Larvae of the Clothing Moth Tineola bisselliella Maintain Gut Bacteria that Secrete Enzyme Cocktails to Facilitate the Digestion of Keratin


Abstract
:1. Introduction
2. Materials and Methods
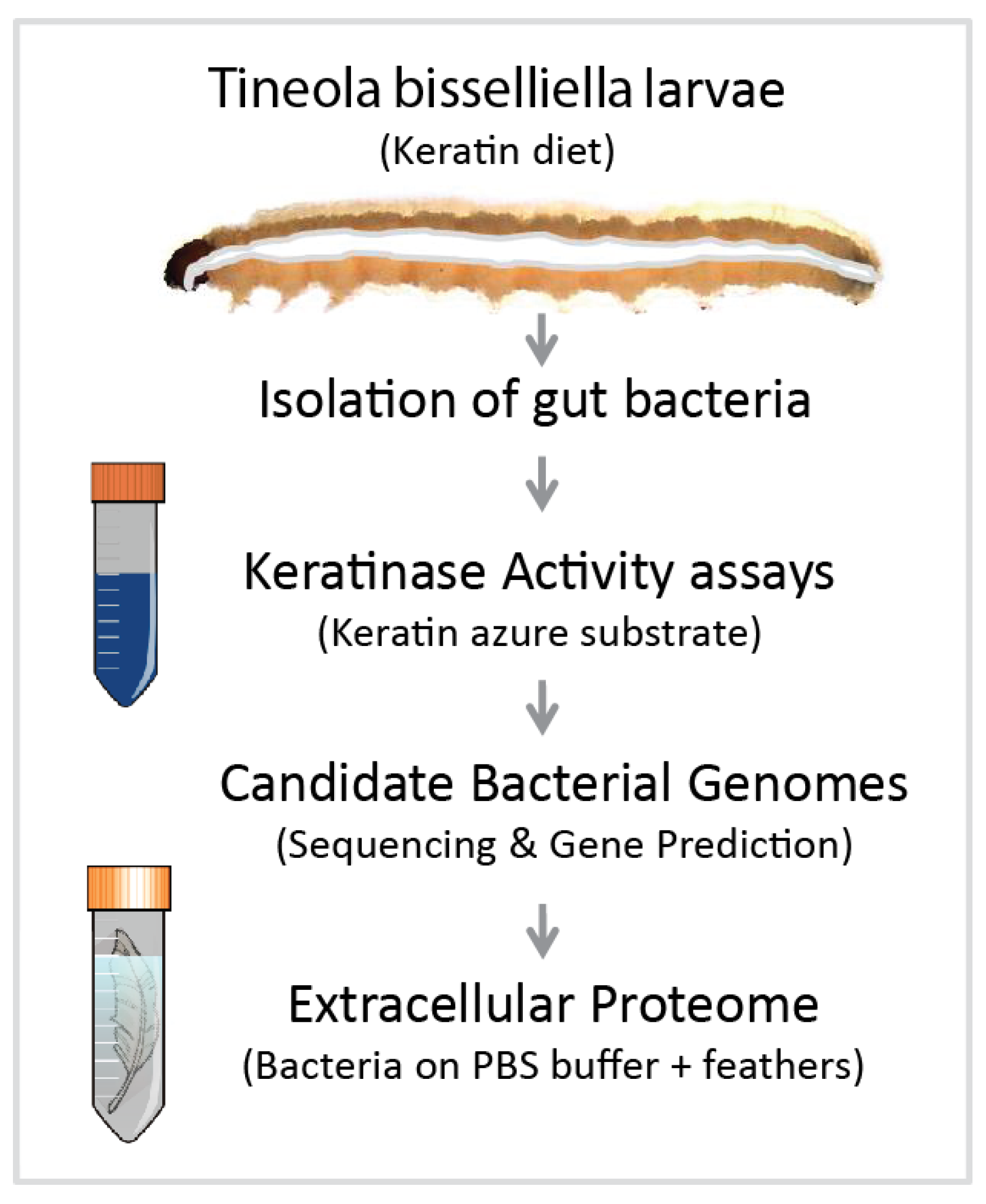
2.1. Insect Rearing and Bacterial Isolation
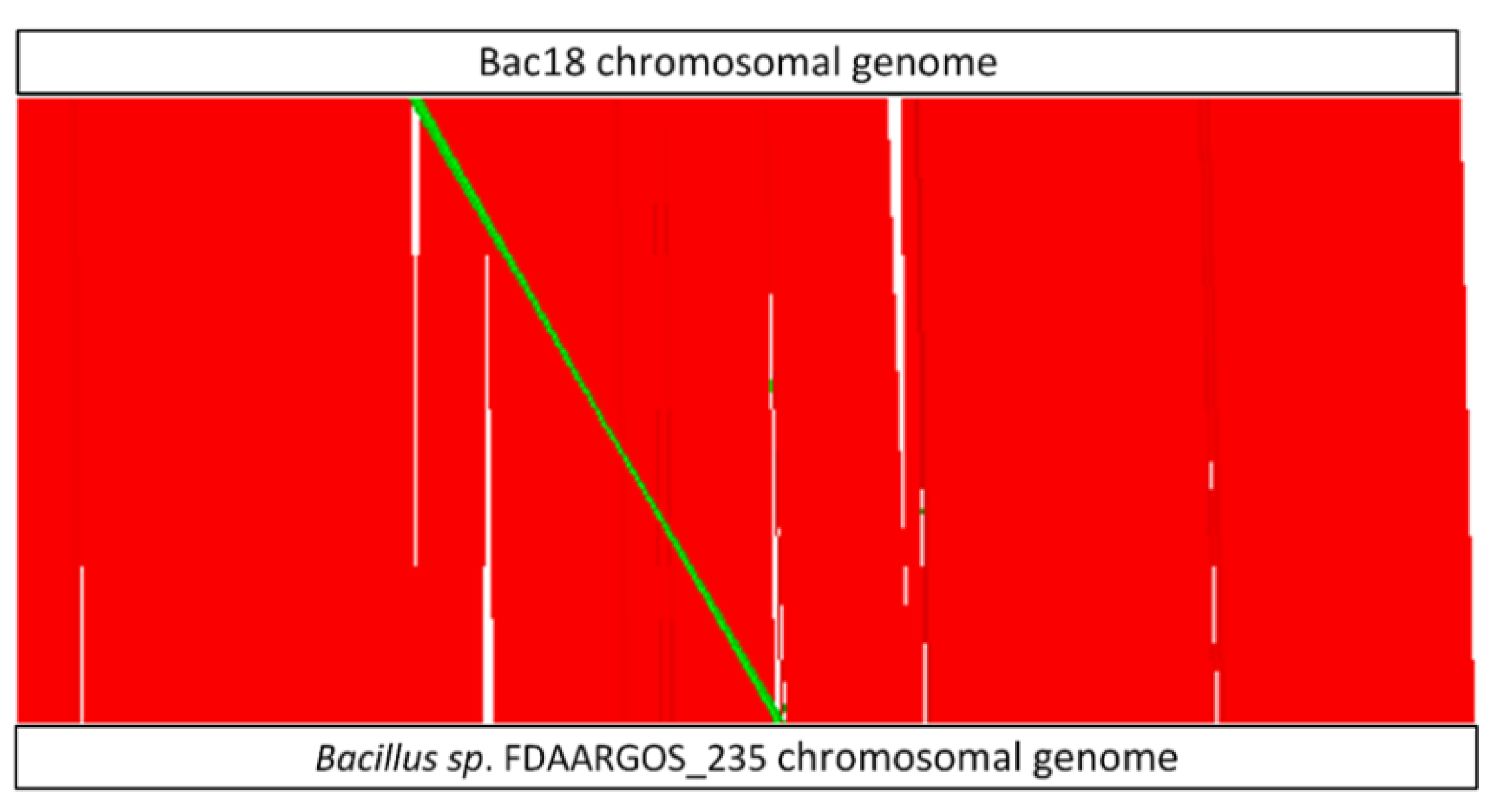
2.2. Genome Sequencing and Characterization of Bac18 Isolated from T. bisselliella
2.3. Target Gene Prediction
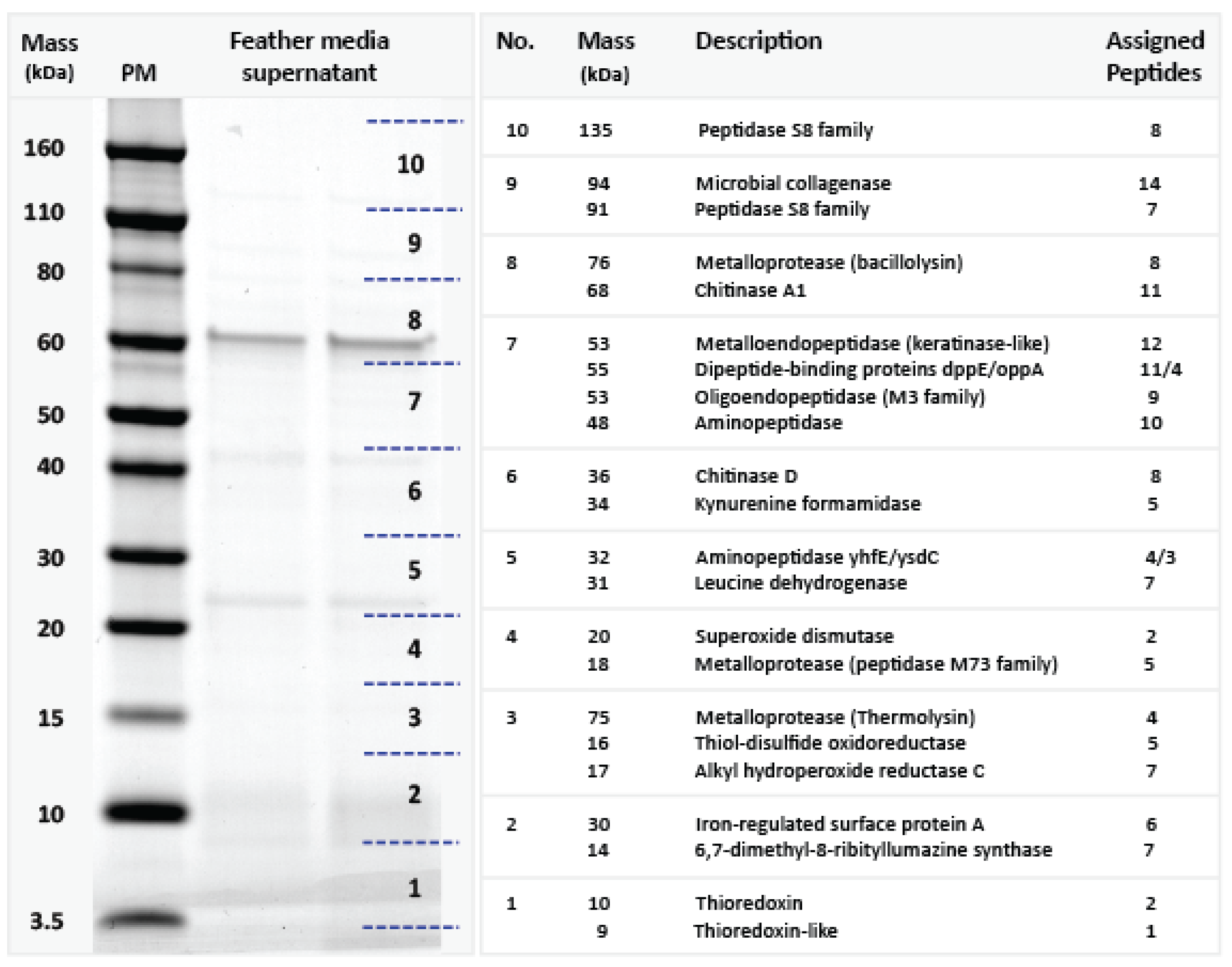
2.4. Proteomic Analysis
3. Results
3.1. Identification of Bacterial Candidates and Genome Sequencing
3.2. Candidate Gene Prediction and Proteomic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berasategui, A.; Shukla, S.; Salem, H.; Kaltenpoth, M. Potential applications of insect symbionts in biotechnology. Appl. Microbiol. Biotechnol. 2016, 100, 1567–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plarre, R.; Krüger-Carstensen, B. An attempt to reconstruct the natural and cultural history of the webbing clothes moth Tineola bisselliella Hummel (Lepidoptera: Tineidae). J. Entomol. Acarol. Res. 2011, 43, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Singh, H.; Anwar, S.; Chattopadhyay, A.; Tiwari, K.K.; Kaur, S.; Dhilon, G. Keratin: Dissolution, extraction and biomedical application. Crit. Rev. Biotechnol. 2017, 37, 476–491. [Google Scholar] [CrossRef]
- Wan, W.-L.; Chen, M.-Y.; Tu, I.-F.; Lin, Y.-C.; EswarKumar, N.; Chen, M.-Y.; Ho, M.-C.; Wu, S.-H. The discovery of novel heat-stable keratinases from Meiothermus taiwanensis WR-220 and other extremophiles. Sci. Rep. 2017, 7, 4658. [Google Scholar] [CrossRef]
- Hughes, J.; Vogler, A.P. Gene expression in the gut of keratin-feeding clothes moths (Tineola) and keratin beetles (Trox) revealed by subtracted cDNA libraries. Insect Biochem. Mol. Biol. 2006, 36, 584–592. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Hunt, M.; Silva, N.D.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Balentin, F.; Vallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Schwengers, O.; Hain, T.; Chakraborty, T.; Goesmann, A. ReferenceSeeker: Rapid Determination of Appropriate Reference Genomes. GitHub. 2019. Available online: https://github.com/oschwengers/referenceseeker (accessed on 13 September 2020).
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Skaljac, M.; Vogel, H.; Wielsch, N.; Mihajlovic, S.; Vilcinskas, A. Transmission of a protease-secreting bacterial symbiont among pea aphids via host plants. Front. Physiol. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, D.F. Wool digestion and mothproofing. In Advances in Pest Control Research, 2nd ed.; Metcalf, R.L., Ed.; Interscience: New York, NY, USA, 1958; pp. 207–262. [Google Scholar]
- Ramnani, P.; Gupta, R. Keratinases vis-a-vis conventional proteases and feather degradation. World J. Microbiol. Biotechnol. 2007, 23, 1537–1540. [Google Scholar] [CrossRef]
- Christeller, J.T.; Markwick, N.P.; Burgess, E.P. Midgut proteinase activities of three keratinolytic larvae, Hofmannophila pseudospretella, Tineola bisseliella, and Anthrenocerus australis, and the effect of proteinase inhibitors on proteolysis. Arch. Insect Biochem. Physiol. 1994, 25, 159–173. [Google Scholar] [CrossRef]
- Yoshimura, T.; Tabata, H.; Nishio, M.; Ide, E.; Yamaoka, R.; Hayashiya, K. L-Cystein lyase of the webbing clothes moth, Tineola bisselliella. Insect Biochem. 1988, 18, 771–777. [Google Scholar] [CrossRef]
- Crewther, W.G.; McQuade, A.B. The intestinal microflora of the clothes moth Tineola bisselliella in relation to wool digestion. J. Gen. Microbiol. 1955, 12, 311–313. [Google Scholar] [CrossRef] [Green Version]
- Kasper, C.S. Ultrastructure of the Digestive System of the Larval Clothes Moth Tineola bisselliella (Humm.). Ph.D. Thesis, Boston University Graduate School, Boston, MA, USA, 1978. Unpublished work. [Google Scholar]
- Shannon, A.L.; Attwood, G.; Hopcroft, D.H.; Christeller, J.T. Characterization of lactic acid bacteria in the larval midgut of the keratinophagous lepidopteran, Hofmannophila pseudospretella. Lett. Appl. Microbiol. 2001, 32, 36–41. [Google Scholar] [CrossRef]
- Daroit, D.J.; Brandelli, A. A current assessment on the production of bacterial keratinases. Crit. Rev. Biotechnol. 2014, 34, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Morita, Y.; Hasan, Q.; Yokoyama, K.; Tamiya, E. Keratin degradation: A cooperative action of two enzymes from Stenotrophomonas sp. Biochem. Biophys. Res. Commun. 2002, 294, 1138–1143. [Google Scholar] [CrossRef]
- Lange, L.; Huang, Y.; Busk, P.K. Microbial decomposition of keratin in nature—A new hypothesis of industrial relevance. Appl. Microbiol. Biotechnol. 2016, 100, 2083–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandelli, A.; Daroit, D.J.; Riffel, A. Biochemical features of microbial keratinases and their production and applications. Appl. Microbiol. Biotechnol. 2010, 85, 1735–1750. [Google Scholar] [CrossRef]
- Gupta, R.; Ramnani, P. Microbial keratinases and their prospective applications: An overview. Appl. Microbiol. Biotechnol. 2006, 70, 21–33. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, T.; Song, N.; Li, Q.; Wang, Z.; Zhang, X.; Lu, X.; Fang, J.; Chen, J. Purification and characterization of four key enzymes from a feather-degrading Bacillus subtilis from the gut of tarantula Chilobrachys guangxiensis. Int. Biodeterior. Biodegrad. 2014, 96, 26–32. [Google Scholar] [CrossRef]
- Mereghetti, V.; Chouaia, B.; Montagna, M. New insights into the microbiota of moth pests. Int. J. Mol. Sci. 2017, 18, 2450. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bac18 | ||
|---|---|---|
| Contig Name | Contig Length [bp] | Sequencing Coverage |
| Bac18_0 | • 5,236,241 | 131 |
| Bac18_2 | 745,926 | 164 |
| Bac18_3 | • 16,484 | 479 |
| Bac18_4 | 39,074 | 365 |
| Bac18_5 | 12,019 | 167 |
| Bac18_6 | 14,098 | 173 |
| ∑ 6,063,842 | ||
| ID Related Genome | ANI | Cons. DNA | Taxonomy ID | Organism Name | |
|---|---|---|---|---|---|
| Bac18 | GCF_002073415.2 | 99.56% | 83.93% | 1839798 | Bacillus sp. FDAARGOS_235 |
| GCF_000300475.1 | 98.54% | 84.50% | 1195464 | Bacillus thuringiensis MC28 | |
| GCF_000496285.1 | 99.44% | 78.04% | 1415784 | Bacillus toyonensis BCT-7112 |
| Prokka ID | Prokka Annotation | Pfam Domains | Keratinase Blast Hit | Proteomics ID |
|---|---|---|---|---|
| PROKKA_00129 | Aminopeptidase YwaD precursor | PA, Peptidase_M28 | n. h. | 02318 |
| PROKKA_00135 | Thermolysin precursor | FTP, Peptidase_M4_C, Peptidase_M4 | n. h. | 02311 |
| PROKKA_00413 | Neutral protease B precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | A0A0B4ZU77 | 02032 |
| PROKKA_00751 | Peptidase propeptide and YPEB domain protein | PepSY | n. h. | 01721 |
| PROKKA_01081 | Minor extracellular protease vpr precursor | Peptidase_S8, Inhibitor_I9, PA | n. h. | 01389 |
| PROKKA_01290 | Peptidase T | Peptidase_M28 * | n. h. | 01180 |
| PROKKA_01747 | Microbial collagenase precursor | Peptidase_S8 *, PPC | n. h. | 00731 |
| PROKKA_02134 | Bacillolysin precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | n. h. | 00344 |
| PROKKA_02729 | Bacillolysin precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | n. h. | |
| PROKKA_02930 | Thermolysin precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | n. h. | |
| PROKKA_03102 | Thermitase | Peptidase_S8 | F8SVT0 ** | |
| PROKKA_03248 | Bacillolysin precursor | FTP *, PepSY | n. h. | 02311 |
| PROKKA_03479 | Intracellular serine protease | Peptidase_S8 | Q45GC8 | |
| PROKKA_04411 | hypothetical protein | PepSY | n. h. | 03334 |
| PROKKA_04743 | Minor extracellular protease vpr | Peptidase_S8, Inhibitor_I9, PA | n. h. | 03020 |
| PROKKA_04780 | Bacillolysin precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | A0A0B4ZU77 | 02981 |
| PROKKA_05541 | Major intracellular serine protease | Peptidase_S8 * | n. h. | |
| PROKKA_05686 | Neutral protease B precursor | FTP, PepSY, Peptidase_M4, Peptidase_M4_C | A0A0B4ZU77 | 02032 |
| PROKKA_05790 | Thermophilic serine proteinase | Peptidase_S8 * | n. h. | |
| PROKKA_05803 | Chitinase A1 precursor | Peptidase_S8 * | n. h. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilcinskas, A.; Schwabe, M.; Brinkrolf, K.; Plarre, R.; Wielsch, N.; Vogel, H. Larvae of the Clothing Moth Tineola bisselliella Maintain Gut Bacteria that Secrete Enzyme Cocktails to Facilitate the Digestion of Keratin. Microorganisms 2020, 8, 1415. https://doi.org/10.3390/microorganisms8091415
Vilcinskas A, Schwabe M, Brinkrolf K, Plarre R, Wielsch N, Vogel H. Larvae of the Clothing Moth Tineola bisselliella Maintain Gut Bacteria that Secrete Enzyme Cocktails to Facilitate the Digestion of Keratin. Microorganisms. 2020; 8(9):1415. https://doi.org/10.3390/microorganisms8091415
Chicago/Turabian StyleVilcinskas, Andreas, Michael Schwabe, Karina Brinkrolf, Rudy Plarre, Natalie Wielsch, and Heiko Vogel. 2020. "Larvae of the Clothing Moth Tineola bisselliella Maintain Gut Bacteria that Secrete Enzyme Cocktails to Facilitate the Digestion of Keratin" Microorganisms 8, no. 9: 1415. https://doi.org/10.3390/microorganisms8091415