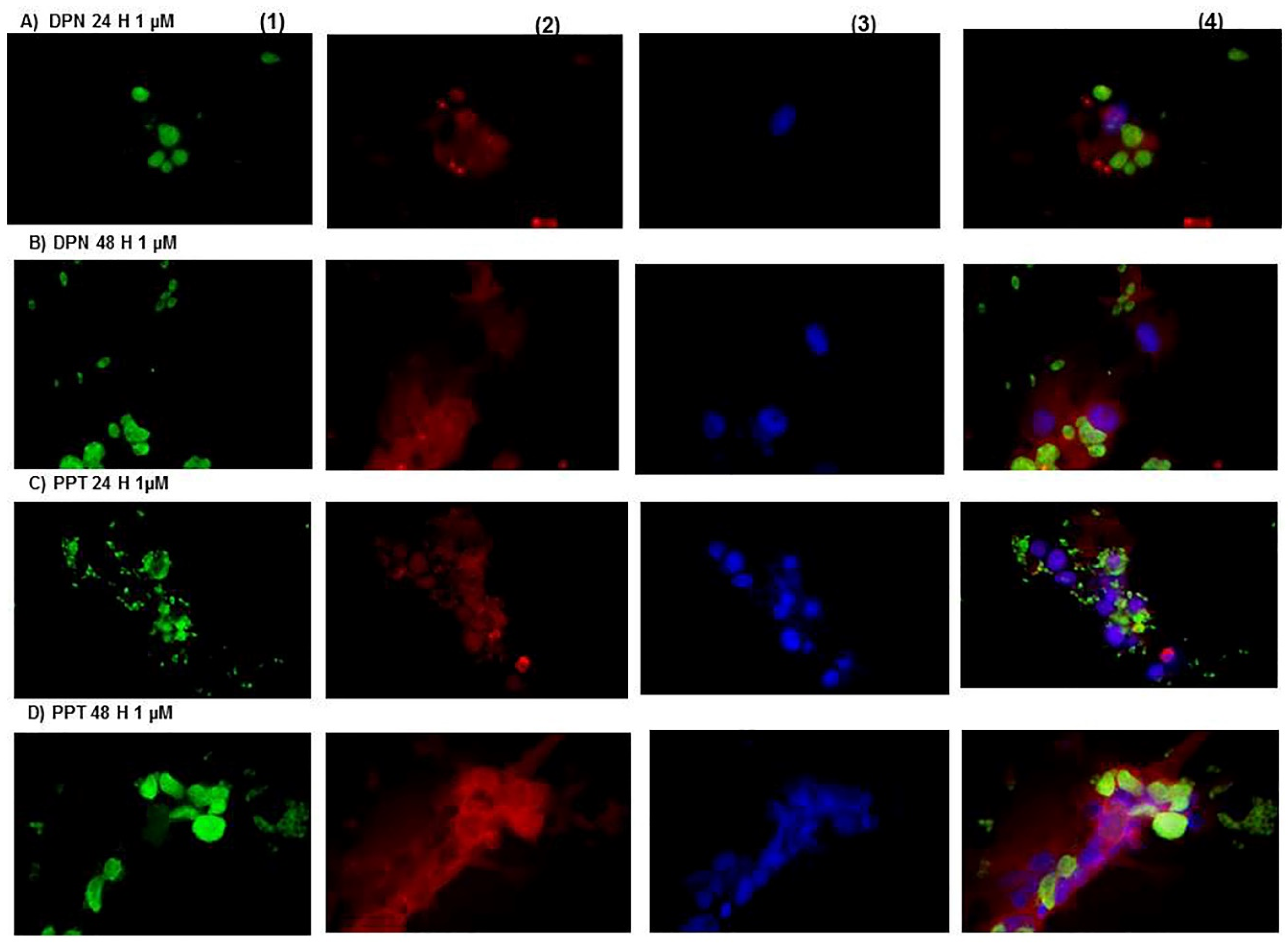{kind=link}
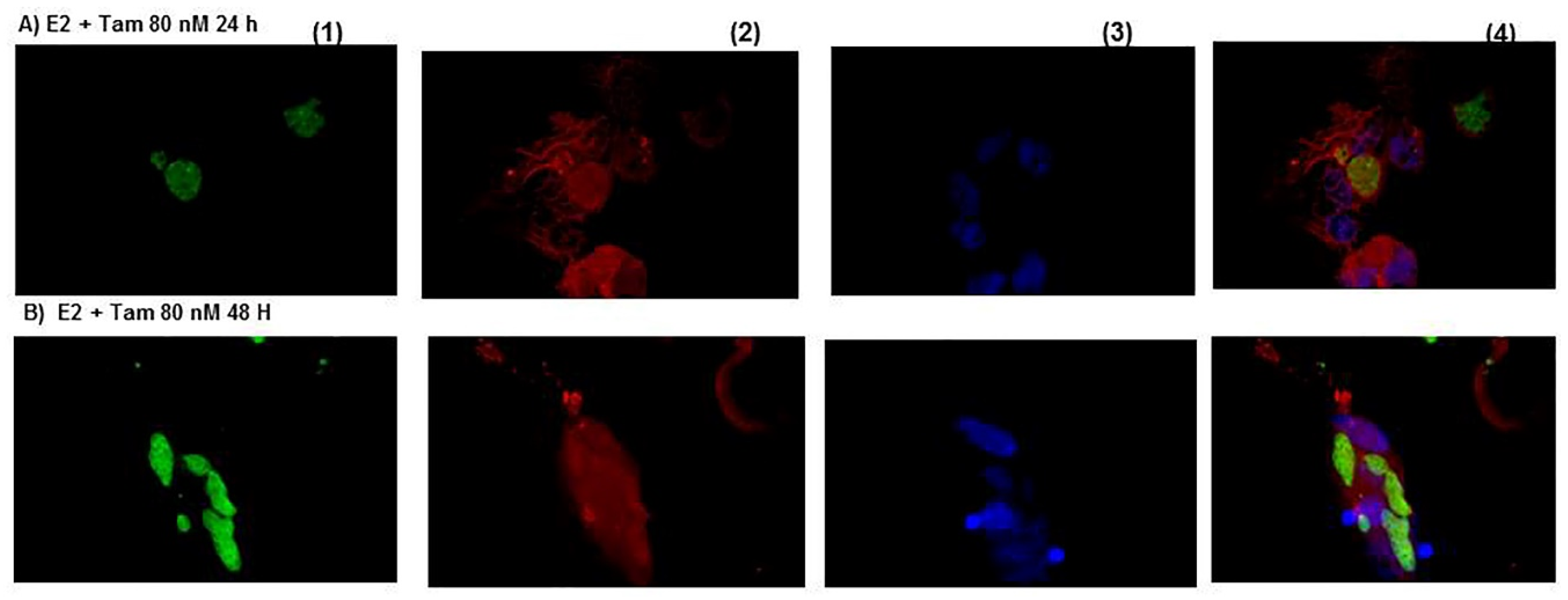{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethical Aspects
2.2. Parasites
2.3. Cortical Neuron Culture
2.4. Neuron Separation
2.5. Experimental Treatments in Neurons In Vitro
2.6. Immunocytochemical Method
2.7. Microscopic Analysis
2.8. T. gondii Replication
2.9. Cytotoxicity Assay
2.10. Statistical Analysis
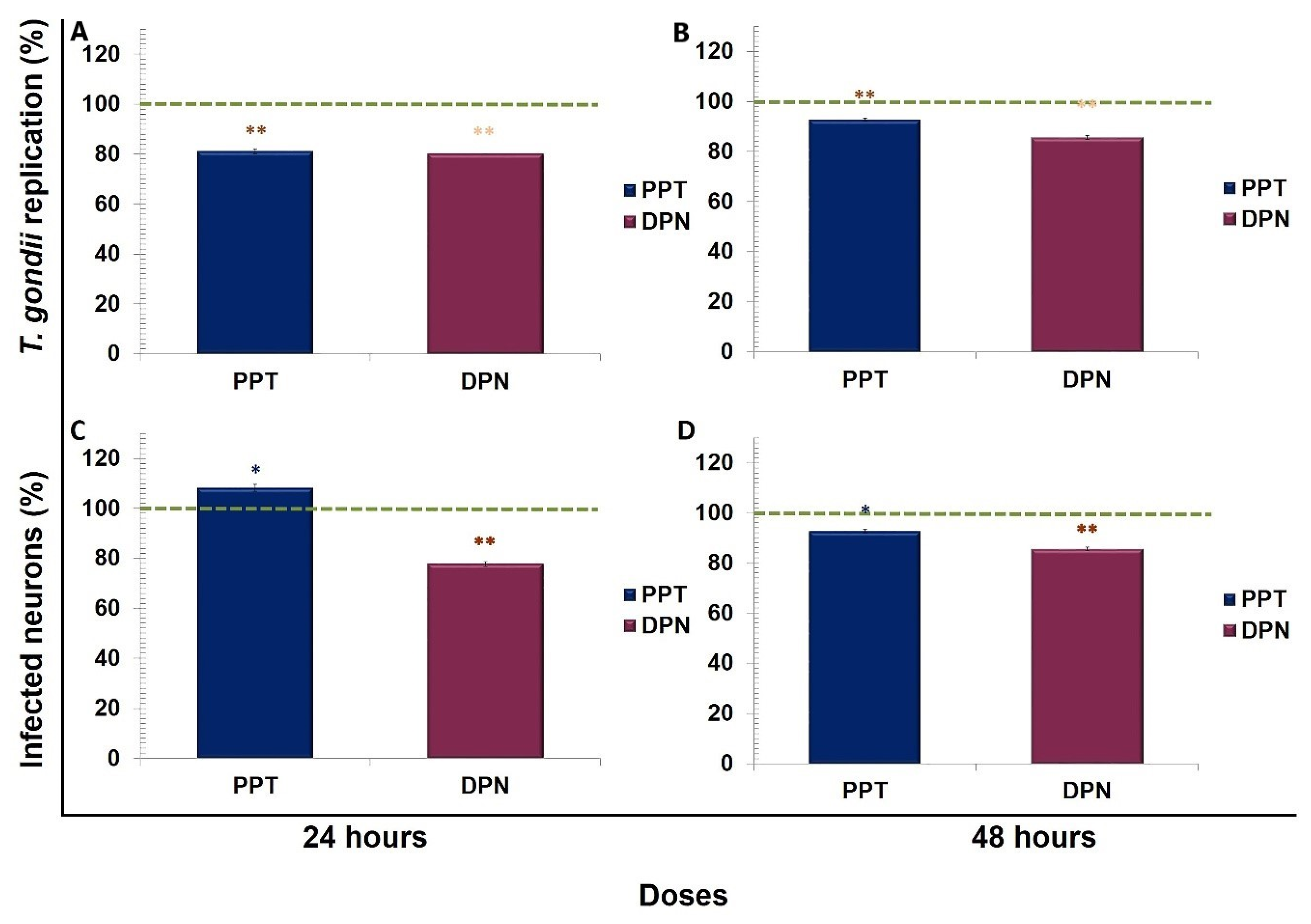
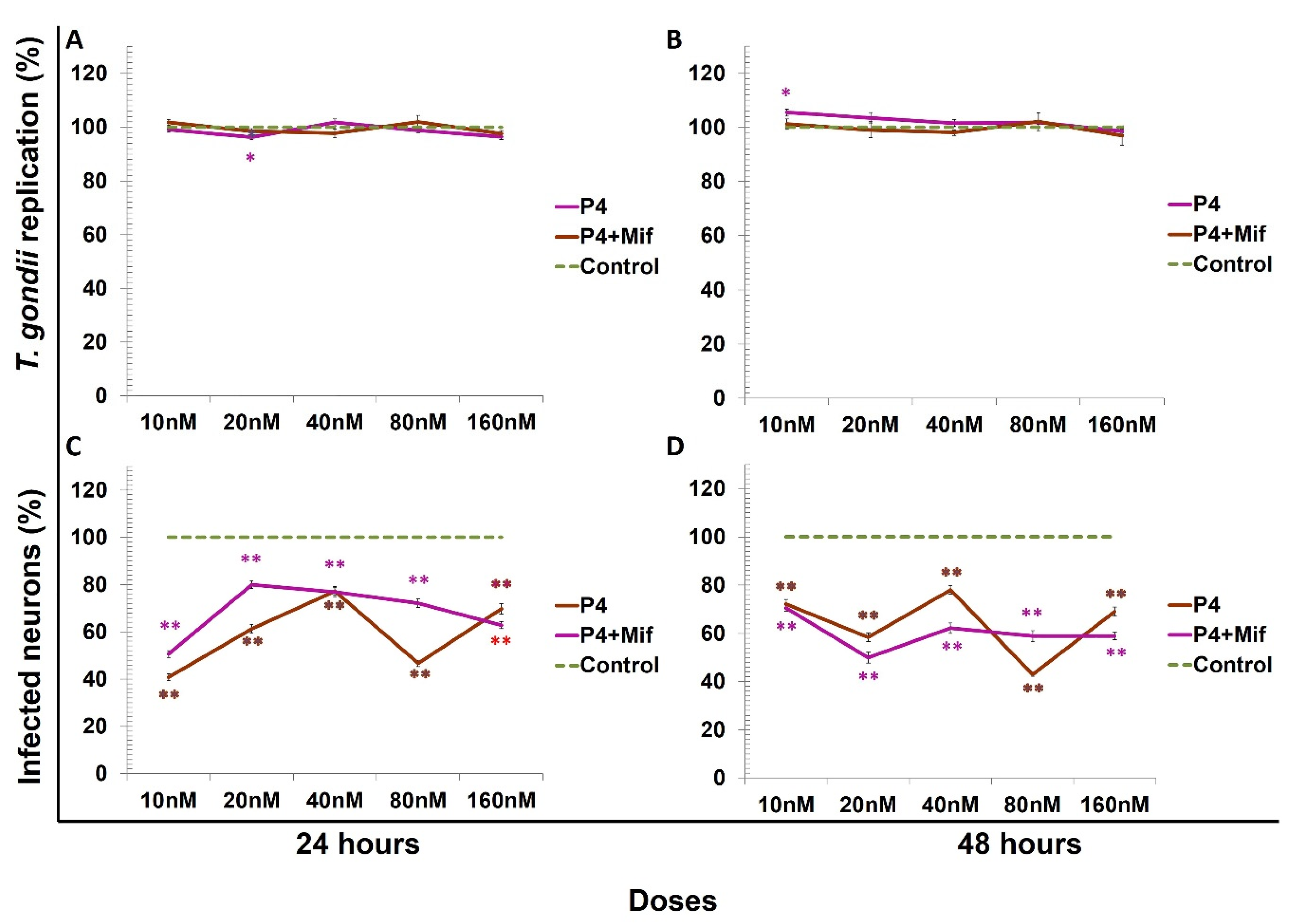
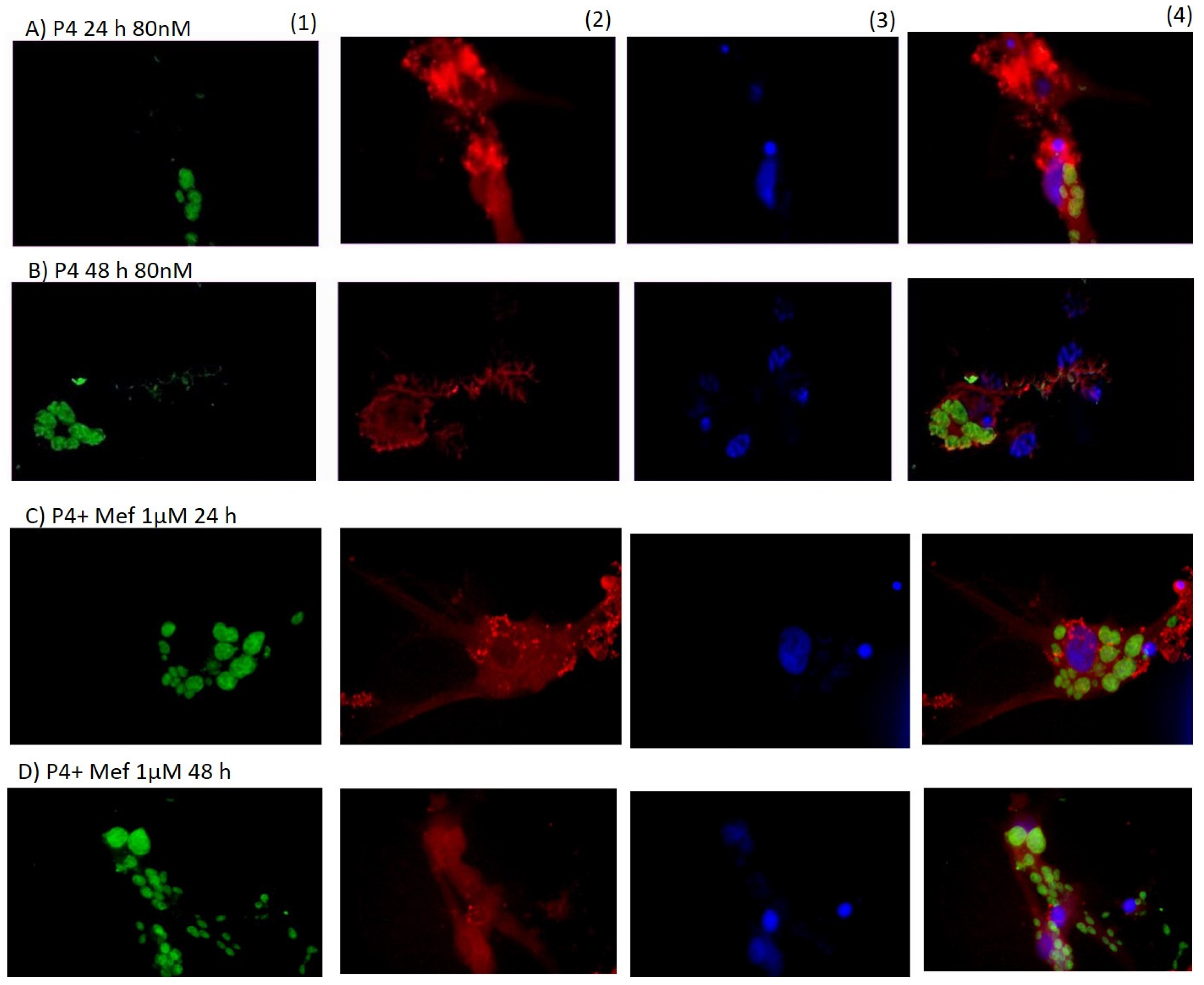
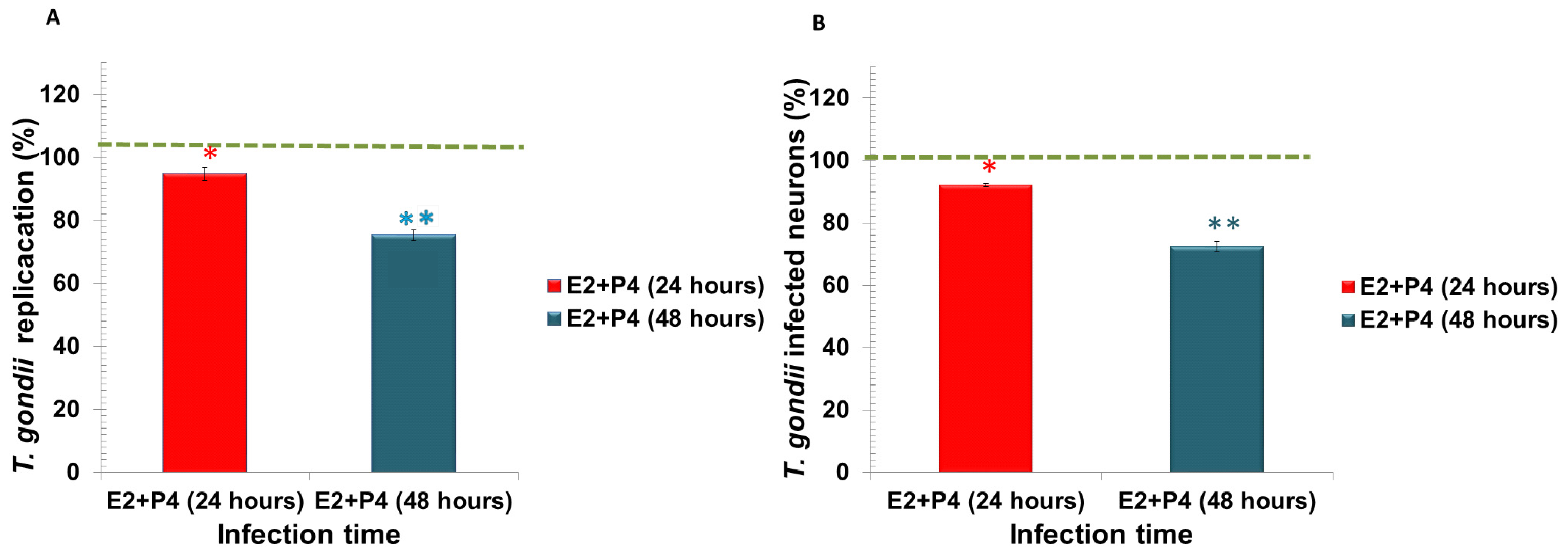
3. Results
Step and Displacement Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barragan, A.; Hitziger, N. Transepithelial Migration by Toxoplasma. Subcell Biochem. 2008, 47, 198–207. [Google Scholar] [CrossRef]
- Cabral, C.M.; Tuladhar, S.; Dietrich, H.K.; Nguyen, E.; Macdonald, W.R.; Trivedi, T.; Devineni, A.; Koshy, A.A. Neurons are the Primary Target Cell for the Brain-Tropic Intracellular Parasite Toxoplasma gondii. PLoS Pathog. 2016, 12, e1005447. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.; Hutchison, W.M. The host-parasite relationship of Toxoplasma gondii in the brains of chronically infected mice. Virchows Arch. A Pathol. Anat Histopathol. 1987, 411, 39–43. [Google Scholar] [CrossRef]
- Suzuki, T.; Schirra, F.; Richards, S.M.; Jensen, R.V.; Sullivan, D.A. Estrogen and Progesterone Control of Gene Expression in the Mouse Meibomian Gland. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1797–1808. [Google Scholar] [CrossRef]
- Parlog, A.; Schlüter, D.; Dunay, I.R. Toxoplasma gondii-induced neuronal alterations. Parasite Immunol. 2015, 37, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Tanakaa, N.; Ashoura, D.; Dratzb, E.; Halonen, S. Use of human induced pluripotent stem cell-derived neurons as a model for Cerebral Toxoplasmosis. Microbes Infect. 2015, 18, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Creuzet, C.; Robert, F.; Roisin, M.P.; Van Tan, H.; Benes, C.; Dupouy-Camet, J.; Fagard, R. Neurons in primary culture are less efficiently infected by Toxoplasma gondii than glial cells. Parasitol. Res. 1998, 84, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Marin, J.E. Manifestaciones clínicas de la toxoplasmosis humana. In Toxoplasmosis Humna; Galván-Ramírez, M., Flores, M., Eds.; Ecorfan: Ecatepec, Mexico, 2017; pp. 139–162. [Google Scholar]
- Galván-Ramírez, M.d.l.L.; Navarro Machuca, G.; Covarrubias Castillo, S.A.; Benavides González, J.C.; Rodríguez Pérez, L.R.; Dueñas Jiménez, S.H.; Dueñas Jiménez, J.M. Toxoplasmosis Is More Frequent in Schizophrenia Patients Than in the General Population in Mexico and Is Not Associated with More Severe Course of Schizophrenia Measured with the Brief Psychiatric Rating Scale. Pathogens 2021, 10, 820. [Google Scholar] [CrossRef]
- Roberts, C.W.; Cruickshank, S.M.; Alexander, J. Sex-determined resistance to Toxoplasma gondii is associated with temporal differences in cytokine production. Infect. Immun. 1995, 63, 2549–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, W.; Roberts, C.W.; Ferguson, D.J.; Jebbari, H.; Alexander, J. Innate immunity to Toxoplasma gondii is influenced by gender and is associated with differences in interleukin-12 and gamma interferon production. Infect. Immun. 1997, 65, 1119–1121. [Google Scholar] [CrossRef] [Green Version]
- Liesenfeld, O.; Nguyen, T.A.; Pharke, C.; Suzuki, Y. Importance of gender and sex hormones in regulation of susceptibility of the small intestine to peroral infection with Toxoplasma gondii tissue cysts. J. Parasitol. 2001, 87, 1491–1493. [Google Scholar] [CrossRef]
- Muñiz-Hernández, S.; Luna-Nophal, A.; León, C.T.G.; Domínguez-Ramírez, L.; Patrón-Soberano, O.A.; Nava-Castro, K.E.; Ostoa-Saloma, P.; Morales-Montor, J. Dehydroepiandrosterone Effect on Toxoplasma gondii: Molecular Mechanisms Associated to Parasite Death. Microorganisms 2021, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Segura, L.M.; Melcangi, R.C. Steroids and glial cell function. Glia 2006, 54, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Shao, R.; Cao, S.; Wang, X.; Feng, Y.; Billig, H. The elusive and controversial roles of estrogen and progesterone receptors in human endometriosis. Am. J. Transl. Res. 2014, 6, 104–113. [Google Scholar] [PubMed]
- Sa, Q.; Ochiai, E.; Tiwari, A.; Perkins, S.; Mullins, J.; Gehman, M.; Huckle, W.R.; Eyestone, W.H.; Saunders, T.L.; Shelton, B.J.; et al. Cutting Edge: IFN-γ Produced by Brain-Resident Cells Is Crucial to Control Cerebral Infection with Toxoplasma gondii. J. Immunol. 2015, 195, 796–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinjak, Z.; Bilalovic, N. Estrogen and Progesterone Receptors in Endometrium in Women with Unexplained Infertility. Mater. Socio Medica 2014, 26, 51–52. [Google Scholar] [CrossRef] [Green Version]
- Verthelyi, D. Sex hormones as immunomodulators in health and disease. Int. Immunopharmacol. 2001, 1, 983–993. [Google Scholar] [CrossRef]
- McEwen, B.S.; Akama, K.T.; Spencer-Segal, J.L.; Milner, T.A.; Waters, E.M. Estrogen effects on the brain: Actions beyond the hypothalamus via novel mechanisms. Behav. Neurosci. 2012, 126, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Roque, C.; Baltazar, G. G protein-coupled estrogen receptor 1 (GPER) activation triggers different signaling pathways on neurons and astrocytes. Neural Regen. Res. 2019, 14, 2069–2070. [Google Scholar] [CrossRef]
- Garrido-Gracia, J.C.; Gordon, A.; Bellido, C.; Aguilar, R.; Barranco, I.; Millán, Y.; de Las Mulas, J.M.; Sánchez-Criado, J.E. The integrated action of estrogen receptor isoforms and sites with progesterone receptor in the gonadotrope modulates LH secretion: Evidence from tamoxifen-treated ovariectomized rats. J. Endocrinol. 2007, 193, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Gottfried-Blackmore, A.; Croft, G.; McEwen, B.S.; Bulloch, K. Transcriptional activity of estrogen receptors ERalpha and ERbeta in the EtC.1 cerebellar granule cell line. Brain Res. 2007, 1186, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Harrington, W.R.; Sheng, S.; Barnett, D.H.; Petz, L.N.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Activities of estrogen receptor alpha- and beta-selective ligands at diverse estrogen responsive gene sites mediating transactivation or transrepression. Mol. Cell. Endocrinol. 2003, 206, 13–22. [Google Scholar] [CrossRef]
- Franco-Rodríguez, N.E.; Dueñas-Jiménez, J.M.; De la Torre Valdovinos, B.; López Ruiz, J.R.; Hernández Hernández, L.; Dueñas Jiménez, S.H. Tamoxifen favoured the rat sensorial cortex regeneration after a penetrating brain injury. Brain Res. Bull. 2013, 98, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-S.Y.; Yin, Z.; Milatovic, D.; Jiang, H.; Aschner, M. Estrogen and Tamoxifen Protect against Mn-Induced Toxicity in Rat Cortical Primary Cultures of Neurons and Astrocytes. Toxicol. Sci. 2009, 110, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Dávila, J.T.; Garrán, A.D.T. Selective estrogen receptors modulators (SERMs): Biochemistry, pharmacology, and clinical use in gynecology. Ginecol. Obstet. Méx. 2005, 73, 424–435. [Google Scholar]
- Melcangi, R.C.; Giatti, S.; Calabrese, D.; Pesaresi, M.; Cermenati, G.; Mitro, N.; Viviani, B.; Garcia-Segura, L.M.; Caruso, D. Levels and actions of progesterone and its metabolites in the nervous system during physiological and pathological conditions. Prog. Neurobiol. 2014, 113, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Diep, C.H.; Daniel, A.R.; Mauro, L.J.; Knutson, T.P.; Lange, C.A. Progesterone action in breast, uterine, and ovarian cancers. J. Mol. Endocrinol. 2015, 54, R31–R53. [Google Scholar] [CrossRef] [Green Version]
- Cadepond, F.; Ulmann, A.; Baulieu, E.E. RU486 (MIFEPRISTONE): Mechanisms of Action and Clinical Uses. Annu. Rev. Med. 1997, 48, 129–156. [Google Scholar] [CrossRef] [Green Version]
- Behl, C.; Trapp, T.; Skutella, T.; Holsboer, F. Protection against oxidative stress-induced neuronal cell death—A novel role for RU486. Eur. J. Neurosci. 1997, 9, 912–920. [Google Scholar] [CrossRef]
- Khan, J.A.; Tikad, A.; Fay, M.; Hamze, A.; Fagart, J.; Chabbert-Buffet, N.; Meduri, G.; Amazit, L.; Brion, J.-D.; Alami, M.; et al. A New Strategy for Selective Targeting of Progesterone Receptor with Passive Antagonists. Mol. Endocrinol. 2013, 27, 909–924. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, P.; Chabbert-Buffet, N.; Fauser, B.C. Selective progesterone receptor modulators in reproductive medicine: Pharmacology, clinical efficacy and safety. Fertil. Steril. 2011, 96, 1175–1189. [Google Scholar] [CrossRef]
- Choi, S.; Shin, H.; Song, H.; Lim, H.J. Suppression of autophagic activation in the mouse uterus by estrogen and progesterone. J. Endocrinol. 2014, 221, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Spitz, I.M. Progesterone receptor antagonists. Curr. Opin. Investig. Drugs 2016, 7, 882–890. [Google Scholar]
- Chabbert-Buffet, N.; Meduri, G.; Bouchard, P.; Spitz, I.M. Selective progesterone receptor modulators and progesterone antagonists: Mechanisms of action and clinical applications. Hum. Reprod. Updat. 2005, 11, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pung, O.J.; Luster, M.I. Toxoplasma gondii: Decreased resistance to infection in mice due to estrogen. Exp. Parasitol. 1986, 61, 48–56. [Google Scholar] [CrossRef]
- Jones, L.A.; Jean-Paul, A.; Henriquez, F.L.; Lyons, R.E.; Nickdel, M.B.; Carter, K.C. Toll-like receptor-4-mediated macrophage activation is differentially regulated by progesterone via the glucocorticoid and progesterone receptors. Immunology 2008, 125, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Galván-Ramírez, M.d.l.L.; Gutiérrez-Maldonado, A.F.; Verduzco-Grijalva, F.; Jiménez, J.M.D. The role of hormones on Toxoplasma gondii infection: A systematic review. Front. Microbiol. 2014, 5, 503. [Google Scholar] [CrossRef]
- Galván-Ramírez, M.d.l.L.; De Arellano, A.R.; Rodríguez-Pérez, L.R.; Lopez-Pulido, E.I.; Muñoz-Valle, J.F.; Pereira-Suárez, A.L. Hormonal modulation of Toxoplasma gondii infection: Regulation of hormonal receptors and cytokine production in THP-1 cells. Exp. Parasitol. 2019, 204, 107721. [Google Scholar] [CrossRef]
- Galván-Ramírez, M.d.l.L.; Jiménez, J.M.D.; Pérez, L.R.R.; Troyo-Sanroman, R.; Ramírez-Herrera, M.A.; García-Iglesias, T. Effect of Nitaxozanide and Pyrimethamine on Astrocytes Infected by Toxoplasma gondii In Vitro. Arch. Med. Res. 2013, 44, 415–421. [Google Scholar] [CrossRef]
- Mossman, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Anand, N.; Kanwar, K.R.; Dubey, M.L.; Vahishta, R.K.; Sehgal, R.; Verma, A.K. Effect of lactoferrin protein on red blood cells and macrophages: Mechanism of parasite–host interaction. Drug Des. Dev. Ther. 2015, 9, 3821–3835. [Google Scholar] [CrossRef] [Green Version]
- Dzitko, K.; Gatkowska, J.; Płociński, P.; Dziadek, B.; Długońska, H. The effect of prolactin (PRL) on the growth of Toxoplasma gondii tachyzoites in vitro. Parasitol. Res. 2010, 107, 199–204. [Google Scholar] [CrossRef]
- Courret, N.; Darche, S.; Sonigo, P.; Milon, G.; Buzoni-Gâtel, D.; Tardieux, I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006, 107, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Ochoa, C.O.; Lagunas-Martínez, A.; Belkind-Gerson, J.; Díaz-Chávez, J.; Correa, D. Toxoplasma gondii invasion and replication within neonate mouse astrocytes and changes in apoptosis related molecules. Exp. Parasitol. 2013, 134, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Teppa, G.A.D.; Terán, D.J. New biochemical and molecular aspects of estrogen’s action. Ginecol. Obstet. Méx. 2005, 73, 436–442. [Google Scholar]
- Halonen, S.K.; Lyman, W.D.; Chiu, F.C. Growth and Development of Toxoplasma gondii Human Neurons and Astrocytes. J. Neuropathol. Exp. Neurol. 1996, 55, 1150–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galván-Ramírez, M.L.; Gutiérrez-Maldonado, A.F.; Dueñas-Jiménez, J.M.; Rodríguez-Pérez, L.R.; Troyo-Sanromán, R. Effect of 17β-estradiol and progesterone on astrocytes infected with Toxoplasma. J. Adv. Parasitol. 2016, 3, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Dittmar, A.J.; Drozda, A.A.; Blader, I.J. Drug Repurposing Screening Identifies Novel Compounds That Effectively Inhibit Toxoplasma gondii Growth. mSphere 2016, 1, e00042-15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, J.; Li, M.; Fu, Y.; Zhang, T.; Han, Q.; Liu, Q. Role of an estradiol regulatory factor-hydroxysteroid dehydrogenase (HSD) in Toxoplasma gondii infection and pathogenicity. J. Steroid Biochem. Mol. Biol. 2017, 174, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Segura, L.M.; Azcoitia, I.; DonCarlos, L.L. Neuroprotection by estradiol. Prog. Neurobiol. 2001, 63, 29–60. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Segura, L.M.; Dueñas, M.; Busiguina, S.; Naftolin, F.; Chowen, J.A. Gonadal hormone regulation of neuronal-glial interactions in the developing neuroendocrine hypothalamus. J. Steroid Biochem. Mol. Biol. 1995, 53, 293–298. [Google Scholar] [CrossRef]
- Garcia-Segura, L.M.; Sanz, A.; Mendez, P. Cross-Talk between IGF-I and Estradiol in the Brain: Focus on Neuroprotection. Neuroendocrinology 2006, 84, 275–279. [Google Scholar] [CrossRef]
- Spampinato, S.F.; Molinaro, G.; Merlo, S.; Iacovelli, L.; Caraci, F.; Battaglia, G.; Nicoletti, F.; Bruno, V.; Sortino, M.A. Estrogen Receptors and Type 1 Metabotropic Glutamate Receptors Are Interdependent in Protecting Cortical Neurons against β-Amyloid Toxicity. Mol. Pharmacol. 2012, 81, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Micevych, P.; Sinchak, K. Estradiol regulation of progesterone synthesis in the brain. Mol. Cell. Endocrinol. 2008, 290, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudry, M.; Bi, X.; Aguirre, C. Progesterone–estrogen interactions in synaptic plasticity and neuroprotection. Neuroscience 2013, 239, 280–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagard, R.; Van Tan, H.; Creuzet, C.; Pelloux, H. Differential Development of Toxoplasma gondii in Neural Cells. Parasitol. Today 1999, 15, 504–507. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galván Ramírez, M.d.l.L.; Dueñas-Jiménez, J.M.; Gutiérrez-Maldonado, A.F.; Rodríguez Pérez, L.R. Effect of 17β-Estradiol, Progesterone, and Tamoxifen on Neurons Infected with Toxoplasma gondii In Vitro. Microorganisms 2021, 9, 2174. https://doi.org/10.3390/microorganisms9102174
Galván Ramírez MdlL, Dueñas-Jiménez JM, Gutiérrez-Maldonado AF, Rodríguez Pérez LR. Effect of 17β-Estradiol, Progesterone, and Tamoxifen on Neurons Infected with Toxoplasma gondii In Vitro. Microorganisms. 2021; 9(10):2174. https://doi.org/10.3390/microorganisms9102174
Chicago/Turabian StyleGalván Ramírez, María de la Luz, Judith Marcela Dueñas-Jiménez, Adrián Fernando Gutiérrez-Maldonado, and Laura Rocío Rodríguez Pérez. 2021. "Effect of 17β-Estradiol, Progesterone, and Tamoxifen on Neurons Infected with Toxoplasma gondii In Vitro" Microorganisms 9, no. 10: 2174. https://doi.org/10.3390/microorganisms9102174