Analysis of Tick Surface Decontamination Methods



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Tick Sample Collection
2.3. Contamination and Decontamination of Ticks
2.4. Nucleic Acid Extraction
2.5. Quantitative PCR Amplification for DNA and cDNA Samples
2.6. Bacterial 16S rRNA Gene Sequencing for DNA and cDNA Samples
2.7. Bioinformatics
2.8. Statistics
2.9. Nucleotide Sequence Accession Numbers
3. Results
3.1. Reduction of Artificial Bacterial Contaminants
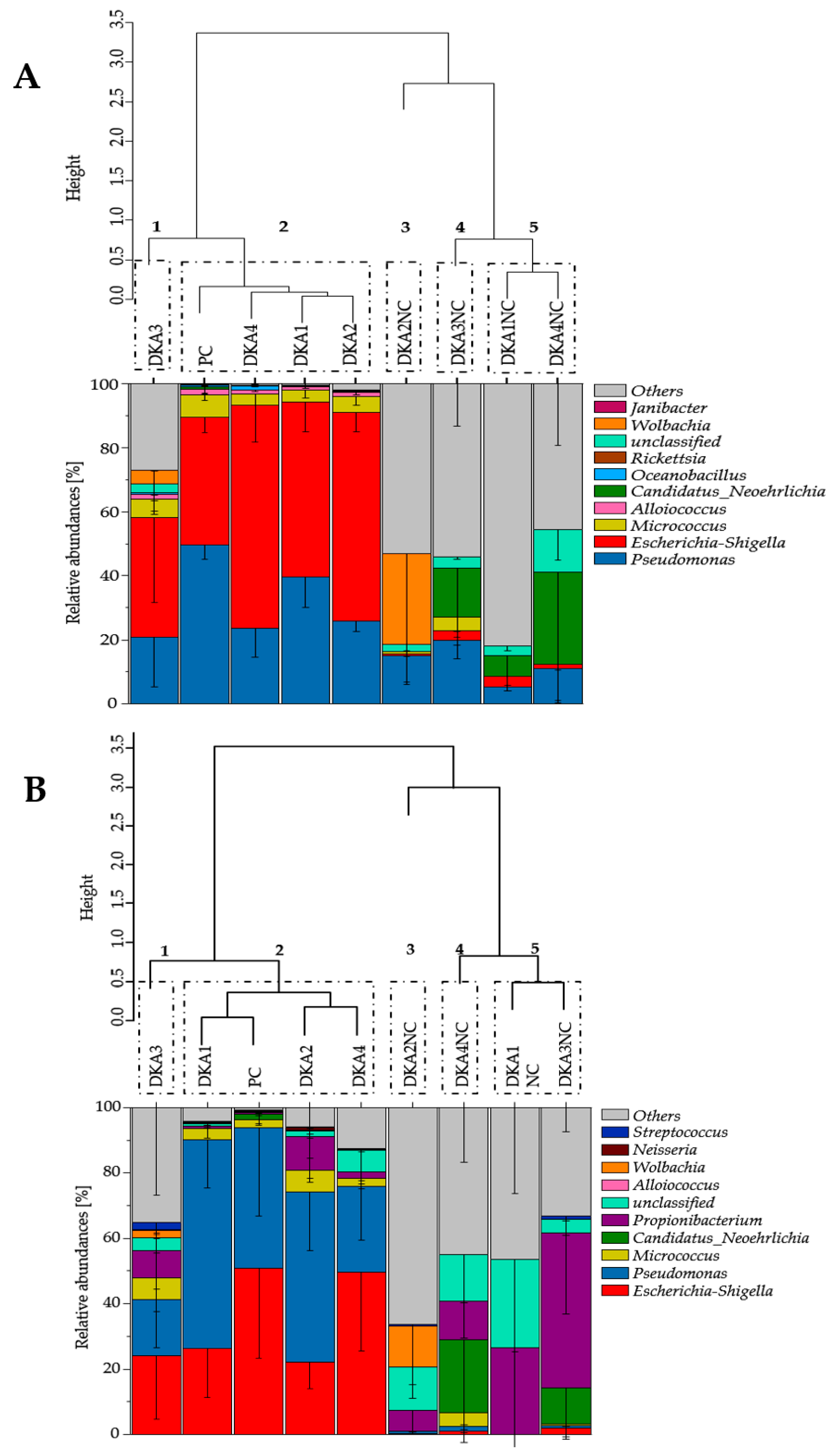
3.2. Decontamination Strategy Shifted Bacterial Contamination Diversity
3.3. Effect of Decontamination Treatment on Tick-Borne Microbiome
4. Discussion
4.1. Tick Decontamination by 5% Sodium Hypochlorite Treatment Was More Efficient Compared to Other Decontamination Treatments
4.2. Gram-Positive Contaminants Were Removed Less Efficiently Compared to Gram-Negative Contaminants
4.3. Effect of 5% Sodium Hypochlorite Treatment on Tick-Borne Microbiome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasit. Vectors 2018, 11, 12. [Google Scholar] [CrossRef]
- Jongejan, F. and Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, 3–14. [Google Scholar] [CrossRef]
- Dobler, G.; Fingerle, V.; Hagedorn, P.; Pfeffer, M.; Silaghi, C.; Tomaso, H.; Henning, K.; Niedrig, M. Gefahren der Übertragung von Krankheitserregern durch Schildzecken in Deutschland. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2014, 57, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Hellenbrand, W.; Kreusch, T.; Böhmer, M.M.; Wagner-Wiening, C.; Dobler, G.; Wichmann, O.; Altmann, D. Epidemiology of Tick-Borne Encephalitis (TBE) in Germany, 2001–2018. Pathogens 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Dressel, K.M.; Niedrig, M.; Mertens, M.; Schüle, S.A.; Groschup, M.H. Public health and vector-borne diseases—A new concept for risk governance. Zoonoses Public Health 2013, 60, 528–538. [Google Scholar] [CrossRef]
- Noll, M.; Buettner, C.; Lasota, S. Copper containing wood preservatives shifted bacterial and fungal community compositions in pine sapwood in two field sites. Int. Biodeterior. Biodegrad. 2019, 142, 26–35. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Budachetri, K.; Williams, J.; Mukherjee, N.; Sellers, M.; Moore, F.; Karim, S. The microbiome of neotropical ticks parasitizing on passerine migratory birds. Ticks Tick Borne Dis. 2017, 8, 170–173. [Google Scholar] [CrossRef] [Green Version]
- Gall, C.A.; Reif, K.E.; Scoles, G.A.; Mason, K.L.; Mousel, M.; Noh, S.M.; Brayton, K.A. The bacterial microbiome of Dermacentor andersoni ticks influences pathogen susceptibility. ISME J. 2016, 10, 1846–1855. [Google Scholar] [CrossRef] [Green Version]
- Swei, A.; Kwan, J.Y. Tick microbiome and pathogen acquisition altered by host blood meal. ISME J. 2017, 11, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnusamy, L.; Gonzalez, A.; van Treuren, W.; Weiss, S.; Parobek, C.M.; Juliano, J.J.; Knight, R.; Roe, R.M.; Apperson, C.S.; Meshnick, S.R. Diversity of Rickettsiales in the microbiome of the lone star tick, Amblyomma americanum. Appl. Environ. Microbiol. 2014, 80, 354–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binetruy, F.; Dupraz, M.; Buysse, M.; Duron, O. Surface sterilization methods impact measures of internal microbial diversity in ticks. Parasit. Vectors 2019, 12, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Dosmann, A.; Bahet, N.; Gordon, D.M. Experimental modulation of external microbiome affects nestmate recognition in harvester ants (Pogonomyrmex barbatus). PeerJ 2016, 4, e1566. [Google Scholar] [CrossRef] [Green Version]
- Keiser, C.N.; Shearer, T.A.; DeMarco, A.E.; Brittingham, H.A.; Knutson, K.A.; Kuo, C.; Zhao, K.; Pruitt, J.N. Cuticular bacteria appear detrimental to social spiders in mixed but not monoculture exposure. Curr. Zool. 2016, 62, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Mattoso, T.C.; Moreira, D.D.O.; Samuels, R.I. Symbiotic bacteria on the cuticle of the leaf-cutting ant Acromyrmex subterraneus subterraneus protect workers from attack by entomopathogenic fungi. Biol. Lett. 2012, 8, 461–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, E.A.; Kong, H.H.; Renaud, G.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Wolfsberg, T.G.; Turner, M.L.; Segre, J.A. A diversity profile of the human skin microbiota. Genome Res. 2008, 18, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Merrick, M.V.; Simpson, J.D.; Liddell, S. Skin decontamination—A comparison of four methods. Br. J. Radiol. 1982, 55, 317–318. [Google Scholar] [CrossRef]
- Chan, H.P.; Zhai, H.; Hui, X.; Maibach, H.I. Skin decontamination: Principles and perspectives. Toxicol. Ind. Health 2013, 29, 955–968. [Google Scholar] [CrossRef]
- Davies, B.M.; Patel, H.C. Systematic review and meta-analysis of preoperative antisepsis with combination chlorhexidine and povidone-iodine. Surg. J. (N Y) 2016, 2, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Machado-Ferreira, E.; Piesman, J.; Zeidner, N.S.; Soares, C.A.G. A prevalent alpha-proteobacterium Paracoccus sp. in a population of the Cayenne ticks (Amblyomma cajennense) from Rio de Janeiro, Brazil. Genet. Mol. Biol. 2012, 35, 862–867. [Google Scholar] [CrossRef] [PubMed]
- Menchaca, A.C.; Visi, D.K.; Strey, O.F.; Teel, P.D.; Kalinowski, K.; Allen, M.S.; Williamson, P.C. Preliminary assessment of microbiome changes following blood-feeding and survivorship in the Amblyomma americanum nymph-to-adult transition using semiconductor sequencing. PLoS ONE 2013, 8, e67129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, K.A.; Gall, C.A.; Mason, K.L.; Scoles, G.A.; Brayton, K.A. The characterization and manipulation of the bacterial microbiome of the Rocky Mountain wood tick, Dermacentor andersoni. Parasit. Vectors 2015, 8, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall, C.A.; Scoles, G.A.; Magori, K.; Mason, K.L.; Brayton, K.A. Laboratory colonization stabilizes the naturally dynamic microbiome composition of field collected Dermacentor andersoni ticks. Microbiome 2017, 5, 133. [Google Scholar] [CrossRef] [Green Version]
- Paolin, A.; Trojan, D.; Carniato, A.; Tasca, F.; Massarin, E.; Tugnoli, A.; Cogliati, E. Analysis of the effectiveness of sodium hypochlorite decontamination of cadaveric human tissues at retrieval. Cell Tissue Bank. 2016, 17, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Carpi, G.; Cagnacci, F.; Wittekindt, N.E.; Zhao, F.; Qi, J.; Tomsho, L.P.; Drautz, D.I.; Rizzoli, A.; Schuster, S.C. Metagenomic profile of the bacterial communities associated with Ixodes ricinus ticks. PLoS ONE 2011, 6, e25604. [Google Scholar] [CrossRef]
- Fischer, M.; Renevey, N.; Thür, B.; Hoffmann, D.; Beer, M.; Hoffmann, B. Efficacy assessment of nucleic acid decontamination reagents used in molecular diagnostic laboratories. PLoS ONE 2016, 11, e0159274. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Hurst, C.G.; Kirk, M.A.; Reedy, S.J.D.; Braue, E.H. Reactive skin decontamination lotion (RSDL) for the decontamination of chemical warfare agent (CWA) dermal exposure. Curr. Pharm. Biotechnol. 2012, 13, 1971–1979. [Google Scholar] [CrossRef]
- Thors, L.; Koch, M.; Wigenstam, E.; Koch, B.; Hägglund, L.; Bucht, A. Comparison of skin decontamination efficacy of commercial decontamination products following exposure to VX on human skin. Chem. Biol. Interact. 2017, 273, 82–89. [Google Scholar] [CrossRef]
- Bakken, L.R.; Frostegård, Å. Nucleic Acid Extraction from Soil. In Nucleic Acids and Proteins in Soil; Nannipieri, P., Smalla, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 49–73. [Google Scholar]
- Ballantyne, K.N.; Salemi, R.; Guarino, F.; Pearson, J.R.; Garlepp, D.; Fowler, S.; van Oorschot, R.A.H. DNA contamination minimisation—Finding an effective cleaning method. Aust. J. Forensic Sci. 2015, 47, 428–439. [Google Scholar] [CrossRef]
- Gray, J.S. Studies on the larval activity of the tick Ixodes ricinus L. in Co. Wicklow, Ireland. Exp. Appl. Acarol. 1985, 1, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Okła, H.; Sosnowska, M.; Jasik, K.P.; Slodki, J.A.N.; Wojtyczka, R.D. Nonspecific Bacterial Flora Isolated from the Body Surface and Inside Ixodes ricinus Ticks. Pol. J. Microbiol. 2012, 61, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.M.; Liu, L.; Jutras, B.L.; Yadav, A.K.; Narasimhan, S.; Gopalakrishnan, V.; Ansari, J.M.; Jefferson, K.K.; Cava, F.; Jacobs-Wagner, C.; et al. Pathogen-mediated manipulation of arthropod microbiota to promote infection. Proc. Natl. Acad. Sci. USA 2017, 114, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buresová, V.; Franta, Z.; Kopácek, P. A comparison of Chryseobacterium indologenes pathogenicity to the soft tick Ornithodoros moubata and hard tick Ixodes ricinus. J. Invertebr. Pathol. 2006, 93, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Khoo, J.-J.; Chen, F.; Kho, K.L.; Ahmad Shanizza, A.I.; Lim, F.-S.; Tan, K.-K.; Chang, L.-Y.; AbuBakar, S. Bacterial community in Haemaphysalis ticks of domesticated animals from the Orang Asli communities in Malaysia. Ticks Tick Borne Dis. 2016, 7, 929–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, T.; Okoda, Y.; Badgar, B.; Inoue, N.; Xuan, X.; Taylor, D.; Fujisaki, K. Fate of GFP-expressing Escherichia coli in the midgut and response to ingestion in a tick, Ornithodoros moubata (Acari: Argasidae). Exp. Parasitol. 2004, 108, 67–73. [Google Scholar] [CrossRef]
- Urbanová, V.; Hartmann, D.; Grunclová, L.; Šíma, R.; Flemming, T.; Hajdušek, O.; Kopáček, P. IrFC—An Ixodes ricinus injury-responsive molecule related to Limulus Factor C. Dev. Comp. Immunol. 2014, 46, 439–447. [Google Scholar] [CrossRef]
- Ruparell, A.; Inui, T.; Staunton, R.; Wallis, C.; Deusch, O.; Holcombe, L.J. The canine oral microbiome: Variation in bacterial populations across different niches. BMC Microbiol. 2020, 20, 42. [Google Scholar] [CrossRef] [Green Version]
- Teufel, L.; Pipal, A.; Schuster, K.C.; Staudinger, T.; Redl, B. Material-dependent growth of human skin bacteria on textiles investigated using challenge tests and DNA genotyping. J. Appl. Microbiol. 2010, 108, 450–461. [Google Scholar] [CrossRef]
- Kemp, B.M.; Smith, D.G. Use of bleach to eliminate contaminating DNA from the surface of bones and teeth. Forensic Sci. Int. 2005, 154, 53–61. [Google Scholar] [CrossRef]
- Noll, M.; Matthies, D.; Frenzel, P.; Derakshani, M.; Liesack, W. Succession of bacterial community structure and diversity in a paddy soil oxygen gradient. Environ. Microbiol. 2005, 7, 382–395. [Google Scholar] [CrossRef]
- Huijsdens, X.W.; Linskens, R.K.; Mak, M.; Meuwissen, S.G.M.; Vandenbroucke-Grauls, C.M.J.E.; Savelkoul, P.H.M. Quantification of bacteria adherent to gastrointestinal mucosa by real-time PCR. J. Clin. Microbiol. 2002, 40, 4423–4427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, R.; Bestervelt, L.L.; Donofrio, R.S. Development and validation of a real-time TaqMan assay for the detection and enumeration of Pseudomonas fluorescens ATCC 13525 used as a challenge organism in testing of food equipments. J. Food Sci. 2012, 77, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.A. Real-Time PCR Arrays. In Real-Time PCR: Advanced Technologies and Applications; Saunders, N.A., Lee, M.A., Eds.; Caister Academic Press: Poole, UK, 2013; pp. 43–68. [Google Scholar]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef]
- FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 30 June 2020).
- Buettner, C.; Noll, M. Differences in microbial key players in anaerobic degradation between biogas and sewage treatment plants. Int. Biodeterior. Biodegrad. 2018, 133, 124–132. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, R.A.; Doranalli, K.; Rinttilä, T.; Vienola, K.; Jurgens, G.; Apajalahti, J. The impact of Bacillus subtilis DSM 32315 on the pathology, performance, and intestinal microbiome of broiler chickens in a necrotic enteritis challenge. Poult. Sci. 2019, 98, 3450–3463. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Updating the 97% identity threshold for 16S ribosomal RNA OTUs. Bioinformatics 2018, 34, 2371–2375. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-4. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 4 May 2020).
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 28 April 2020).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 13 December 2019).
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- Galili, T.; Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; et al. Various R Programming Tools for Plotting Data. Available online: https://github.com/talgalili/gplots (accessed on 30 April 2020).
- Nakao, R.; Abe, T.; Nijhof, A.M.; Yamamoto, S.; Jongejan, F.; Ikemura, T.; Sugimoto, C. A novel approach, based on BLSOMs (Batch Learning Self-Organizing Maps), to the microbiome analysis of ticks. ISME J. 2013, 7, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Frough-Reyhani, M.; Ghasemi, N.; Soroush-Barhaghi, M.; Amini, M.; Gholizadeh, Y. Antimicrobial efficacy of different concentration of sodium hypochlorite on the biofilm of Enterococcus faecalis at different stages of development. J. Clin. Exp. Dent. 2016, 8, e480–e484. [Google Scholar] [CrossRef] [PubMed]
- Estrela, C.; Estrela, C.R.A.; Barbin, E.L.; Spanó, J.C.E.; Marchesan, M.A.; Pécora, J.D. Mechanism of action of sodium hypochlorite. Braz. Dent. J. 2002, 13, 113–117. [Google Scholar] [CrossRef]
- Soares, J.A.; Pires Júnior, D.R. Influence of sodium hypochlorite-based irrigants on the susceptibility of intracanal microbiota to biomechanical preparation. Braz. Dent. J. 2006, 17, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Fouad, A.F. Endodontic Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Mai-Prochnow, A.; Clauson, M.; Hong, J.; Murphy, A.B. Gram positive and gram negative bacteria differ in their sensitivity to cold plasma. Sci. Rep. 2016, 6, 38610. [Google Scholar] [CrossRef] [Green Version]
- Wintzingerode, F.V.; Göbel, U.B.; Stackebrandt, E. Determination of microbial diversity in environmental samples: Pitfalls of PCR-based rRNA analysis. FEMS Microbiol. Rev. 1997, 21, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Kampmann, M.-L.; Børsting, C.; Morling, N. Decrease DNA contamination in the laboratories. Forensic Sci. Int. Genet. Suppl. Ser. 2017, 6, 577–578. [Google Scholar] [CrossRef] [Green Version]
- Prince, A.M.; Andrus, L. PCR: How to kill unwanted DNA. BioTechniques 1992, 12, 358–360. [Google Scholar]
- Illumina, I. MiSeq: System Guide. Available online: https://support.illumina.com/content/dam/illumina-support/documents/documentation/system_documentation/miseq/miseq-system-guide-for-miseq-reporter-1000000061014-00.pdf (accessed on 6 December 2019).
- Andreotti, R.; Pérez de León, A.A.; Dowd, S.E.; Guerrero, F.D.; Bendele, K.G.; Scoles, G.A. Assessment of bacterial diversity in the cattle tick Rhipicephalus (Boophilus) microplus through tag-encoded pyrosequencing. BMC Microbiol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Tekin, S.; Dowd, S.E.; Davinic, M.; Bursali, A.; Keskin, A. Pyrosequencing based assessment of bacterial diversity in Turkish Rhipicephalus annulatus and Dermacentor marginatus ticks (Acari: Ixodidae). Parasitol. Res. 2017, 116, 1055–1061. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Decontamination Solution (5 min) | Abbreviation | Contamination | Number of Independent Ticks |
|---|---|---|---|
| 70% ethanol | DKA 1 | yes | 10 |
| DNA Away | DKA 2 | yes | 10 |
| 5% sodium hypochlorite | DKA 3 | yes | 10 |
| Reactive Skin Decontamination Lotion (RSDL) | DKA 4 | yes | 10 |
| 70% ethanol | DKA 1 NC | no | 3 |
| DNA Away | DKA 2 NC | no | 3 |
| 5% Sodium hypochlorite | DKA 3 NC | no | 3 |
| RSDL | DKA 4 NC | no | 3 |
| No | PC | yes | 10 |
| Contaminants | Cell Number mL−1 | Volume of Solution (in Total) | Percentage Volume in Solution per Tick |
|---|---|---|---|
| Micrococcus luteus | 1.1 × 109 | 650 µL | 28.9% |
| Pseudomonas fluorescens | 1.4 × 109 | 650 µL | 28.9% |
| Escherichia coli | 6.5 × 109 | 650 µL | 28.9% |
| Human sweat | not determined | 200 µL | 8.9% |
| Dog salvia | not determined | 100 µL | 4.4% |
| A | DNA | OTU Richness | Shannon | Pielou’s Evenness | S.chao1 * | S.ACE ** |
| DKA 1 | 86 ± 17 | 0.9 ± 0.1 | 0.2 ± 0.0 | 359 ± 95 | 371 ± 118 | |
| DKA 2 | 101 ± 50 | 1.0 ± 0.2 | 0.2 ± 0.1 | 1229 ± 1750 | 697 ± 599 | |
| DKA 3 | 43 ± 22 | 1.6 ± 0.5 | 0.5 ± 0.2 | 136 ± 147 | 124 ± 122 | |
| DKA 4 | 99 ± 65 | 0.8 ± 0.2 | 0.2 ± 0.1 | 536 ± 563 | 384 ± 378 | |
| DKA 1 NC | 35 ± 8 | 1.8 ± 0.2 | 0.5 ± 0.1 | 44 ± 16 | 41 ± 12 | |
| DKA 2 NC | 22 ± 4 | 1.4 ± 0.5 | 0.5 ± 0.2 | 26 ± 6 | 27 ± 6 | |
| DKA 3 NC | 17 ± 1 | 1.8 ± 0.2 | 0.6 ± 0.1 | 21 ± 3 | 21 ± 3 | |
| DKA 4 NC | 29 ± 3 | 1.9 ± 0.9 | 0.6 ± 0.3 | 42 ± 8 | 41 ± 6 | |
| PC | 112 ± 31 | 1.1 ± 0.1 | 0.2 ± 0.0 | 530 ± 270 | 512 ± 205 | |
| B | cDNA | OTU Richness | Shannon | Pielou’s Evenness | S.chao1 * | S.ACE ** |
| DKA 1 | 101 ± 60 | 1.0 ± 0.3 | 0.2 ± 0.1 | 370 ± 298 | 389 ± 348 | |
| DKA 2 | 31 ± 24 | 1.3 ± 0.2 | 0.5 ± 0.2 | 44 ± 34 | 46 ± 32 | |
| DKA 3 | 44 ± 24 | 2.0 ± 0.5 | 0.6 ± 0.1 | 88 ± 65 | 78 ± 52 | |
| DKA 4 | 104 ± 144 | 1.6 ± 1.0 | 0.4 ± 0.1 | 168 ± 239 | 145 ± 194 | |
| DKA 1 NC | 12 ± 6 | 1.7 ± 0.6 | 0.8 ± 0.2 | 20 ± 10 | 28 ± 7 | |
| DKA 2 NC | 60 ± 29 | 2.7 ± 0.7 | 0.7 ± 0.1 | 75 ± 31 | 77 ± 32 | |
| DKA 3 NC | 20 ± 12 | 1.8 ± 0.6 | 0.6 ± 0.1 | 27 ± 10 | 33 ± 6 | |
| DKA 4 NC | 20 ± 8 | 1.9 ± 0.8 | 0.6 ± 0.2 | 34 ± 3 | 39 ± 2 | |
| PC | 92 ± 49 | 0.8 ± 0.3 | 0.2 ± 0.1 | 300 ± 162 | 313 ± 162 |
| A | DNA | Relative Abundance of Sequences of Bacterial Contaminants [%] | B | cDNA | Relative Abundance of Sequences of Bacterial Contaminants [%] | ||||
|---|---|---|---|---|---|---|---|---|---|
| M. luteus | P. fluorescens | E. coli | M. luteus | P. fluorescens | E. coli | ||||
| DKA 1 | 3.6 | 39.7 | 54.6 | DKA 1 | 3.4 | 64.1 | 26.2 | ||
| DKA 2 | 5.1 | 25.8 | 65.2 | DKA 2 | 6.8 | 52.2 | 22.0 | ||
| DKA 3 | 5.7 | 20.8 | 37.4 | DKA 3 | 6.6 | 17.1 | 24.0 | ||
| DKA 4 | 3.5 | 23.5 | 69.7 | DKA 4 | 2.3 | 26.4 | 49.6 | ||
| DKA 1 NC | 0.1 | 5.3 | 3.1 | DKA 1 NC | 0.0 | 0.0 | 0.0 | ||
| DKA 2 NC | 0.7 | 15.1 | 0.6 | DKA 2 NC | 0.1 | 0.8 | 0.2 | ||
| DKA 3 NC | 4.4 | 19.7 | 3.0 | DKA 3 NC | 0.5 | 0.6 | 2.1 | ||
| DKA 4 NC | 0.0 | 11.0 | 1.2 | DKA 4 NC | 4.3 | 1.3 | 1.1 | ||
| PC | 7.0 | 49.8 | 39.7 | PC | 2.4 | 43.1 | 50.8 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, A.; Fingerle, V.; Noll, M. Analysis of Tick Surface Decontamination Methods. Microorganisms 2020, 8, 987. https://doi.org/10.3390/microorganisms8070987
Hoffmann A, Fingerle V, Noll M. Analysis of Tick Surface Decontamination Methods. Microorganisms. 2020; 8(7):987. https://doi.org/10.3390/microorganisms8070987
Chicago/Turabian StyleHoffmann, Angeline, Volker Fingerle, and Matthias Noll. 2020. "Analysis of Tick Surface Decontamination Methods" Microorganisms 8, no. 7: 987. https://doi.org/10.3390/microorganisms8070987