Feature of Adhesins Produced by Human Clinical Isolates of Mycobacterium intracellulare, Mycobacterium intracellulare subsp. chimaera and Closely Related Species

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Strain Identification and Genotyping
2.2.1. Species Identification by GenoType CM and Sequence Analysis of the hsp65 Gene
2.2.2. MLVA Genotyping
2.3. PCR DNA Fragment Analysis
2.4. Construction of Bioluminescent and Fluorescent M. intracelullare Strain
2.5. Cytoadherence of M. intracelullare to A549 Epithelial Cells
2.6. Purification by Heparin-Sepharose Chromatography
2.7. Identification of the Purified M. intracellulare Proteins by High-Resolution Mass Spectrometry Analysis
2.8. SDS-PAGE and Immunoblotting
2.9. ELISA
2.10. Nucleotide Sequence Accession Numbers
3. Results
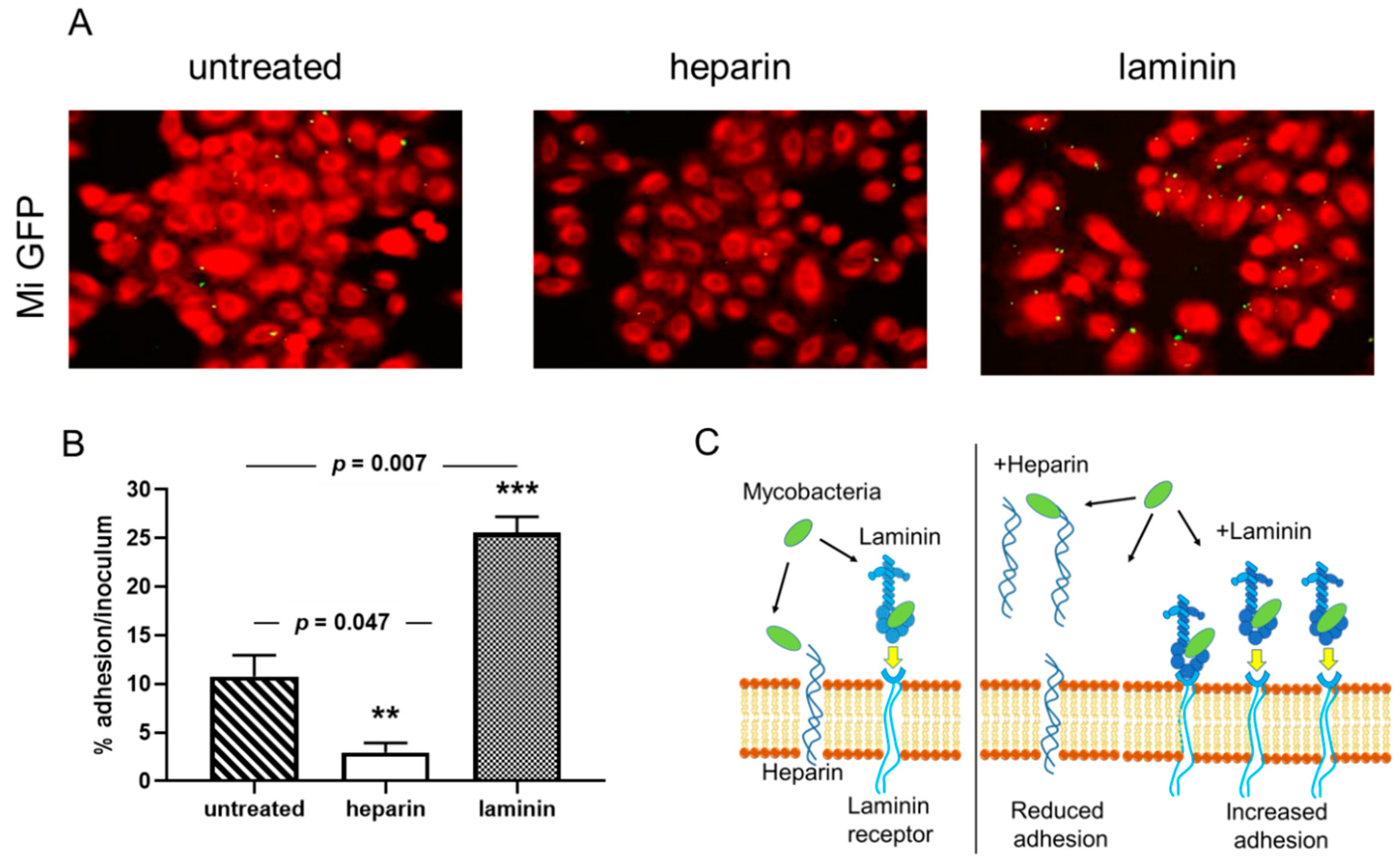
3.1. Adherence of M. intracellulare to Epithelial Cells is Modulated by Heparin and Laminin
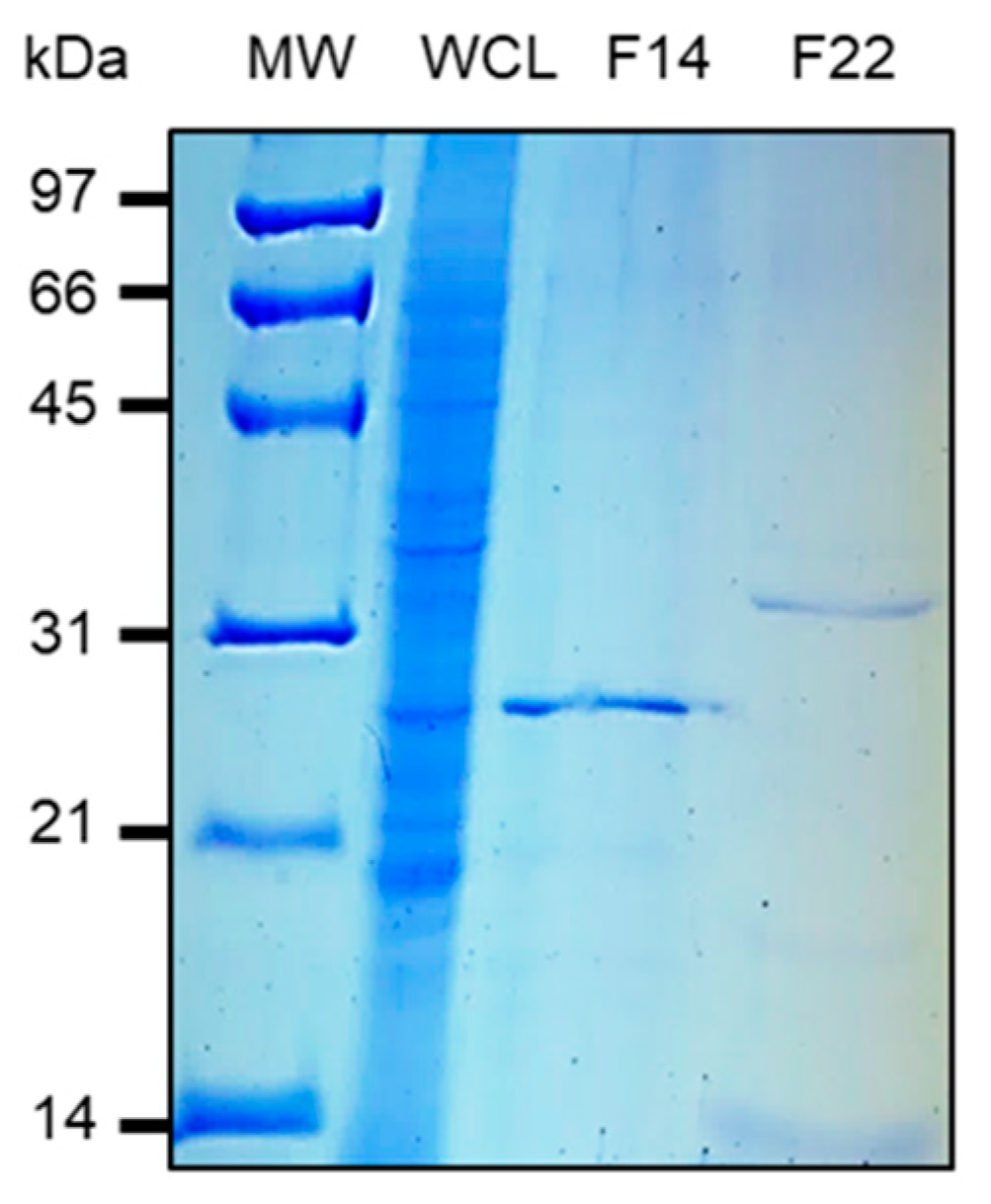
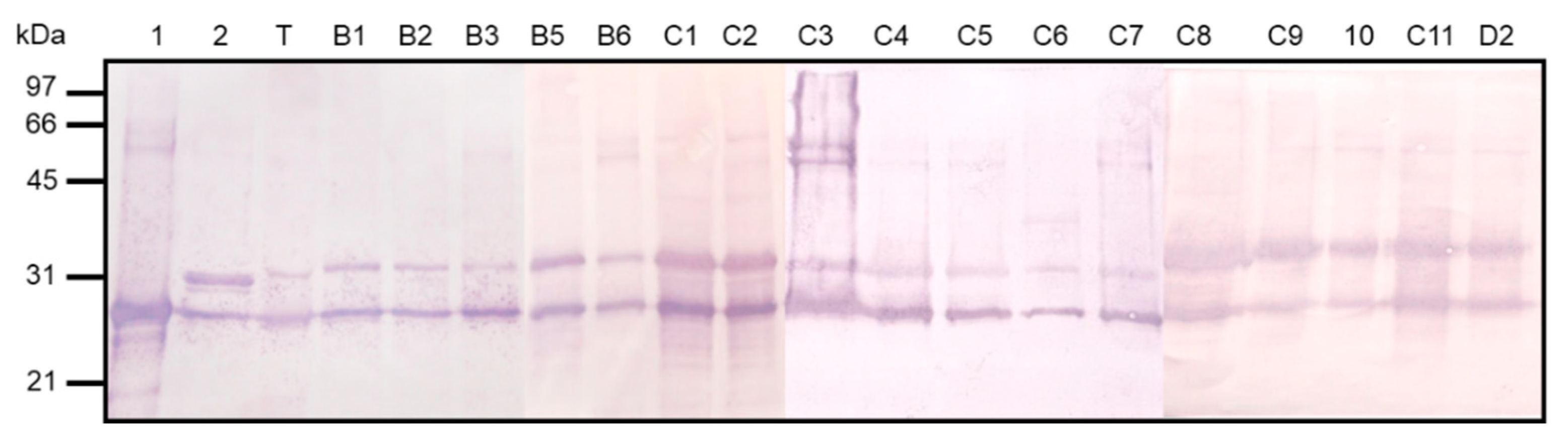
3.2. Purification and Characterization of HBHA and LBP from M. intracellulare by Heparin-Sepharose Chromatography
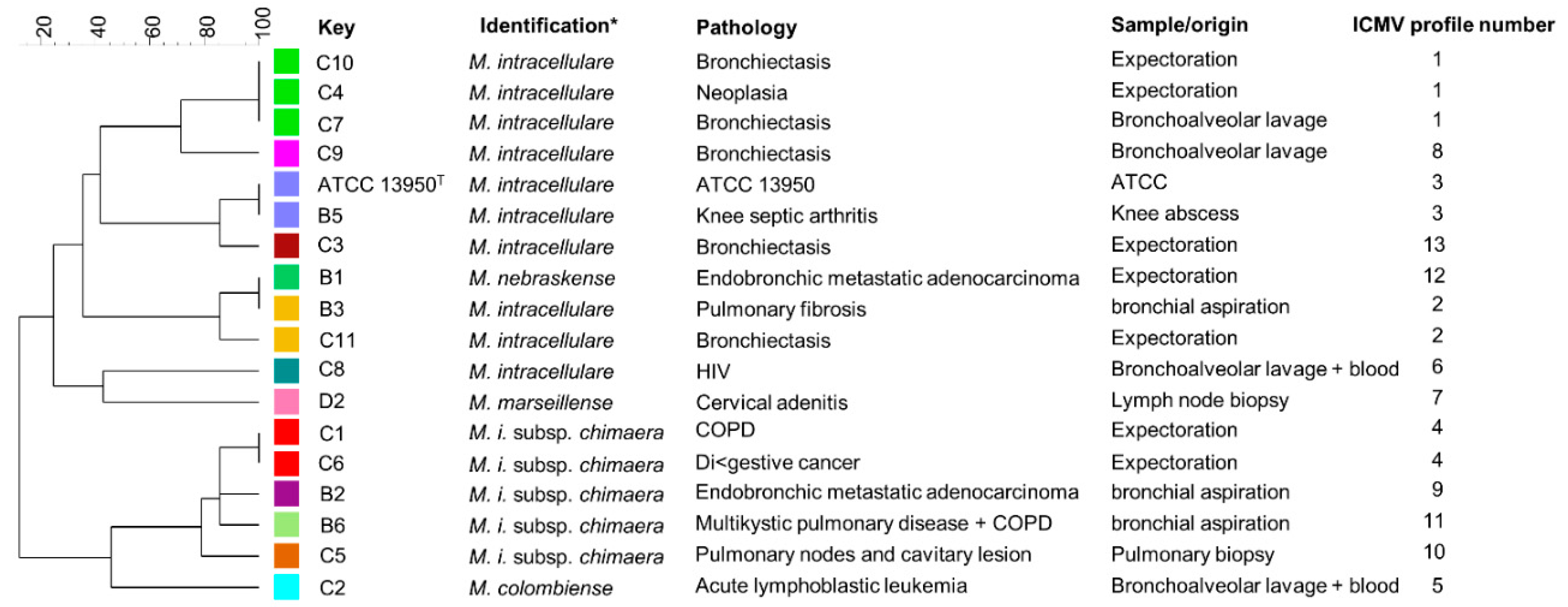
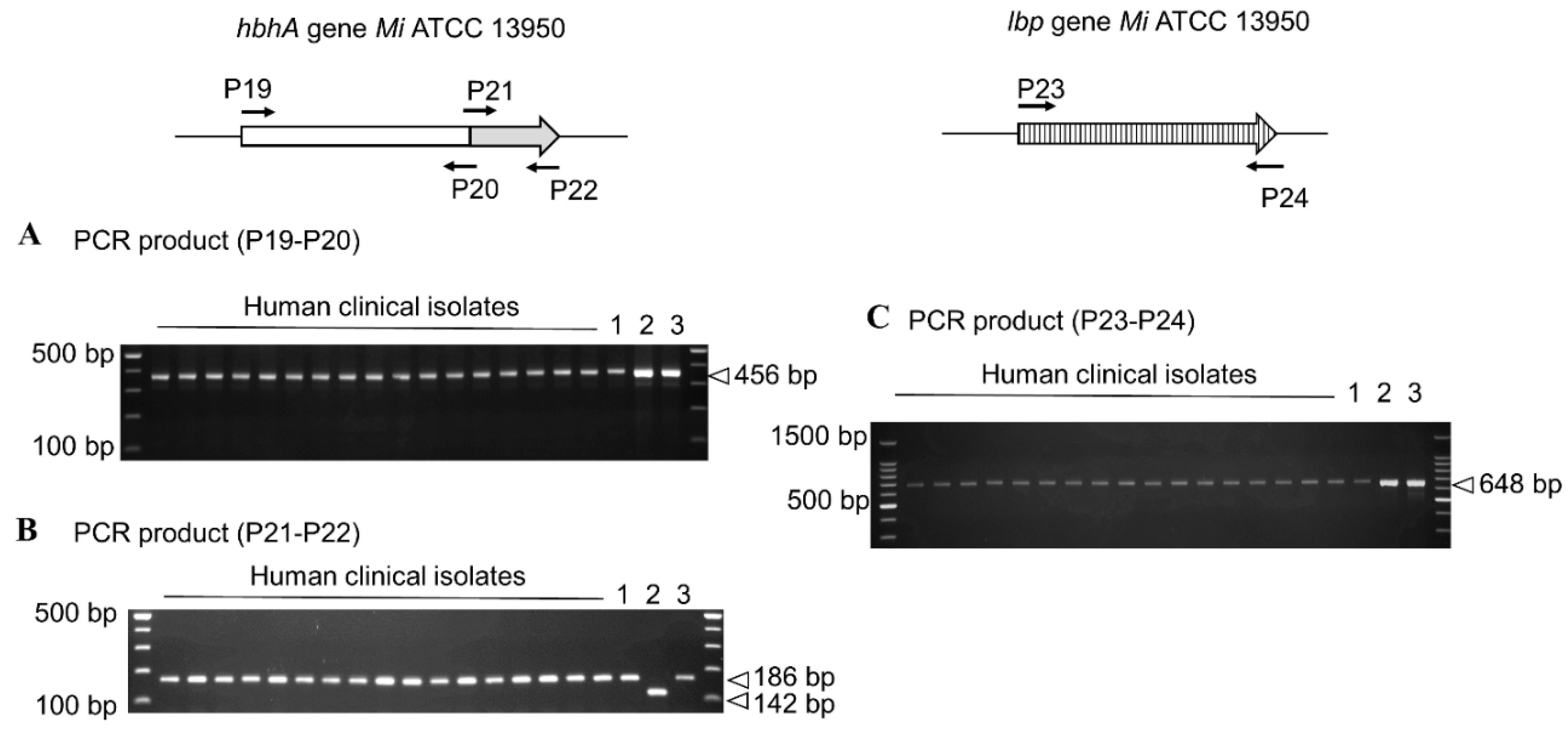
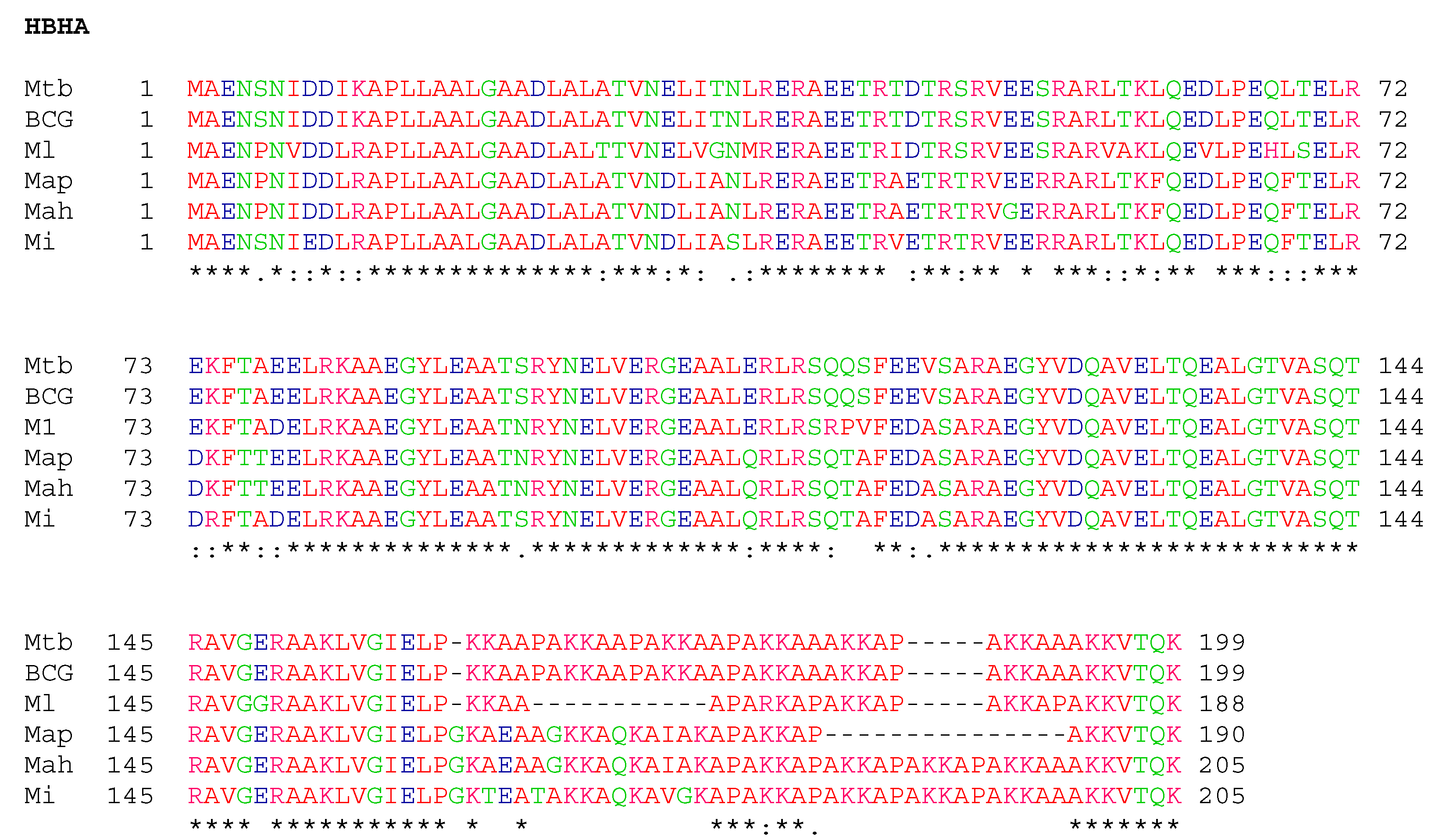
3.3. Conservation of HBHA and LBP within the Panel of Clinical M. intracellulare Isolates and Closely Related Species
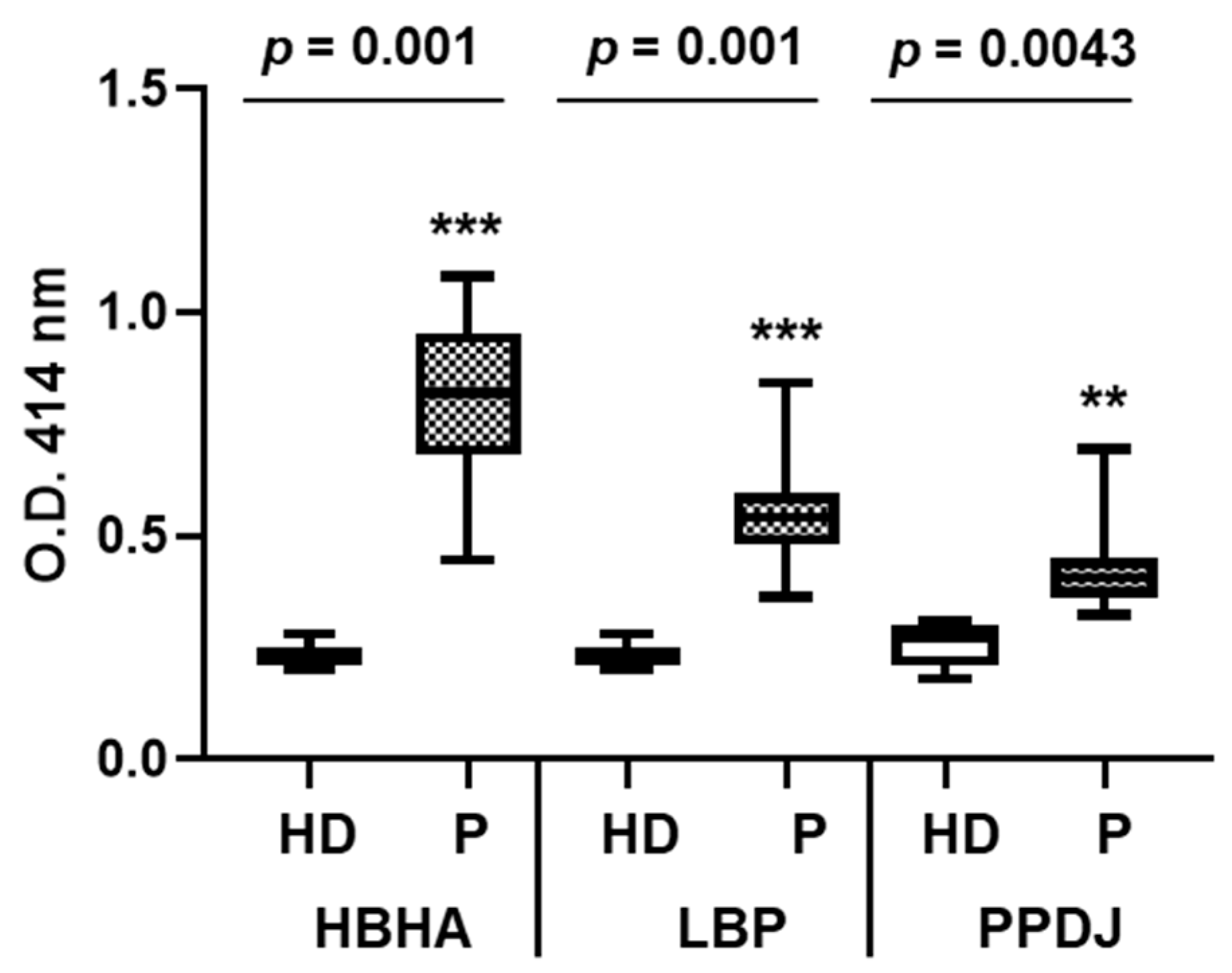
3.4. Both HBHA and LBP are Recognized by Sera from Infected Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dartois, V.; Sizemore, C.; Dick, T. Editorial: NTM-The New Uber-Bugs. Front. Microbiol. 2019, 10, 1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Hoefsloot, W.; van Ingen, J.; Andrejak, C.; Angeby, K.; Bauriaud, R.; Bemer, P.; Beylis, N.; Boeree, M.J.; Cacho, J.; Chihota, V.; et al. The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: An NTM-NET collaborative study. Eur. Respir. J. 2013, 42, 1604–1613. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Shin, S.H.; Moon, S.M.; Yang, B.; Kim, H.; Kwon, O.J.; Huh, H.J.; Ki, C.S.; Lee, N.Y.; Shin, S.J.; et al. Distribution and clinical significance of Mycobacterium avium complex species isolated from respiratory specimens. Diagn. Microbiol. Infect. Dis 2017, 88, 125–137. [Google Scholar] [CrossRef]
- Falkinham, J.O., 3rd. Epidemiology of infection by nontuberculous mycobacteria. Clin. Microbiol. Rev. 1996, 9, 177–215. [Google Scholar] [CrossRef]
- Lacroix, A.; Piau, C.; Lanotte, P.; Carricajo, A.; Guillouzouic, A.; Peuchant, O.; Cady, A.; Dupin, C.; Fangous, M.S.; Martin, C.; et al. Emergence of Nontuberculous Mycobacterial Lymphadenitis in Children After the Discontinuation of Mandatory Bacillus Calmette and GuErin Immunization in France. Pediatr. Infect. Dis. J. 2018, 37, e257–e260. [Google Scholar] [CrossRef]
- Johne, H.A.; Frothingham, L. Ein eigenthuemlicher fall von tuberkulose beim rind. Deutsche Zeitschrift fur tiermedicin und pathologie 1895, 21, 438–454. [Google Scholar]
- Thorel, M.F.; Krichevsky, M.; Levy-Frebault, V.V. Numerical taxonomy of mycobactin-dependent mycobacteria, emended description of Mycobacterium avium, and description of Mycobacterium avium subsp. avium subsp. nov., Mycobacterium avium subsp. paratuberculosis subsp. nov., and Mycobacterium avium subsp. silvaticum subsp. nov. Int. J. Syst. Bacteriol. 1990, 40, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Mijs, W.; de Haas, P.; Rossau, R.; Van der Laan, T.; Rigouts, L.; Portaels, F.; van Soolingen, D. Molecular evidence to support a proposal to reserve the designation Mycobacterium avium subsp. avium for bird-type isolates and ‘M. avium subsp. hominissuis’ for the human/porcine type of M. avium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1505–1518. [Google Scholar] [CrossRef]
- Van Ingen, J.; Turenne, C.Y.; Tortoli, E.; Wallace, R.J., Jr.; Brown-Elliott, B.A. A definition of the Mycobacterium avium complex for taxonomical and clinical purposes, a review. Int. J. Syst. Evol. Microbiol. 2018, 68, 3666–3677. [Google Scholar] [CrossRef]
- Tortoli, E.; Meehan, C.J.; Grottola, A.; Fregni Serpini, G.; Fabio, A.; Trovato, A.; Pecorari, M.; Cirillo, D.M. Genome-based taxonomic revision detects a number of synonymous taxa in the genus Mycobacterium. Infect. Genet. Evol. 2019, 75, 103983. [Google Scholar] [CrossRef] [PubMed]
- Tortoli, E.; Rindi, L.; Garcia, M.J.; Chiaradonna, P.; Dei, R.; Garzelli, C.; Kroppenstedt, R.M.; Lari, N.; Mattei, R.; Mariottini, A.; et al. Proposal to elevate the genetic variant MAC-A, included in the Mycobacterium avium complex, to species rank as Mycobacterium chimaera sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Sax, H.; Bloemberg, G.; Hasse, B.; Sommerstein, R.; Kohler, P.; Achermann, Y.; Rossle, M.; Falk, V.; Kuster, S.P.; Bottger, E.C.; et al. Prolonged Outbreak of Mycobacterium chimaera Infection After Open-Chest Heart Surgery. Clin. Infect. Dis. 2015, 61, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, H.J.; Kim, S.Y.; Shim, H.J.; Kim, D.H.; Yoo, I.Y.; Kang, O.K.; Ki, C.S.; Shin, S.Y.; Jhun, B.W.; Shin, S.J.; et al. GenoType NTM-DR Performance Evaluation for Identification of Mycobacterium avium Complex and Mycobacterium abscessus and Determination of Clarithromycin and Amikacin Resistance. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wang, L.; Ma, P.; Fan, W.; Gu, B.; Ju, S. Accuracy of Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry for Identification of Mycobacteria: A systematic review and meta-analysis. Sci. Rep. 2018, 8, 4131. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Fleury, C.; Jalalvand, F.; Riesbeck, K. Human pathogens utilize host extracellular matrix proteins laminin and collagen for adhesion and invasion of the host. FEMS Microbiol. Rev. 2012, 36, 1122–1180. [Google Scholar] [CrossRef] [Green Version]
- Menozzi, F.D.; Bischoff, R.; Fort, E.; Brennan, M.J.; Locht, C. Molecular characterization of the mycobacterial heparin-binding hemagglutinin, a mycobacterial adhesin. Proc. Natl. Acad. Sci. USA 1998, 95, 12625–12630. [Google Scholar] [CrossRef] [Green Version]
- Menozzi, F.D.; Rouse, J.H.; Alavi, M.; Laude-Sharp, M.; Muller, J.; Bischoff, R.; Brennan, M.J.; Locht, C. Identification of a heparin-binding hemagglutinin present in mycobacteria. J. Exp. Med. 1996, 184, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Biet, F.; Angela de Melo Marques, M.; Grayon, M.; Xavier da Silveira, E.K.; Brennan, P.J.; Drobecq, H.; Raze, D.; Vidal Pessolani, M.C.; Locht, C.; Menozzi, F.D. Mycobacterium smegmatis produces an HBHA homologue which is not involved in epithelial adherence. Microbes Infect. 2007, 9, 175–182. [Google Scholar] [CrossRef]
- Lefrancois, L.H.; Bodier, C.C.; Cochard, T.; Canepa, S.; Raze, D.; Lanotte, P.; Sevilla, I.A.; Stevenson, K.; Behr, M.A.; Locht, C.; et al. Novel feature of Mycobacterium avium subsp. paratuberculosis, highlighted by characterization of the heparin-binding hemagglutinin adhesin. J. Bacteriol. 2013, 195, 4844–4853. [Google Scholar] [CrossRef] [Green Version]
- Eraghi, V.; Derakhshandeh, A.; Hosseini, A.; Haghkhah, M.; Sechi, L.A.; Motamedi Boroojeni, A. Recombinant fusion protein of Heparin-Binding Hemagglutinin Adhesin and Fibronectin Attachment Protein (rHBHA-FAP) of Mycobacterium avium subsp. paratuberculosis elicits a strong gamma interferon response in peripheral blood mononuclear cell culture. Gut Pathog. 2019, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Sechi, L.A.; Ahmed, N.; Felis, G.E.; Dupre, I.; Cannas, S.; Fadda, G.; Bua, A.; Zanetti, S. Immunogenicity and cytoadherence of recombinant heparin binding haemagglutinin (HBHA) of Mycobacterium avium subsp. paratuberculosis: Functional promiscuity or a role in virulence? Vaccine 2006, 24, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, P.; Raze, D.; Fritzinger, B.; Wieruszeski, J.M.; Biet, F.; Dose, A.; Carpentier, M.; Schwarzer, D.; Allain, F.; Lippens, G.; et al. Differential contribution of the repeats to heparin binding of HBHA, a major adhesin of Mycobacterium tuberculosis. PLoS ONE 2012, 7, e32421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pethe, K.; Alonso, S.; Biet, F.; Delogu, G.; Brennan, M.J.; Locht, C.; Menozzi, F.D. The heparin-binding haemagglutinin of M. tuberculosis is required for extrapulmonary dissemination. Nature 2001, 412, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Masungi, C.; Temmerman, S.; Van Vooren, J.P.; Drowart, A.; Pethe, K.; Menozzi, F.D.; Locht, C.; Mascart, F. Differential T and B cell responses against Mycobacterium tuberculosis heparin-binding hemagglutinin adhesin in infected healthy individuals and patients with tuberculosis. J. Infect. Dis. 2002, 185, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temmerman, S.; Pethe, K.; Parra, M.; Alonso, S.; Rouanet, C.; Pickett, T.; Drowart, A.; Debrie, A.S.; Delogu, G.; Menozzi, F.D.; et al. Methylation-dependent T cell immunity to Mycobacterium tuberculosis heparin-binding hemagglutinin. Nat. Med. 2004, 10, 935–941. [Google Scholar] [CrossRef]
- Bitti, M.L.; Masala, S.; Capasso, F.; Rapini, N.; Piccinini, S.; Angelini, F.; Pierantozzi, A.; Lidano, R.; Pietrosanti, S.; Paccagnini, D.; et al. Mycobacterium avium subsp. paratuberculosis in an Italian cohort of type 1 diabetes pediatric patients. Clin. Dev. Immunol. 2012, 2012, 785262. [Google Scholar] [CrossRef] [Green Version]
- Rosu, V.; Ahmed, N.; Paccagnini, D.; Gerlach, G.; Fadda, G.; Hasnain, S.E.; Zanetti, S.; Sechi, L.A. Specific immunoassays confirm association of Mycobacterium avium Subsp. paratuberculosis with type-1 but not type-2 diabetes mellitus. PLoS ONE 2009, 4, e4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Melo Marques, M.A.; Mahapatra, S.; Nandan, D.; Dick, T.; Sarno, E.N.; Brennan, P.J.; Vidal Pessolani, M.C. Bacterial and host-derived cationic proteins bind alpha2-laminins and enhance Mycobacterium leprae attachment to human Schwann cells. Microbes Infect. 2000, 2, 1407–1417. [Google Scholar] [CrossRef]
- Shimoji, Y.; Ng, V.; Matsumura, K.; Fischetti, V.A.; Rambukkana, A. A 21-kDa surface protein of Mycobacterium leprae binds peripheral nerve laminin-2 and mediates Schwann cell invasion. Proc. Natl. Acad. Sci. USA 1999, 96, 9857–9862. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.A.; Danelishvili, L.; McNamara, M.; Berredo-Pinho, M.; Bildfell, R.; Biet, F.; Rodrigues, L.S.; Oliveira, A.V.; Bermudez, L.E.; Pessolani, M.C. Interaction of Mycobacterium leprae with human airway epithelial cells: Adherence, entry, survival, and identification of potential adhesins by surface proteome analysis. Infect. Immun. 2013, 81, 2645–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefrancois, L.H.; Pujol, C.; Bodier, C.C.; Teixeira-Gomez, A.P.; Drobecq, H.; Rosso, M.L.; Raze, D.; Dias, A.A.; Hugot, J.P.; Chacon, O.; et al. Characterization of the Mycobacterium avium subsp. paratuberculosis laminin-binding/histone-like protein (Lbp/Hlp) which reacts with sera from patients with Crohn’s disease. Microbes Infect. 2011, 13, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Soares de Lima, C.; Zulianello, L.; Marques, M.A.; Kim, H.; Portugal, M.I.; Antunes, S.L.; Menozzi, F.D.; Ottenhoff, T.H.; Brennan, P.J.; Pessolani, M.C. Mapping the laminin-binding and adhesive domain of the cell surface-associated Hlp/LBP protein from Mycobacterium leprae. Microbes Infect. 2005, 7, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Telenti, A.; Marchesi, F.; Balz, M.; Bally, F.; Bottger, E.C.; Bodmer, T. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 1993, 31, 175–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauchy, F.A.; Degrange, S.; Charron, A.; Dupon, M.; Xin, Y.; Bebear, C.; Maugein, J. Variable-number tandem-repeat markers for typing Mycobacterium intracellulare strains isolated in humans. BMC Microbiol. 2010, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, I.R.; Stewart, G.R.; Turner, D.J.; Patel, J.; Karamanou, D.; Snelgrove, R.J.; Young, D.B. A role for dendritic cells in the dissemination of mycobacterial infection. Microbes Infect. 2006, 8, 1339–1346. [Google Scholar] [CrossRef]
- Lefrancois, L.H.; Bodier, C.C.; Lecher, S.; Gilbert, F.B.; Cochard, T.; Harichaux, G.; Labas, V.; Teixeira-Gomes, A.P.; Raze, D.; Locht, C.; et al. Purification of native HBHA from Mycobacterium avium subsp. paratuberculosis. BMC Res. Notes 2013, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M.; Gautron, J.; Berges, M.; Attucci, S.; Le Blay, G.; Labas, V.; Nys, Y.; Rehault-Godbert, S. Antimicrobial potential of egg yolk ovoinhibitor, a multidomain Kazal-like inhibitor of chicken egg. J. Agric. Food Chem. 2011, 59, 12368–12374. [Google Scholar] [CrossRef]
- Pethe, K.; Puech, V.; Daffe, M.; Josenhans, C.; Drobecq, H.; Locht, C.; Menozzi, F.D. Mycobacterium smegmatis laminin-binding glycoprotein shares epitopes with Mycobacterium tuberculosis heparin-binding haemagglutinin. Mol. Microbiol. 2001, 39, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Rouse, D.A.; Morris, S.L.; Karpas, A.B.; Mackall, J.C.; Probst, P.G.; Chaparas, S.D. Immunological characterization of recombinant antigens isolated from a Mycobacterium avium lambda gt11 expression library by using monoclonal antibody probes. Infect. Immun. 1991, 59, 2595–2600. [Google Scholar] [CrossRef] [Green Version]
- Pethe, K.; Aumercier, M.; Fort, E.; Gatot, C.; Locht, C.; Menozzi, F.D. Characterization of the heparin-binding site of the mycobacterial heparin-binding hemagglutinin adhesin. J. Biol. Chem. 2000, 275, 14273–14280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallstrom, T.; Haupt, K.; Kraiczy, P.; Hortschansky, P.; Wallich, R.; Skerka, C.; Zipfel, P.F. Complement regulator-acquiring surface protein 1 of Borrelia burgdorferi binds to human bone morphogenic protein 2, several extracellular matrix proteins, and plasminogen. J. Infect. Dis. 2010, 202, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pethe, K.; Bifani, P.; Drobecq, H.; Sergheraert, C.; Debrie, A.S.; Locht, C.; Menozzi, F.D. Mycobacterial heparin-binding hemagglutinin and laminin-binding protein share antigenic methyllysines that confer resistance to proteolysis. Proc. Natl. Acad. Sci. USA 2002, 99, 10759–10764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbiere, V.; Segers, J.; Desmet, R.; Lecher, S.; Loyens, M.; Petit, E.; Melnyk, O.; Locht, C.; Mascart, F. Natural T Cell Epitope Containing Methyl Lysines on Mycobacterial Heparin-Binding Hemagglutinin. J. Immunol. 2020, 204, 1715–1723. [Google Scholar] [CrossRef]
- Parra, M.; Pickett, T.; Delogu, G.; Dheenadhayalan, V.; Debrie, A.S.; Locht, C.; Brennan, M.J. The mycobacterial heparin-binding hemagglutinin is a protective antigen in the mouse aerosol challenge model of tuberculosis. Infect. Immun. 2004, 72, 6799–6805. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Identification | Pathology | Specimen/Sample | Reference |
|---|---|---|---|---|
| B1 | M. nebraskense | Endobronchic metastatic adenocarcinoma | Expectoration | This study |
| B2 | M. i. subsps chimaera | Endobronchic metastatic adenocarcinoma | bronchial aspiration | This study |
| B3 | M. intracellulare | Pulmonary fibrosis | bronchial aspiration | This study |
| B5 | M. intracellulare | Knee septic arthritis | Knee abscess | This study |
| B6 | M. i. subsps chimaera | Multikysitic pulmonary disease + COPD * | bronchial aspiration | This study |
| C1 | M. i. subsps chimaera | COPD | Expectoration | This study |
| C2 | M. colombiense | Acute lymphoblastic leukemia | Bronchoalveolar lavage + blood | This study |
| C3 | M. intracellulare | Bronchiectasis | Expectoration | This study |
| C4 | M. intracellulare | Neoplasia | Expectoration | This study |
| C5 | M. i. subsps. chimaera | Pulmonary nodes and cavitary lesion | Pulmonary biopsy | This study |
| C6 | M. i. subsps chimaera | Digestive cancer | Expectoration | This study |
| C7 | M. intracellulare | Bronchiectasis | Bronchoalveolar lavage | This study |
| C8 | M. intracellulare | HIV | Bronchoalveolar lavage + blood | This study |
| C9 | M. intracellulare | Bronchiectasis | Bronchoalveolar lavage | This study |
| C10 | M. intracellulare | Bronchiectasis | Expectoration | This study |
| C11 | M. intracellulare | Bronchiectasis | Expectoration | This study |
| D2 | M. marseillense | Cervical adenitis | Lymph node biopsy | This study |
| ATCC13950 | M. intracellulare | Type strain | ATCC | |
| Plasmid | Description | |||
| pSMT3LxEGFP | Lux, GFP, (Hygromycin) |
| Name | Target 1 Gene 1 | Sequence | Use |
|---|---|---|---|
| P1 | Hsp65-F | ACCAACGATGGTGTGTCCAT | |
| P2 | Hsp65-R | CTTGTCGAACCGCATACCCT | Subspecies |
| P3 | rpoB-F | GGCAAGGTCACCCCGAAGGG | identification |
| P4 | rpoB-R | AGCGGCTGCTGGGTGATCATC | |
| P5 | Min18-F | GGATTCGGCCGCGCAATTC | |
| P6 | Min18-R | GCCGAACCATTTGGCGAAC | |
| P7 | Min19-F | TAGGGGCAGGTCATCGAAG | |
| P8 | Min19-R | CATGGTTCGCCCTCTACAC | |
| P9 | Min20-F | CGACGCCGATGACGTAAAC | |
| P10 | Min20-R | GCTGAGCTACAGCCTCGAC | |
| P11 | Min22-F | AGCTCGTGACGACGGAAAC | MLVA |
| P12 | Min22-R | TCAGGAATGGGTCCGGTTC | Typing 2 |
| P13 | Min31-F | GCTCTATGACGACCTCAAG | |
| P14 | Min31-R | CGACCGCATCCAGAAACAG | |
| P15 | Min33-F | GGCGTTGAACACGTTGGTG | |
| P16 | Min33-R | GTGCAGTTCAACCACGAAC | |
| P17 | MIRU3 new-F | GCAAGCCGGGAACCGGATCG | |
| P18 | MIRU3 new-R | CACCACGGTGGCCTCAAAGC | |
| P19 | HBHA-F | TATACATATGACCATGGCGGAAAACCCGAACATCG | |
| P20 | HBHA-intra-R | GCCGACCGCGCGGGTCTGCGA | Polymorphism analysis |
| P21 | HBHA intra-F | TATAGAATTCCGCCAAGCTGGTGGGCATCGAGCTGCCG | and |
| P22 | HBHA-R | CTACCTACTTCTGGGTGACCTTCTTGGC | sequencing |
| P23 | LBP-F | ATGAATAAGGCAGAGCTC | |
| P24 | LBP-R | CTACCGGCGGCCGCGACGCG |
| Peptides Identified by HR-MS/MS * | Start–End Position | Peptide Modification |
|---|---|---|
| HBHA | ||
| 70LQEDLPEQFTELR82 | 70–82 | |
| 93AAEGYLEAATSR104 | 93–104 | |
| 105YNELVER111 | 105–111 | |
| 119LRSQTAFEDASAR131 | 119–131 | |
| 121SQTAFEDASAR131 | 121–131 | |
| 132AEGYVDQAVELTQEALGTVASQTR155 | 132–155 | |
| 161AAKmeLVGIELPGK172 | 161–172 | Methylated |
| 164LVGIELPGKTEATAKme178 | 164–178 | Methylated |
| LBP | ||
| 9AELIDVLTQK18 | 9–18 | |
| 25QATAAVENVVDTIVR39 | 25–39 | |
| 40AVHKGDSVTITGFGVFEQR58 | 40–58 | |
| 44GDSVTITGFGVFEQR58 | 44–58 | |
| 76VKPTSVPAFRPGAQFK91 | 76–91 | |
| 92AVVSGAQRLPSDGPAVK108 | 92–108 | |
| 92AVVSGAQRLPSDGPAVKmeR109 | 92–109 | Methylated |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefrancois, L.H.; Cochard, T.; Branger, M.; Peuchant, O.; Conde, C.; Pastuszka, A.; Locht, C.; Lanotte, P.; Biet, F. Feature of Adhesins Produced by Human Clinical Isolates of Mycobacterium intracellulare, Mycobacterium intracellulare subsp. chimaera and Closely Related Species. Microorganisms 2020, 8, 1154. https://doi.org/10.3390/microorganisms8081154
Lefrancois LH, Cochard T, Branger M, Peuchant O, Conde C, Pastuszka A, Locht C, Lanotte P, Biet F. Feature of Adhesins Produced by Human Clinical Isolates of Mycobacterium intracellulare, Mycobacterium intracellulare subsp. chimaera and Closely Related Species. Microorganisms. 2020; 8(8):1154. https://doi.org/10.3390/microorganisms8081154
Chicago/Turabian StyleLefrancois, Louise H., Thierry Cochard, Maxime Branger, Olivia Peuchant, Cyril Conde, Adeline Pastuszka, Camille Locht, Philippe Lanotte, and Franck Biet. 2020. "Feature of Adhesins Produced by Human Clinical Isolates of Mycobacterium intracellulare, Mycobacterium intracellulare subsp. chimaera and Closely Related Species" Microorganisms 8, no. 8: 1154. https://doi.org/10.3390/microorganisms8081154