Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii

 ,
, 
Abstract
:1. Introduction
2. AR Profiles of A. baumannii Strains
3. Short Characterization of the Molecular Mechanisms of AR
3.1. Enzymatic Mechanisms
β-lactamases

{kind=link}
| Class/Group | Enzyme | Location | References |
|---|---|---|---|
| Class A β-lactamases | CTX-M (-2, -15, -43) | C, P a | [85,86,87,88,89,90,91] |
| TEM (-1, -92, -116) | P | [92,93,94,95,96,97,98] | |
| GES (-1, -5, -11, -14, -15) | P | [99,100,101,102] | |
| PER (-1, -2, -3, -7) | C, P | [103,104,105,106] | |
| VEB (-1, -3, -7) | C | [104,107] | |
| KPC (-2, -3, -10) | – | [108,109,110] | |
| SCO-1 | P | [111] | |
| CARB (-4, -10) | C, P | [112,113] | |
| SHV (-5, -12) | C | [114,115] | |
| Class B β-lactamases | IMP (-1, -2, -4, -5, -6, -8, -10, -11, -14, -19, -24) | I | [116,117,118,119,120,121,122,123,124] |
| VIM (-1, -2, -3, -4, -6, -11) | I | [121,125,126] | |
| NDM (-1, -2, -3) | C, P | [127,128,129,130] | |
| SIM-1 | I | [131] | |
| SPM-1 | P | [68] | |
| GIM-1 | I, P | [69] | |
| FIM-1 | C | [71] | |
| Class C β-lactamases | AmpC | C | [132,133,134] |
| Class D β-lactamases | OXA-23-like (-23, -27, -49, -73, -102, -103, -105, -133, -134, -146, -165- OXA-171, -225, -239) | C, P | [35,135,136,137,138] |
| OXA-40/24-like (-40, -25, -26, -72, -139, -160, -207) | C, P | [138,139,140] | |
| OXA-51-like (-51, OXA-64– OXA-71, OXA-75– OXA-80, OXA-82- OXA-84, OXA-86– OXA-95, OXA-98– OXA-100, -104, OXA-106– OXA-113, OXA-115– OXA-117, OXA-120– OXA-128, OXA-130– OXA-132, -138, -144, OXA-148– OXA-150, OXA-172– OXA-180, OXA-194– OXA-197, OXA-200– OXA-203, -206, -208, -216, -217, -219, -223, -241, -242, OXA-248– OXA-250, -254) | C, P | [32,141,142,143,144,145,146] | |
| OXA-58-like (-58, -96, -97, -164) | C, P | [122,147,148,149] | |
| OXA-143-like (-143, -182, -231, -253, -255 | P | [150,151,152,153] | |
| OXA-48-like (-48, -48b, -162, -163, -181, -199, -204, -232, -244, -245, -247). | C, P | [154,155,156,157,158] | |
| OXA-235 | C, P | [159] |
3.2. Non-Enzymatic Mechanisms
3.2.1. Activation of the Efflux Pumps
3.2.2. Decreased Membrane Permeability
3.2.3. Changing the Target Site
4. Innovative Strategies for Treatment of A. baumannii Infections
4.1. Antimicrobial Peptides (AMP)
4.2. Bacteriophages Therapy
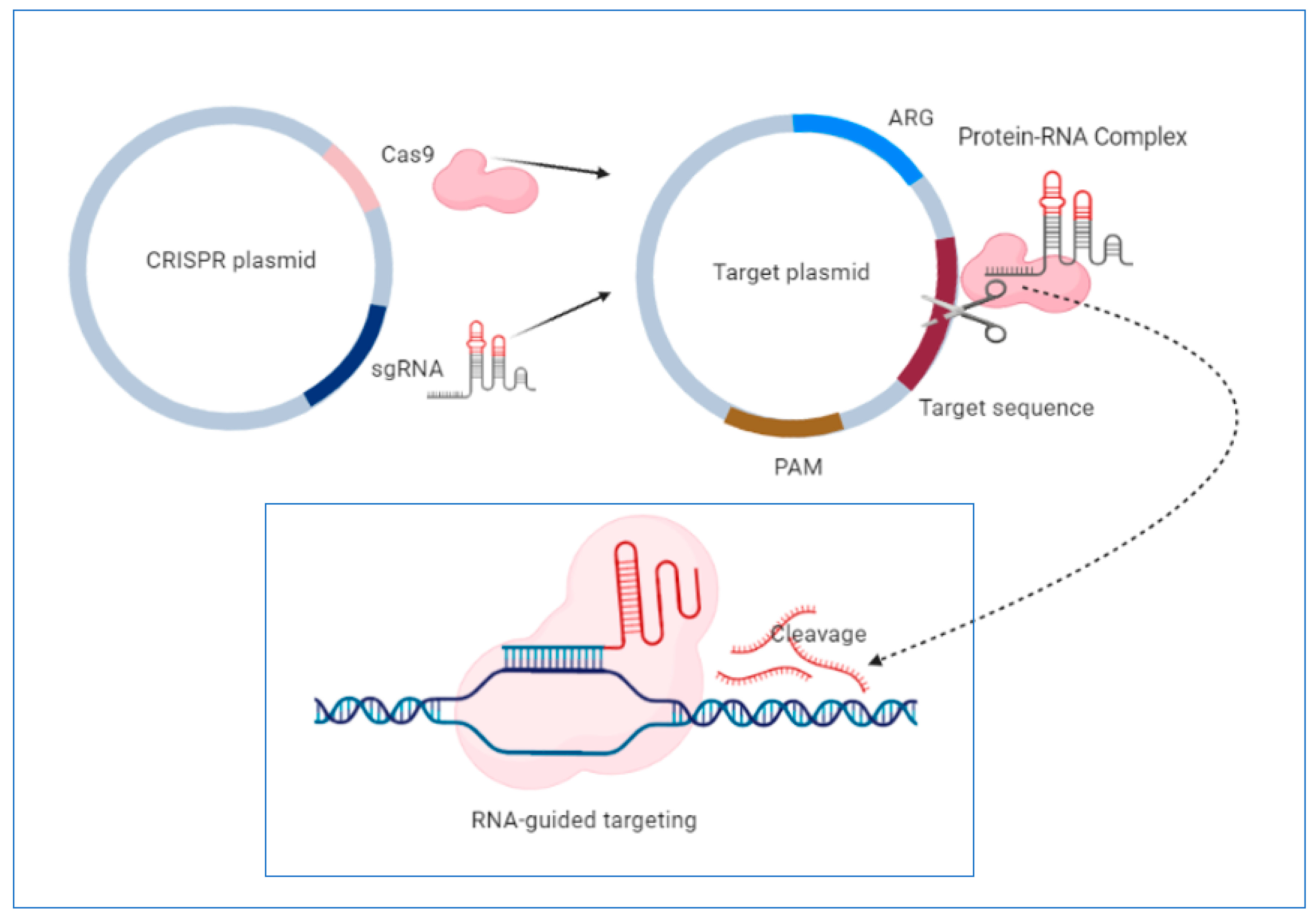
4.3. CRISPR System-a New Approach in the “Post-Antibiotic Era”?
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistance Infections Globally: Final Report and Recommendations. The Review on Antimicrobial Resistance; Government of the United Kingdom: London, UK, 2016; pp. 1–84. [Google Scholar]
- Monserrat-Martinez, A.; Gambin, Y.; Sierecki, E. Thinking Outside the Bug: Molecular Targets and Strategies to Overcome Antibiotic Resistance. Int. J. Mol. Sci. 2019, 20, 1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the Mechanisms and Drivers of Antimicrobial Resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter Baumannii Biofilms: Effects of Physicochemical Factors, Virulence, Antibiotic Resistance Determinants, Gene Regulation, and Future Antimicrobial Treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.K.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and Phenotypic Characterization of the Acinetobacter Calcoaceticus-Acinetobacter Baumannii Complex with the Proposal of Acinetobacter Pittii Sp. Nov. (Formerly Acinetobacter Genomic Species 3) and Acinetobacter Nosocomialis Sp. Nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter Seifertii Sp. Nov., a Member of the Acinetobacter Calcoaceticus-Acinetobacter Baumannii Complex Isolated from Human Clinical Specimens. Int. J. Syst. Evol. Microbiol. 2015, 65, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Cosgaya, C.; Marí-Almirall, M.; Van Assche, A.; Fernández-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter Dijkshoorniae Sp. Nov., a Member of the Acinetobacter Calcoaceticus-Acinetobacter Baumannii Complex Mainly Recovered from Clinical Samples in Different Countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef]
- Gerner-Smidt, P.; Tjernberg, I. Acinetobacter in Denmark: II. Molecular Studies of the Acinetobacter Calcoaceticus-Acinetobacter Baumannii Complex. APMIS 1993, 101, 826–832. [Google Scholar] [CrossRef]
- Maragakis, L.L.; Perl, T.M. Acinetobacter Baumannii: Epidemiology, Antimicrobial Resistance, and Treatment Options. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2008, 46, 1254–1263. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter Baumannii: Emergence of a Successful Pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An Increasing Threat in Hospitals: Multidrug-Resistant Acinetobacter Baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef]
- Willyard, C. The Drug-Resistant Bacteria That Pose the Greatest Health Threats. Nature 2017, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, V.C.J.; Rodrigues, B.Á.; Bonatto, G.D.; Gallo, S.W.; Pagnussatti, V.E.; Ferreira, C.A.S.; de Oliveira, S.D. Heterogeneous Persister Cells Formation in Acinetobacter Baumannii. PLoS ONE 2013, 8, e84361. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Khyriem, A.B.; Bhattacharya, P.; Bhattacharjee, A.; Joshi, S.R. High-Level Aminoglycoside Resistance in Acinetobacter Baumannii Recovered from Intensive Care Unit Patients in Northeastern India. Indian J. Med. Microbiol. 2018, 36, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kwon, N.Y.; Kim, J.D.; Pai, H.J. The Resistance Mechanisms of B-Lactam Antimicrobials in Clinical Isolates of Acinetobacter Baumannii. Korean J. Intern. Med. 2002, 17, 94–99. [Google Scholar] [CrossRef]
- Maniatis, A.N.; Pournaras, S.; Orkopoulou, S.; Tassios, P.T.; Legakis, N.J. Multiresistant Acinetobacter Baumannii Isolates in Intensive Care Units in Greece. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2003, 9, 547–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turton, J.F.; Kaufmann, M.E.; Warner, M.; Coelho, J.; Dijkshoorn, L.; van der Reijden, T.; Pitt, T.L. A Prevalent, Multiresistant Clone of Acinetobacter Baumannii in Southeast England. J. Hosp. Infect. 2004, 58, 170–179. [Google Scholar] [CrossRef]
- Yau, W.; Owen, R.J.; Poudyal, A.; Bell, J.M.; Turnidge, J.D.; Yu, H.H.; Nation, R.L.; Li, J. Colistin Hetero-Resistance in Multidrug-Resistant Acinetobacter Baumannii Clinical Isolates from the Western Pacific Region in the SENTRY Antimicrobial Surveillance Programme. J. Infect. 2009, 58, 138–144. [Google Scholar] [CrossRef]
- Rodríguez, C.H.; Nastro, M.; Famiglietti, A. Carbapenemases in Acinetobacter Baumannii. Review of Their Dissemination in Latin America. Rev. Argent. Microbiol. 2018, 50, 327–333. [Google Scholar] [CrossRef]
- Rodríguez, C.H.; Nastro, M.; Fiorilli, G.; Dabos, L.; Lopez Calvo, J.; Fariña, M.E.; Vay, C.; Famiglietti, A. Trends in the Resistance Profiles of Acinetobacter Baumannii Endemic Clones in a University Hospital of Argentina. J. Chemother. 2016, 28, 25–27. [Google Scholar] [CrossRef]
- D’Onofrio, V.; Conzemius, R.; Varda-Brkić, D.; Bogdan, M.; Grisold, A.; Gyssens, I.C.; Bedenić, B.; Barišić, I. Epidemiology of Colistin-Resistant, Carbapenemase-Producing Enterobacteriaceae and Acinetobacter Baumannii in Croatia. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 81, 104263. [Google Scholar] [CrossRef]
- Makke, G.; Bitar, I.; Salloum, T.; Panossian, B.; Alousi, S.; Arabaghian, H.; Medvecky, M.; Hrabak, J.; Merheb-Ghoussoub, S.; Tokajian, S. Whole-Genome-Sequence-Based Characterization of Extensively Drug-Resistant Acinetobacter Baumannii Hospital Outbreak. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammina, C.; Palma, D.M.; Bonura, C.; Aleo, A.; Fasciana, T.; Sodano, C.; Saporito, M.A.; Verde, M.S.; Calà, C.; Cracchiolo, A.N.; et al. Epidemiology and Clonality of Carbapenem-Resistant Acinetobacter Baumannii from an Intensive Care Unit in Palermo, Italy. BMC Res. Notes 2012, 5, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, J.; Zander, E.; Stefanik, D.; Higgins, P.G.; Roca, I.; Vila, J.; McConnell, M.J.; Cisneros, J.M.; Seifert, H. High Incidence of Pandrug-Resistant Acinetobacter Baumannii Isolates Collected from Patients with Ventilator-Associated Pneumonia in Greece, Italy and Spain as Part of the MagicBullet Clinical Trial. J. Antimicrob. Chemother. 2017, 72, 3277–3282. [Google Scholar] [CrossRef] [PubMed]
- Dahdouh, E.; Gómez-Gil, R.; Pacho, S.; Mingorance, J.; Daoud, Z.; Suárez, M. Clonality, Virulence Determinants, and Profiles of Resistance of Clinical Acinetobacter Baumannii Isolates Obtained from a Spanish Hospital. PLoS ONE 2017, 12, e0176824. [Google Scholar] [CrossRef]
- Simo Tchuinte, P.L.; Rabenandrasana, M.A.N.; Kowalewicz, C.; Andrianoelina, V.H.; Rakotondrasoa, A.; Andrianirina, Z.Z.; Enouf, V.; Ratsima, E.H.; Randrianirina, F.; Collard, J.-M. Phenotypic and Molecular Characterisations of Carbapenem-Resistant Acinetobacter Baumannii Strains Isolated in Madagascar. Antimicrob. Resist. Infect. Control 2019, 8, 31. [Google Scholar] [CrossRef]
- Shamsizadeh, Z.; Nikaeen, M.; Nasr Esfahani, B.; Mirhoseini, S.H.; Hatamzadeh, M.; Hassanzadeh, A. Detection of Antibiotic Resistant Acinetobacter Baumannii in Various Hospital Environments: Potential Sources for Transmission of Acinetobacter Infections. Environ. Health Prev. Med. 2017, 22, 44. [Google Scholar] [CrossRef]
- da Silva, K.E.; Maciel, W.G.; Croda, J.; Cayô, R.; Ramos, A.C.; de Sales, R.O.; Kurihara, M.N.L.; Vasconcelos, N.G.; Gales, A.C.; Simionatto, S. A High Mortality Rate Associated with Multidrug-Resistant Acinetobacter Baumannii ST79 and ST25 Carrying OXA-23 in a Brazilian Intensive Care Unit. PLoS ONE 2018, 13, e0209367. [Google Scholar] [CrossRef] [Green Version]
- Arhoune, B.; Oumokhtar, B.; Hmami, F.; El Fakir, S.; Moutaouakkil, K.; Chami, F.; Bouharrou, A. Intestinal Carriage of Antibiotic Resistant Acinetobacter Baumannii among Newborns Hospitalized in Moroccan Neonatal Intensive Care Unit. PLoS ONE 2019, 14, e0209425. [Google Scholar] [CrossRef]
- Tafreshi, N.; Babaeekhou, L.; Ghane, M. Antibiotic Resistance Pattern of Acinetobacter Baumannii from Burns Patients: Increase in Prevalence of Bla (OXA-24-like) and Bla (OXA-58-like) Genes. Iran. J. Microbiol. 2019, 11, 502–509. [Google Scholar] [CrossRef]
- Araújo Lima, A.V.; da Silva, S.M.; do Nascimento Júnior, J.A.A.; Correia, M.D.S.; Luz, A.C.; Leal-Balbino, T.C.; da Silva, M.V.; Lima, J.L.D.C.; Maciel, M.A.V.; Napoleão, T.H.; et al. Occurrence and Diversity of Intra- and Interhospital Drug-Resistant and Biofilm-Forming Acinetobacter Baumannii and Pseudomonas Aeruginosa. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef]
- Hu, W.S.; Yao, S.-M.; Fung, C.-P.; Hsieh, Y.-P.; Liu, C.-P.; Lin, J.-F. An OXA-66/OXA-51-like Carbapenemase and Possibly an Efflux Pump Are Associated with Resistance to Imipenem in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2007, 51, 3844–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammina, C.; Bonura, C.; Aleo, A.; Calà, C.; Caputo, G.; Cataldo, M.C.; Di Benedetto, A.; Distefano, S.; Fasciana, T.; Labisi, M.; et al. Characterization of Acinetobacter Baumannii from Intensive Care Units and Home Care Patients in Palermo, Italy. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2011, 17, E12–E15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajand, O.; Hojabri, Z.; Nahaei, M.R.; Hajibonabi, F.; Pirzadeh, T.; Aghazadeh, M.; Fasciana, T.; Bonura, C.; Mammina, C. In Vitro Activities of Tetracyclines against Different Clones of Multidrug-Resistant Acinetobacter Baumannii Isolates from Two Iranian Hospitals. Int. J. Antimicrob. Agents 2014, 476–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.E.; Bell, J.M.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Emergence and Widespread Dissemination of OXA-23, -24/40 and -58 Carbapenemases among Acinetobacter spp. in Asia-Pacific Nations: Report from the SENTRY Surveillance Program. J. Antimicrob. Chemother. 2009, 63, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Vahaboglu, H.; Budak, F.; Kasap, M.; Gacar, G.; Torol, S.; Karadenizli, A.; Kolayli, F.; Eroglu, C. High Prevalence of OXA-51-Type Class D Beta-Lactamases among Ceftazidime-Resistant Clinical Isolates of Acinetobacter spp.: Co-Existence with OXA-58 in Multiple Centres. J. Antimicrob. Chemother. 2006, 58, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Lolans, K.; Rice, T.W.; Munoz-Price, L.S.; Quinn, J.P. Multicity Outbreak of Carbapenem-Resistant Acinetobacter Baumannii Isolates Producing the Carbapenemase OXA-40. Antimicrob. Agents Chemother. 2006, 50, 2941–2945. [Google Scholar] [CrossRef] [Green Version]
- Mugnier, P.D.; Poirel, L.; Naas, T.; Nordmann, P. Worldwide Dissemination of the BlaOXA-23 Carbapenemase Gene of Acinetobacter Baumannii. Emerg. Infect. Dis. 2010, 16, 35–40. [Google Scholar] [CrossRef]
- Chagas, T.P.G.; Carvalho, K.R.; de Oliveira Santos, I.C.; Carvalho-Assef, A.P.D.; Asensi, M.D. Characterization of Carbapenem-Resistant Acinetobacter Baumannii in Brazil (2008–2011): Countrywide Spread of OXA-23-Producing Clones (CC15 and CC79). Diagn. Microbiol. Infect. Dis. 2014, 79, 468–472. [Google Scholar] [CrossRef] [Green Version]
- Pagano, M.; Martins, A.F.; Machado, A.B.M.P.; Barin, J.; Barth, A.L. Carbapenem-Susceptible Acinetobacter Baumannii Carrying the ISAba1 Upstream BlaOXA-51-like Gene in Porto Alegre, Southern Brazil. Epidemiol. Infect. 2013, 141, 330–333. [Google Scholar] [CrossRef]
- Ben Othman, A.; Zribi, M.; Masmoudi, A.; Abdellatif, S.; Ben Lakhal, S.; Fendri, C. Multiresistance and Endemic Status of Acinetobacter Baumannii Associated with Nosocomial Infections in a Tunisian Hospital: A Critical Situation in the Intensive Care Units. Braz. J. Microbiol. 2011, 42, 415–422. [Google Scholar] [CrossRef]
- Akbari, M.; Niakan, M.; Taherikalani, M.; Feizabadi, M.M.; Azadi, N.A.; Soroush, S.; Emaneini, M.; Abdolkarimi, A.; Maleki, A.; Hematian, A. Rapid Identification of Iranian Acinetobacter Baumannii Strains by Single PCR Assay Using BLA Oxa-51 -like Carbapenemase and Evaluation of the Antimicrobial Resistance Profiles of the Isolates. Acta Microbiol. Immunol. Hung. 2010, 57, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Çiftci, I.H.; Aşik, G.; Karakeçe, E.; Öksüz, L.; Yaǧci, S.; Sesli Çetin, E.; Özdemir, M.; Atasoy, A.R.; Koçoǧlu, E.; Gül, M.; et al. Distribution of Bla OXA Genes in Acinetobacter Baumannii Strains: A Multicenter Study. Mikrobiyol. Bul. 2013, 47, 592–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, B.S.; Gallego, L.; Amyes, S.G.B. Multi-Drug Resistance Profiles and the Genetic Features of Acinetobacter Baumannii Isolates from Bolivia. J. Infect. Dev. Ctries. 2013, 7, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rammelkamp, C.H.; Maxon, T. Resistance of Staphylococcus Aureus to the Action of Penicillin. Proc. Soc. Exp. Biol. Med. 1942, 51, 386–389. [Google Scholar] [CrossRef]
- Sultan, I.; Rahman, S.; Jan, A.T.; Siddiqui, M.T.; Mondal, A.H.; Haq, Q.M.R. Antibiotics, Resistome and Resistance Mechanisms: A Bacterial Perspective. Front. Microbiol. 2018, 9, 2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.L.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic Resistance Is Ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef]
- Wright, G.D. Molecular Mechanisms of Antibiotic Resistance. Chem. Commun. 2011, 47, 4055–4061. [Google Scholar] [CrossRef]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria Able to Destroy Penicillin. 1940. Rev. Infect. Dis. 1988, 10, 677–678. [Google Scholar]
- Hall, B.G.; Barlow, M. Evolution of the Serine Beta-Lactamases: Past, Present and Future. Drug Resist. Updat. Rev. Comment. Antimicrob. Anticancer Chemother. 2004, 7, 111–123. [Google Scholar] [CrossRef]
- Barlow, M.; Hall, B.G. Phylogenetic Analysis Shows That the OXA Beta-Lactamase Genes Have Been on Plasmids for Millions of Years. J. Mol. Evol. 2002, 55, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Frère, J.M.; Duez, C.; Ghuysen, J.M.; Vandekerkhove, J. Occurrence of a Serine Residue in the Penicillin-Binding Site of the Exocellular DD-Carboxy-Peptidase-Transpeptidase from Streptomyces R61. FEBS Lett. 1976, 70, 257–260. [Google Scholar] [CrossRef] [Green Version]
- Massova, I.; Mobashery, S. Kinship and Diversification of Bacterial Penicillin-Binding Proteins and Beta-Lactamases. Antimicrob. Agents Chemother. 1998, 42, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, J.F.; Mobashery, S. β-Lactam Resistance Mechanisms: Gram-Positive Bacteria and Mycobacterium Tuberculosis. Cold Spring Harb. Perspect. Med. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Matthew, M.; Harris, A.M. Identification of Beta-Lactamases by Analytical Isoelectric Focusing: Correlation with Bacterial Taxonomy. J. Gen. Microbiol. 1976, 94, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Ambler, R.P. The Structure of Beta-Lactamases. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A.; Medeiros, A.A. A Functional Classification Scheme for Beta-Lactamases and Its Correlation with Molecular Structure. Antimicrob. Agents Chemother. 1995, 39, 1211–1233. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of Beta-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Bush, K. Proliferation and Significance of Clinically Relevant β-Lactamases. Ann. N. Y. Acad. Sci. 2013, 1277, 84–90. [Google Scholar] [CrossRef]
- Jeon, J.H.; Lee, J.H.; Lee, J.J.; Park, K.S.; Karim, A.M.; Lee, C.-R.; Jeong, B.C.; Lee, S.H. Structural Basis for Carbapenem-Hydrolyzing Mechanisms of Carbapenemases Conferring Antibiotic Resistance. Int. J. Mol. Sci. 2015, 16, 9654–9692. [Google Scholar] [CrossRef]
- Naas, T.; Dortet, L.; Iorga, B.I. Structural and Functional Aspects of Class A Carbapenemases. Curr. Drug Targets 2016, 17, 1006–1028. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. The ABCD’s of β-Lactamase Nomenclature. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2013, 19, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Palzkill, T. Metallo-β-Lactamase Structure and Function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Aoki, N.; Ishii, Y.; Tateda, K.; Saga, T.; Kimura, S.; Kikuchi, Y.; Kobayashi, T.; Tanabe, Y.; Tsukada, H.; Gejyo, F.; et al. Efficacy of Calcium-EDTA as an Inhibitor for Metallo-β-Lactamase in a Mouse Model of Pseudomonas Aeruginosa Pneumonia. Antimicrob. Agents Chemother. 2010, 54, 4582–4588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, Y.; Murakami, M.; Suzuki, K.; Ito, H.; Wacharotayankun, R.; Ohsuka, S.; Kato, N.; Ohta, M. A Novel Integron-like Element Carrying the Metallo-Beta-Lactamase Gene BlaIMP. Antimicrob. Agents Chemother. 1995, 39, 1612–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauretti, L.; Riccio, M.L.; Mazzariol, A.; Cornaglia, G.; Amicosante, G.; Fontana, R.; Rossolini, G.M. Cloning and Characterization of BlaVIM, a New Integron-Borne Metallo-Beta-Lactamase Gene from a Pseudomonas Aeruginosa Clinical Isolate. Antimicrob. Agents Chemother. 1999, 43, 1584–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleman, M.A.; Simm, A.M.; Murphy, T.A.; Gales, A.C.; Biedenbach, D.J.; Jones, R.N.; Walsh, T.R. Molecular Characterization of SPM-1, a Novel Metallo-β-Lactamase Isolated in Latin America: Report from the SENTRY Antimicrobial Surveillance Programme. J. Antimicrob. Chemother. 2002, 50, 673–679. [Google Scholar] [CrossRef]
- Castanheira, M.; Toleman, M.A.; Jones, R.N.; Schmidt, F.J.; Walsh, T.R. Molecular Characterization of a Beta-Lactamase Gene, BlaGIM-1, Encoding a New Subclass of Metallo-Beta-Lactamase. Antimicrob. Agents Chemother. 2004, 48, 4654–4661. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a New Metallo-Beta-Lactamase Gene, Bla(NDM-1), and a Novel Erythromycin Esterase Gene Carried on a Unique Genetic Structure in Klebsiella Pneumoniae Sequence Type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Pollini, S.; Maradei, S.; Pecile, P.; Olivo, G.; Luzzaro, F.; Docquier, J.-D.; Rossolini, G.M. FIM-1, a New Acquired Metallo-β-Lactamase from a Pseudomonas Aeruginosa Clinical Isolate from Italy. Antimicrob. Agents Chemother. 2013, 57, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. AmpC Beta-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, K.S. Extended-Spectrum-Beta-Lactamase, AmpC, and Carbapenemase Issues. J. Clin. Microbiol. 2010, 48, 1019–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, N.C.; Wareham, D.W. Multidrug-Resistant Acinetobacter Baumannii: Mechanisms of Virulence and Resistance. Int. J. Antimicrob. Agents 2010, 35, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liu, B.; Chen, Y.; Huang, H.; Wang, G.; Li, F.; Ni, Z. Molecular Characterization of Ambler Class A to D β-Lactamases, ISAba1, and Integrons Reveals Multidrug-Resistant Acinetobacter spp. Isolates in Northeastern China. J. Chemother. 2016, 28, 469–475. [Google Scholar] [CrossRef]
- Lin, M.-F.; Chang, K.-C.; Lan, C.-Y.; Chou, J.; Kuo, J.-W.; Chang, C.-K.; Liou, M.-L. Molecular Epidemiology and Antimicrobial Resistance Determinants of Multidrug-Resistant Acinetobacter Baumannii in Five Proximal Hospitals in Taiwan. Jpn. J. Infect. Dis. 2011, 64, 222–227. [Google Scholar]
- Lin, M.-F.; Lan, C.-Y. Antimicrobial Resistance in Acinetobacter Baumannii: From Bench to Bedside. World J. Clin. Cases 2014, 2, 787–814. [Google Scholar] [CrossRef]
- Nowak, P.; Paluchowska, P. Acinetobacter Baumannii: Biology and Drug Resistance—Role of Carbapenemases. Folia Histochem. Cytobiol. 2016, 54, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Lyon, J.A. Imipenem/Cilastatin: The First Carbapenem Antibiotic. Drug Intell. Clin. Pharm. 1985, 19, 895–899. [Google Scholar]
- Périchon, B.; Goussard, S.; Walewski, V.; Krizova, L.; Cerqueira, G.; Murphy, C.; Feldgarden, M.; Wortman, J.; Clermont, D.; Nemec, A.; et al. Identification of 50 Class D β-Lactamases and 65 Acinetobacter-Derived Cephalosporinases in Acinetobacter spp. Antimicrob. Agents Chemother. 2014, 58, 936–949. [Google Scholar] [CrossRef] [Green Version]
- Al Atrouni, A.; Hamze, M.; Jisr, T.; Lemarié, C.; Eveillard, M.; Joly-Guillou, M.-L.; Kempf, M. Wide Spread of OXA-23-Producing Carbapenem-Resistant Acinetobacter Baumannii Belonging to Clonal Complex II in Different Hospitals in Lebanon. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2016, 52, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Davandeh, I.; Eraç, B.; Aydemir, S.Ş. Investigation of Class-d Beta-Lactamases Causing Carbapenem Resistance in Clinical Acinetobacter Baumannii Isolates. Turk. J. Med. Sci. 2017, 47, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donald, H.M.; Scaife, W.; Amyes, S.G.; Young, H.K. Sequence Analysis of ARI-1, a Novel OXA Beta-Lactamase, Responsible for Imipenem Resistance in Acinetobacter Baumannii 6B92. Antimicrob. Agents Chemother. 2000, 44, 196–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potron, A.; Munoz-Price, L.S.; Nordmann, P.; Cleary, T.; Poirel, L. Genetic Features of CTX-M-15-Producing Acinetobacter Baumannii from Haiti. Antimicrob. Agents Chemother. 2011, 55, 5946–5948. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Malik, A.; Rizvi, M.; Ahmed, M. Presence of Metallo-Beta-Lactamases (MBL), Extended-Spectrum Beta-Lactamase (ESBL) & AmpC Positive Non-Fermenting Gram-Negative Bacilli among Intensive Care Unit Patients with Special Reference to Molecular Detection of Bla(CTX-M) & Bla(AmpC) Genes. Indian J. Med. Res. 2016, 144, 271–275. [Google Scholar] [CrossRef]
- Beriş, F.Ş.; Budak, E.E.; Gülek, D.; Uzun, A.; Çizmeci, Z.; Mengeloğlu, F.Z.; Direkel, Ş.; Çetinkol, Y.; Ay Altıntop, Y.; Iraz, M.; et al. [Investigation of the frequency and distribution of beta-lactamase genes in the clinical isolates of Acinetobacter baumannii collected from different regions of Turkey: A multicenter study]. Mikrobiyol. Bull. 2016, 50, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Freitas, D.Y.; Araújo, S.; Folador, A.R.C.; Ramos, R.T.J.; Azevedo, J.S.N.; Tacão, M.; Silva, A.; Henriques, I.; Baraúna, R.A. Extended Spectrum Beta-Lactamase-Producing Gram-Negative Bacteria Recovered From an Amazonian Lake Near the City of Belém, Brazil. Front. Microbiol. 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Alyamani, E.J.; Khiyami, M.A.; Booq, R.Y.; Alnafjan, B.M.; Altammami, M.A.; Bahwerth, F.S. Molecular Characterization of Extended-Spectrum Beta-Lactamases (ESBLs) Produced by Clinical Isolates of Acinetobacter Baumannii in Saudi Arabia. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 38. [Google Scholar] [CrossRef] [Green Version]
- Mayanskiy, N.; Chebotar, I.; Alyabieva, N.; Kryzhanovskaya, O.; Savinova, T.; Turenok, A.; Bocharova, Y.; Lazareva, A.; Polikarpova, S.; Karaseva, O. Emergence of the Uncommon Clone ST944/ST78 Carrying Bla(OXA-40-like) and Bla(CTX-M-like) Genes Among Carbapenem-Nonsusceptible Acinetobacter Baumannii in Moscow, Russia. Microb. Drug Resist. 2017, 23, 864–870. [Google Scholar] [CrossRef]
- Zago, M.C.B.; Viana, G.F.; Ecker, A.B.S.; Nishiyama, S.A.B.; Zarpellon, M.N.; Dias, J.R.C.; Cardoso, C.L.; Tognim, M.C.B. First Report of CTX-M-15-Producing Acinetobacter Baumannii in Brazil. J. Hosp. Infect. 2016, 298–299. [Google Scholar] [CrossRef]
- Krizova, L.; Poirel, L.; Nordmann, P.; Nemec, A. TEM-1 β-Lactamase as a Source of Resistance to Sulbactam in Clinical Strains of Acinetobacter Baumannii. J. Antimicrob. Chemother. 2013, 68, 2786–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ling, B.; Zhou, L. Prevalence of 16S RRNA Methylase, Modifying Enzyme, and Extended-Spectrum Beta-Lactamase Genes among Acinetobacter Baumannii Isolates. J. Chemother. 2015, 27, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Ibrahimagić, A.; Kamberović, F.; Uzunović, S.; Bedenić, B.; Idrizović, E. Molecular Characteristics and Antibiotic Resistance of Acinetobacter Baumanniibeta-Lactamase-Producing Isolates, a Predominance of Intrinsic BlaOXA-51, and Detection of TEM and CTX-M Genes. Turk. J. Med. Sci. 2017, 47, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Yu, P.; Ge, P.; Jiang, Y.; Xu, R.; Chen, R.; Liu, X. Identification of Antibiotic Resistance Genes in the Multidrug-Resistant Acinetobacter Baumannii Strain, MDR-SHH02, Using Whole-Genome Sequencing. Int. J. Mol. Med. 2017, 39, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Abdar, M.H.; Taheri-Kalani, M.; Taheri, K.; Emadi, B.; Hasanzadeh, A.; Sedighi, A.; Pirouzi, S.; Sedighi, M. Prevalence of Extended-Spectrum Beta-Lactamase Genes in Acinetobacter Baumannii Strains Isolated from Nosocomial Infections in Tehran, Iran. GMS Hyg. Infect. Control 2019, 14, Doc02. [Google Scholar] [CrossRef]
- Asgin, N.; Otlu, B.; Cakmakliogullari, E.K.; Celik, B. High Prevalence of TEM, VIM, and OXA-2 Beta-Lactamases and Clonal Diversity among Acinetobacter Baumannii Isolates in Turkey. J. Infect. Dev. Ctries. 2019, 13, 794–801. [Google Scholar] [CrossRef] [Green Version]
- Agoba, E.E.; Govinden, U.; Peer, A.K.C.; Osei Sekyere, J.; Essack, S.Y. ISAba1 Regulated OXA-23 Carbapenem Resistance in Acinetobacter Baumannii Strains in Durban, South Africa. Microb. Drug Resist. 2018, 24, 1289–1295. [Google Scholar] [CrossRef]
- Al-Agamy, M.H.; Jeannot, K.; El-Mahdy, T.S.; Shibl, A.M.; Kattan, W.; Plésiat, P.; Courvalin, P. First Detection of GES-5 Carbapenemase-Producing Acinetobacter Baumannii Isolate. Microb. Drug Resist. 2017, 23, 556–562. [Google Scholar] [CrossRef]
- Chihi, H.; Bonnin, R.A.; Bourouis, A.; Mahrouki, S.; Besbes, S.; Moussa, M.B.; Belhadj, O.; Naas, T. GES-11-Producing Acinetobacter Baumannii Clinical Isolates from Tunisian Hospitals: Long-Term Dissemination of GES-Type Carbapenemases in North Africa. J. Glob. Antimicrob. Resist. 2016, 5, 47–50. [Google Scholar] [CrossRef]
- Mabrouk, A.; Grosso, F.; Botelho, J.; Achour, W.; Ben Hassen, A.; Peixe, L. GES-14-Producing Acinetobacter Baumannii Isolates in a Neonatal Intensive Care Unit in Tunisia Are Associated with a Typical Middle East Clone and a Transferable Plasmid. Antimicrob. Agents Chemother. 2017. [Google Scholar] [CrossRef] [Green Version]
- El-Shazly, S.; Dashti, A.; Vali, L.; Bolaris, M.; Ibrahim, A.S. Molecular Epidemiology and Characterization of Multiple Drug-Resistant (MDR) Clinical Isolates of Acinetobacter Baumannii. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2015, 41, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, M.M.; Abu Alsoud, N.M.; Elrobh, M.S.; Al Johani, S.M.; Balkhy, H.H. High Prevalence of the PER-1 Gene among Carbapenem-Resistant Acinetobacter Baumannii in Riyadh, Saudi Arabia. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2016, 35, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Pasterán, F.; Rapoport, M.; Petroni, A.; Faccone, D.; Corso, A.; Galas, M.; Vázquez, M.; Procopio, A.; Tokumoto, M.; Cagnoni, V. Emergence of PER-2 and VEB-1a in Acinetobacter Baumannii Strains in the Americas. Antimicrob. Agents Chemother. 2006, 3222–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnin, R.A.; Potron, A.; Poirel, L.; Lecuyer, H.; Neri, R.; Nordmann, P. PER-7, an Extended-Spectrum Beta-Lactamase with Increased Activity toward Broad-Spectrum Cephalosporins in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2011, 55, 2424–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hassan, L.; El Mahallawy, H.; Amyes, S.G.B. First Report of Bla(PER-3) in Acinetobacter Baumannii. Int. J. Antimicrob. Agents. 2013, 93–94. [Google Scholar] [CrossRef]
- Huang, L.-Y.; Chen, T.-L.; Lu, P.-L.; Tsai, C.-A.; Cho, W.-L.; Chang, F.-Y.; Fung, C.-P.; Siu, L.K. Dissemination of Multidrug-Resistant, Class 1 Integron-Carrying Acinetobacter Baumannii Isolates in Taiwan. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2008, 14, 1010–1019. [Google Scholar] [CrossRef] [Green Version]
- Azimi, L.; Talebi, M.; Pourshafie, M.-R.; Owlia, P.; Rastegar Lari, A. Characterization of Carbapenemases in Extensively Drug Resistance Acinetobacter Baumannii in a Burn Care Center in Iran. Int. J. Mol. Cell. Med. 2015, 4, 46–53. [Google Scholar]
- Robledo, I.E.; Aquino, E.E.; Santé, M.I.; Santana, J.L.; Otero, D.M.; León, C.F.; Vázquez, G.J. Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob. Agents Chemother. 2010, 54, 1354–1357. [Google Scholar] [CrossRef] [Green Version]
- Martinez, T.; Martinez, I.; Vazquez, G.J.; Aquino, E.E.; Robledo, I.E. Genetic Environment of the KPC Gene in Acinetobacter Baumannii ST2 Clone from Puerto Rico and Genomic Insights into Its Drug Resistance. J. Med. Microbiol. 2016, 65, 784–792. [Google Scholar] [CrossRef]
- Poirel, L.; Corvec, S.; Rapoport, M.; Mugnier, P.; Petroni, A.; Pasteran, F.; Faccone, D.; Galas, M.; Drugeon, H.; Cattoir, V.; et al. Identification of the Novel Narrow-Spectrum Beta-Lactamase SCO-1 in Acinetobacter spp. from Argentina. Antimicrob. Agents Chemother. 2007, 51, 2179–2184. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Poirel, L.; Croizé, J.; Chanteperdrix, V.; Nordmann, P. Genetic and Biochemical Characterization of the First Extended-Spectrum CARB-Type Beta-Lactamase, RTG-4, from Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2009, 53, 3010–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.S.; Piñeiro, S.; Centrón, D. Novel Insights about Class 2 Integrons from Experimental and Genomic Epidemiology. Antimicrob. Agents Chemother. 2010, 54, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, T.; Namdari, F.; Réglier-Poupet, H.; Poyart, C.; Nordmann, P. Panresistant Extended-Spectrum Beta-Lactamase SHV-5-Producing Acinetobacter Baumannii from New York City. J. Antimicrob. Chemother. 2007, 1174–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Mao, P.; Chen, Y.; Wu, L.; Wu, J. Study on the molecular epidemiology of SHV type beta-lactamase-encoding genes of multiple-drug-resistant acinetobacter baumannii. Zhonghua Liu Xing Bing Xue Za Zhi 2004, 25, 425–427. [Google Scholar]
- Potron, A.; Poirel, L.; Nordmann, P. Emerging Broad-Spectrum Resistance in Pseudomonas Aeruginosa and Acinetobacter Baumannii: Mechanisms and Epidemiology. Int. J. Antimicrob. Agents 2015, 45, 568–585. [Google Scholar] [CrossRef] [Green Version]
- Gales, A.C.; Tognim, M.C.B.; Reis, A.O.; Jones, R.N.; Sader, H.S. Emergence of an IMP-like Metallo-Enzyme in an Acinetobacter Baumannii Clinical Strain from a Brazilian Teaching Hospital. Diagn. Microbiol. Infect. Dis. 2003, 45, 77–79. [Google Scholar] [CrossRef]
- Riccio, M.L.; Franceschini, N.; Boschi, L.; Caravelli, B.; Cornaglia, G.; Fontana, R.; Amicosante, G.; Rossolini, G.M. Characterization of the Metallo-Beta-Lactamase Determinant of Acinetobacter Baumannii AC-54/97 Reveals the Existence of Bla(IMP) Allelic Variants Carried by Gene Cassettes of Different Phylogeny. Antimicrob. Agents Chemother. 2000, 44, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.W.; Afzal-Shah, M.; Houang, E.T.; Palepou, M.I.; Lyon, D.J.; Woodford, N.; Livermore, D.M. IMP-4, a Novel Metallo-Beta-Lactamase from Nosocomial Acinetobacter spp. Collected in Hong Kong between 1994 and 1998. Antimicrob. Agents Chemother. 2001, 45, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Interspecies Dissemination of a Novel Class 1 Integron Carrying BlaIMP-19 among Acinetobacter Species in Japan. J. Antimicrob. Chemother. 2011, 66, 2480–2483. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-F.; Peng, C.-F.; Hsu, H.-J.; Chen, Y.-H. Molecular Characterisation of the Metallo-Beta-Lactamase Genes in Imipenem-Resistant Gram-Negative Bacteria from a University Hospital in Southern Taiwan. Int. J. Antimicrob. Agents 2008, 32, 475–480. [Google Scholar] [CrossRef]
- Koh, T.H.; Sng, L.-H.; Wang, G.C.Y.; Hsu, L.-Y.; Zhao, Y. IMP-4 and OXA Beta-Lactamases in Acinetobacter Baumannii from Singapore. J. Antimicrob. Chemother. 2007, 59, 627–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognim, M.C.B.; Gales, A.C.; Penteado, A.P.; Silbert, S.; Sader, H.S. Dissemination of IMP-1 Metallo- Beta -Lactamase-Producing Acinetobacter Species in a Brazilian Teaching Hospital. Infect. Control Hosp. Epidemiol. 2006, 27, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Cayô, R.; Rodrigues-Costa, F.; Matos, A.P.; Carvalhaes, C.G.; Jové, T.; Gales, A.C. Identification of a New Integron Harboring Bla(IMP-10) in Carbapenem-Resistant Acinetobacter Baumannii Clinical Isolates. Antimicrob. Agents Chemother. 2015, 3687–3689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, A.; Koulourida, V.; Souliou, E. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter Baumannii in a Newly Established Greek Hospital. Microb. Drug Resist. 2009, 15, 257–260. [Google Scholar] [CrossRef]
- Tsakris, A.; Ikonomidis, A.; Pournaras, S.; Tzouvelekis, L.S.; Sofianou, D.; Legakis, N.J.; Maniatis, A.N. VIM-1 Metallo-Beta-Lactamase in Acinetobacter Baumannii. Emerg. Infect. Dis. 2006, 12, 981–983. [Google Scholar] [CrossRef] [Green Version]
- Bonnin, R.A.; Poirel, L.; Naas, T.; Pirs, M.; Seme, K.; Schrenzel, J.; Nordmann, P. Dissemination of New Delhi Metallo-β-Lactamase-1-Producing Acinetobacter Baumannii in Europe. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2012, 18, E362-5. [Google Scholar] [CrossRef] [Green Version]
- Voulgari, E.; Politi, L.; Pitiriga, V.; Dendrinos, J.; Poulou, A.; Georgiadis, G.; Tsakris, A. First Report of an NDM-1 Metallo-β-Lactamase-Producing Acinetobacter Baumannii Clinical Isolate in Greece. Int. J. Antimicrob. Agents. 2016, 761–762. [Google Scholar] [CrossRef]
- Espinal, P.; Fugazza, G.; López, Y.; Kasma, M.; Lerman, Y.; Malhotra-Kumar, S.; Goossens, H.; Carmeli, Y.; Vila, J. Dissemination of an NDM-2-Producing Acinetobacter Baumannii Clone in an Israeli Rehabilitation Center. Antimicrob. Agents Chemother. 2011, 55, 5396–5398. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M. Identification of a Novel NDM Variant, BlaNDM-3, From a Multidrug-Resistant Acinetobacter Baumannii. Infect. Control Hosp. Epidemiol. 2016, 747–748. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Yum, J.H.; Yong, D.; Lee, H.M.; Kim, H.D.; Docquier, J.-D.; Rossolini, G.M.; Chong, Y. Novel Acquired Metallo-Beta-Lactamase Gene, Bla(SIM-1), in a Class 1 Integron from Acinetobacter Baumannii Clinical Isolates from Korea. Antimicrob. Agents Chemother. 2005, 49, 4485–4491. [Google Scholar] [CrossRef] [Green Version]
- Segal, H.; Nelson, E.C.; Elisha, B.G. Genetic Environment and Transcription of AmpC in an Acinetobacter Baumannii Clinical Isolate. Antimicrob. Agents Chemother. 2004, 48, 612–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hujer, K.M.; Hamza, N.S.; Hujer, A.M.; Perez, F.; Helfand, M.S.; Bethel, C.R.; Thomson, J.M.; Anderson, V.E.; Barlow, M.; Rice, L.B.; et al. Identification of a New Allelic Variant of the Acinetobacter Baumannii Cephalosporinase, ADC-7 Beta-Lactamase: Defining a Unique Family of Class C Enzymes. Antimicrob. Agents Chemother. 2005, 49, 2941–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, X. Detection of AmpC β-Lactamases in Acinetobacter Baumannii in the Xuzhou Region and Analysis of Drug Resistance. Exp. Ther. Med. 2015, 10, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Figueiredo, S.; Cattoir, V.; Carattoli, A.; Nordmann, P. Acinetobacter Radioresistens as a Silent Source of Carbapenem Resistance for Acinetobacter spp. Antimicrob. Agents Chemother. 2008, 52, 1252–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, T.W.; Crowley, B. Detection of BlaOXA-58 and BlaOXA-23-like Genes in Carbapenem-Susceptible Acinetobacter Clinical Isolates: Should We Be Concerned? J. Med. Microbiol. 2009, 839–841. [Google Scholar] [CrossRef] [Green Version]
- Kaitany, K.-C.J.; Klinger, N.V.; June, C.M.; Ramey, M.E.; Bonomo, R.A.; Powers, R.A.; Leonard, D.A. Structures of the Class D Carbapenemases OXA-23 and OXA-146: Mechanistic Basis of Activity against Carbapenems, Extended-Spectrum Cephalosporins, and Aztreonam. Antimicrob. Agents Chemother. 2013, 57, 4848–4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal-Shah, M.; Woodford, N.; Livermore, D.M. Characterization of OXA-25, OXA-26, and OXA-27, Molecular Class D Beta-Lactamases Associated with Carbapenem Resistance in Clinical Isolates of Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2001, 45, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou, G.; Oliver, A.; Martínez-Beltrán, J. OXA-24, a Novel Class D Beta-Lactamase with Carbapenemase Activity in an Acinetobacter Baumannii Clinical Strain. Antimicrob. Agents Chemother. 2000, 44, 1556–1561. [Google Scholar] [CrossRef] [Green Version]
- Santillana, E.; Beceiro, A.; Bou, G.; Romero, A. Crystal Structure of the Carbapenemase OXA-24 Reveals Insights into the Mechanism of Carbapenem Hydrolysis. Proc. Natl. Acad. Sci. USA 2007, 104, 5354–5359. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Young, H.K.; Amyes, S.G.B. Characterisation of OXA-51, a Novel Class D Carbapenemase Found in Genetically Unrelated Clinical Strains of Acinetobacter Baumannii from Argentina. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2005, 11, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Héritier, C.; Poirel, L.; Fournier, P.-E.; Claverie, J.-M.; Raoult, D.; Nordmann, P. Characterization of the Naturally Occurring Oxacillinase of Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2005, 49, 4174–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, M.; Tayeb, H.T.; Al Johani, S.M.; Alyamani, E.J.; Aldughaishem, F.; Alabdulkarim, I.; Balkhy, H.H. Genetic Diversity of OXA-51-like Genes among Multidrug-Resistant Acinetobacter Baumannii in Riyadh, Saudi Arabia. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2014, 33, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Hamouda, A.; Towner, K.J.; Amyes, S.G.B. OXA-51-like Beta-Lactamases and Their Association with Particular Epidemic Lineages of Acinetobacter Baumannii. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2008, 14, 268–275. [Google Scholar] [CrossRef]
- Rafei, R.; Pailhoriès, H.; Hamze, M.; Eveillard, M.; Mallat, H.; Dabboussi, F.; Joly-Guillou, M.-L.; Kempf, M. Molecular Epidemiology of Acinetobacter Baumannii in Different Hospitals in Tripoli, Lebanon Using Bla(OXA-51-like) Sequence Based Typing. BMC Microbiol. 2015, 15, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.A.; Brown, S.; Hamouda, A.; Findlay, J.; Amyes, S.G.B. Eleven Novel OXA-51-like Enzymes from Clinical Isolates of Acinetobacter Baumannii. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2007, 1137–1138. [Google Scholar] [CrossRef]
- Poirel, L.; Marqué, S.; Héritier, C.; Segonds, C.; Chabanon, G.; Nordmann, P. OXA-58, a Novel Class D {beta}-Lactamase Involved in Resistance to Carbapenems in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2005, 49, 202–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Mansour, W.; Bouallegue, O.; Nordmann, P. Carbapenem-Resistant Acinetobacter Baumannii Isolates from Tunisia Producing the OXA-58-like Carbapenem-Hydrolyzing Oxacillinase OXA-97. Antimicrob. Agents Chemother. 2008, 52, 1613–1617. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Schneiders, T.; Hamprecht, A.; Seifert, H. In Vivo Selection of a Missense Mutation in AdeR and Conversion of the Novel BlaOXA-164 Gene into BlaOXA-58 in Carbapenem-Resistant Acinetobacter Baumannii Isolates from a Hospitalized Patient. Antimicrob. Agents Chemother. 2010, 54, 5021–5027. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Poirel, L.; Lehmann, M.; Nordmann, P.; Seifert, H. OXA-143, a Novel Carbapenem-Hydrolyzing Class D Beta-Lactamase in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2009, 53, 5035–5038. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-K.; Lee, Y.; Lee, H.; Woo, G.-J.; Song, W.; Kim, M.-N.; Lee, W.-G.; Jeong, S.H.; Lee, K.; Chong, Y. Prevalence and Diversity of Carbapenemases among Imipenem-Nonsusceptible Acinetobacter Isolates in Korea: Emergence of a Novel OXA-182. Diagn. Microbiol. Infect. Dis. 2010, 68, 432–438. [Google Scholar] [CrossRef]
- Gionco, B.; Pelayo, J.S.; Venancio, E.J.; Cayô, R.; Gales, A.C.; Carrara-Marroni, F.E. Detection of OXA-231, a New Variant of BlaOXA-143, in Acinetobacter Baumannii from Brazil: A Case Report. J. Antimicrob. Chemother. 2012, 2531–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostachio, A.K.; Levin, A.S.; Rizek, C.; Rossi, F.; Zerbini, J.; Costa, S.F. High Prevalence of OXA-143 and Alteration of Outer Membrane Proteins in Carbapenem-Resistant Acinetobacter spp. Isolates in Brazil. Int. J. Antimicrob. Agents 2012, 39, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.; Hernández, J.M.; Espasa, M.; Fleites, A.; Sáez, D.; Bautista, V.; Pérez-Vázquez, M.; Fernández-García, M.D.; Delgado-Iribarren, A.; Sánchez-Romero, I.; et al. Emergence of OXA-48-Producing Klebsiella Pneumoniae and the Novel Carbapenemases OXA-244 and OXA-245 in Spain. J. Antimicrob. Chemother. 2013, 68, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potron, A.; Poirel, L.; Nordmann, P. Origin of OXA-181, an Emerging Carbapenem-Hydrolyzing Oxacillinase, as a Chromosomal Gene in Shewanella Xiamenensis. Antimicrob. Agents Chemother. 2011, 55, 4405–4407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Z. Discovery of Bla(OXA-199), a Chromosome-Based Bla(OXA-48)-like Variant, in Shewanella Xiamenensis. PLoS ONE 2012, 7, e48280. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Rondinaud, E.; Poirel, L.; Belmonte, O.; Boyer, S.; Camiade, S.; Nordmann, P. Genetic and Biochemical Characterisation of OXA-232, a Carbapenem-Hydrolysing Class D β-Lactamase from Enterobacteriaceae. Int. J. Antimicrob. Agents 2013, 41, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Castanheira, M.; Carrër, A.; Rodriguez, C.P.; Jones, R.N.; Smayevsky, J.; Nordmann, P. OXA-163, an OXA-48-Related Class D β-Lactamase with Extended Activity toward Expanded-Spectrum Cephalosporins. Antimicrob. Agents Chemother. 2011, 55, 2546–2551. [Google Scholar] [CrossRef] [Green Version]
- Boyd, D.A.; Mataseje, L.F.; Pelude, L.; Mitchell, R.; Bryce, E.; Roscoe, D.; Embree, J.; Katz, K.; Kibsey, P.; Lavallee, C.; et al. Results from the Canadian Nosocomial Infection Surveillance Program for Detection of Carbapenemase-Producing Acinetobacter spp. in Canadian Hospitals, 2010–2016. J. Antimicrob. Chemother. 2019, 74, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yu, H.; Guo, Q.; Xu, X.; Ye, X.; Wu, S.; Guo, Y.; Wang, M. Distribution of 16S RRNA Methylases among Different Species of Gram-Negative Bacilli with High-Level Resistance to Aminoglycosides. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2010, 29, 1349–1353. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside Modifying Enzymes. Drug Resist. Update Rev. Comment. Antimicrob. Anticancer Chemother. 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Miyoshi-Akiyama, T.; Kato, Y.; Ohmagari, N.; Takeshita, N.; Hung, N.V.; Phuong, D.M.; Thu, T.A.; Binh, N.G.; Anh, N.Q.; et al. Emergence of 16S RRNA Methylase-Producing Acinetobacter Baumannii and Pseudomonas Aeruginosa Isolates in Hospitals in Vietnam. BMC Infect. Dis. 2013, 13, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.-T.; Zhou, Y.; Yang, L.; Xu, Y. Multidrug-Resistant Genes of Aminoglycoside-Modifying Enzymes and 16S RRNA Methylases in Acinetobacter Baumannii Strains. Genet. Mol. Res. 2014, 13, 3842–3849. [Google Scholar] [CrossRef] [PubMed]
- Heidary, M.; Salimi Chirani, A.; Khoshnood, S.; Eslami, G.; Atyabi, S.M.; Nazem, H.; Fazilati, M.; Hashemi, A.; Soleimani, S. Molecular Detection of Aminoglycoside-Modifying Enzyme Genes in Acinetobacter Baumannii Clinical Isolates. Acta Microbiol. Immunol. Hung. 2017, 64, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hujer, K.M.; Hujer, A.M.; Hulten, E.A.; Bajaksouzian, S.; Adams, J.M.; Donskey, C.J.; Ecker, D.J.; Massire, C.; Eshoo, M.W.; Sampath, R.; et al. Analysis of Antibiotic Resistance Genes in Multidrug-Resistant Acinetobacter Sp. Isolates from Military and Civilian Patients Treated at the Walter Reed Army Medical Center. Antimicrob. Agents Chemother. 2006, 50, 4114–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, S.J.; Post, V.; Hall, R.M. Aminoglycoside Resistance in Multiply Antibiotic-Resistant Acinetobacter Baumannii Belonging to Global Clone 2 from Australian Hospitals. J. Antimicrob. Chemother. 2011, 66, 1504–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghazadeh, M.; Rezaee, M.A.; Nahaei, M.R.; Mahdian, R.; Pajand, O.; Saffari, F.; Hassan, M.; Hojabri, Z. Dissemination of Aminoglycoside-Modifying Enzymes and 16S RRNA Methylases among Acinetobacter Baumannii and Pseudomonas Aeruginosa Isolates. Microb. Drug Resist. 2013, 19, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Hasani, A.; Sheikhalizadeh, V.; Ahangarzadeh Rezaee, M.; Rahmati-Yamchi, M.; Hasani, A.; Ghotaslou, R.; Goli, H.R. Frequency of Aminoglycoside-Modifying Enzymes and ArmA Among Different Sequence Groups of Acinetobacter Baumannii in Iran. Microb. Drug Resist. 2016, 22, 347–353. [Google Scholar] [CrossRef]
- Sheikhalizadeh, V.; Hasani, A.; Ahangarzadeh Rezaee, M.; Rahmati-Yamchi, M.; Hasani, A.; Ghotaslou, R.; Goli, H.R. Comprehensive Study to Investigate the Role of Various Aminoglycoside Resistance Mechanisms in Clinical Isolates of Acinetobacter Baumannii. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2017, 23, 74–79. [Google Scholar] [CrossRef]
- Salimizand, H.; Zomorodi, A.R.; Mansury, D.; Khakshoor, M.; Azizi, O.; Khodaparast, S.; Baseri, Z.; Karami, P.; Zamanlou, S.; Farsiani, H.; et al. Diversity of Aminoglycoside Modifying Enzymes and 16S RRNA Methylases in Acinetobacter Baumannii and Acinetobacter Nosocomialis Species in Iran; Wide Distribution of AadA1 and ArmA. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 66, 195–199. [Google Scholar] [CrossRef]
- Shooshtari, F.S.; Navidifar, T.; Amin, M.; Goodarzi, H. Coexistence of Genes Encoding Aminoglycoside Modifying Enzymes among Clinical Acinetobacter Baumannii Isolates in Ahvaz, Southwest Iran. Acta Microbiol. Immunol. Hung. 2019, 1–9. [Google Scholar] [CrossRef]
- Akers, K.S.; Chaney, C.; Barsoumian, A.; Beckius, M.; Zera, W.; Yu, X.; Guymon, C.; Keen, E.F., 3rd; Robinson, B.J.; Mende, K.; et al. Aminoglycoside Resistance and Susceptibility Testing Errors in Acinetobacter Baumannii-Calcoaceticus Complex. J. Clin. Microbiol. 2010, 48, 1132–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y.; Arakawa, Y. 16S Ribosomal RNA Methylation: Emerging Resistance Mechanism against Aminoglycosides. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of Resistance to Aminoglycoside Antibiotics: Overview and Perspectives. Medchemcomm 2016, 7, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-Nodulation-Cell Division-Type Efflux Pump Involved in Aminoglycoside Resistance in Acinetobacter Baumannii Strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [Green Version]
- Savic, M.; Lovric, J.; Tomic, T.I.; Vasiljevic, B.; Conn, G.L. Determination of the Target Nucleosides for Members of Two Families of 16S RRNA Methyltransferases That Confer Resistance to Partially Overlapping Groups of Aminoglycoside Antibiotics. Nucleic Acids Res. 2009, 37, 5420–5431. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Cao, J.; Zhou, C.; Liu, H.; Zhang, X.; Zhou, T. Biofilm Formation Restrained by Subinhibitory Concentrations of Tigecyclin in Acinetobacter Baumannii Is Associated with Downregulation of Efflux Pumps. Chemotherapy 2017, 62, 128–133. [Google Scholar] [CrossRef]
- Butaye, P.; Cloeckaert, A.; Schwarz, S. Mobile Genes Coding for Efflux-Mediated Antimicrobial Resistance in Gram-Positive and Gram-Negative Bacteria. Int. J. Antimicrob. Agents 2003, 22, 205–210. [Google Scholar] [CrossRef]
- Poole, K. Efflux-Mediated Antimicrobial Resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Coyne, S.; Courvalin, P.; Périchon, B. Efflux-Mediated Antibiotic Resistance in Acinetobacter spp. Antimicrob. Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Yoon, E.-J.; Chabane, Y.N.; Goussard, S.; Snesrud, E.; Courvalin, P.; Dé, E.; Grillot-Courvalin, C. Contribution of Resistance-Nodulation-Cell Division Efflux Systems to Antibiotic Resistance and Biofilm Formation in Acinetobacter Baumannii. MBio 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-Type Efflux Pump AdeABC in Acinetobacter Baumannii Is Regulated by the AdeRS Two-Component System. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [Green Version]
- Damier-Piolle, L.; Magnet, S.; Brémont, S.; Lambert, T.; Courvalin, P. AdeIJK, a Resistance-Nodulation-Cell Division Pump Effluxing Multiple Antibiotics in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2008, 52, 557–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richmond, G.E.; Evans, L.P.; Anderson, M.J.; Wand, M.E.; Bonney, L.C.; Ivens, A.; Chua, K.L.; Webber, M.A.; Sutton, J.M.; Peterson, M.L.; et al. The Acinetobacter Baumannii Two-Component System AdeRS Regulates Genes Required for Multidrug Efflux, Biofilm Formation, and Virulence in a Strain-Specific Manner. MBio 2016, 7, e00430-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, N.; Bouchier, C.; Courvalin, P.; Périchon, B. Expression of the Resistance-Nodulation-Cell Division Pump AdeIJK in Acinetobacter Baumannii Is Regulated by AdeN, a TetR-Type Regulator. Antimicrob. Agents Chemother. 2012, 56, 2504–2510. [Google Scholar] [CrossRef] [Green Version]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Périchon, B. Overexpression of Resistance-Nodulation-Cell Division Pump AdeFGH Confers Multidrug Resistance in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef] [Green Version]
- Su, X.-Z.; Chen, J.; Mizushima, T.; Kuroda, T.; Tsuchiya, T. AbeM, an H+-Coupled Acinetobacter Baumannii Multidrug Efflux Pump Belonging to the MATE Family of Transporters. Antimicrob. Agents Chemother. 2005, 49, 4362–4364. [Google Scholar] [CrossRef] [Green Version]
- Hou, P.F.; Chen, X.Y.; Yan, G.F.; Wang, Y.P.; Ying, C.M. Study of the Correlation of Imipenem Resistance with Efflux Pumps AdeABC, AdeIJK, AdeDE and AbeM in Clinical Isolates of Acinetobacter Baumannii. Chemotherapy 2012, 58, 152–158. [Google Scholar] [CrossRef]
- Rumbo, C.; Gato, E.; López, M.; Ruiz de Alegría, C.; Fernández-Cuenca, F.; Martínez-Martínez, L.; Vila, J.; Pachón, J.; Cisneros, J.M.; Rodríguez-Baño, J.; et al. Contribution of Efflux Pumps, Porins, and β-Lactamases to Multidrug Resistance in Clinical Isolates of Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2013, 57, 5247–5257. [Google Scholar] [CrossRef] [Green Version]
- Roca, I.; Marti, S.; Espinal, P.; Martínez, P.; Gibert, I.; Vila, J. CraA, a Major Facilitator Superfamily Efflux Pump Associated with Chloramphenicol Resistance in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2009, 53, 4013–4014. [Google Scholar] [CrossRef] [Green Version]
- Ribera, A.; Roca, I.; Ruiz, J.; Gibert, I.; Vila, J. Partial Characterization of a Transposon Containing the Tet(A) Determinant in a Clinical Isolate of Acinetobacter Baumannii. J. Antimicrob. Chemother. 2003, 52, 477–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foong, W.E.; Wilhelm, J.; Tam, H.-K.; Pos, K.M. Tigecycline Efflux in Acinetobacter Baumannii Is Mediated by TetA in Synergy with RND-Type Efflux Transporters. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, G.; Srinivasan, V.B.; Gebreyes, W.A. Molecular and Functional Characterization of a Novel Efflux Pump, AmvA, Mediating Antimicrobial and Disinfectant Resistance in Acinetobacter Baumannii. J. Antimicrob. Chemother. 2010, 65, 1919–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Sharma, R.; Bhattacharyya, T.; Bhando, T.; Pathania, R. Fosfomycin Resistance in Acinetobacter Baumannii Is Mediated by Efflux through a Major Facilitator Superfamily (MFS) Transporter-AbaF. J. Antimicrob. Chemother. 2017, 72, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Varela, M.; Corral, J.; Aranda, J.; Barbé, J. Functional Characterization of AbaQ, a Novel Efflux Pump Mediating Quinolone Resistance in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, V.B.; Rajamohan, G.; Gebreyes, W.A. Role of AbeS, a Novel Efflux Pump of the SMR Family of Transporters, in Resistance to Antimicrobial Agents in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2009, 53, 5312–5316. [Google Scholar] [CrossRef] [Green Version]
- Lytvynenko, I.; Brill, S.; Oswald, C.; Pos, K.M. Molecular Basis of Polyspecificity of the Small Multidrug Resistance Efflux Pump AbeS from Acinetobacter Baumannii. J. Mol. Biol. 2016, 428, 644–657. [Google Scholar] [CrossRef]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter Baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou, G.; Cerveró, G.; Domínguez, M.A.; Quereda, C.; Martínez-Beltrán, J. Characterization of a Nosocomial Outbreak Caused by a Multiresistant Acinetobacter Baumannii Strain with a Carbapenem-Hydrolyzing Enzyme: High-Level Carbapenem Resistance in A. Baumannii Is Not Due Solely to the Presence of Beta-Lactamases. J. Clin. Microbiol. 2000, 38, 3299–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, M.; Pagès, J.-M.; Lafitte, D.; Siroy, A.; Bollet, C. Identification of an OprD Homologue in Acinetobacter Baumannii. J. Proteome Res. 2005, 4, 2386–2390. [Google Scholar] [CrossRef] [PubMed]
- Quale, J.; Bratu, S.; Landman, D.; Heddurshetti, R. Molecular Epidemiology and Mechanisms of Carbapenem Resistance in Acinetobacter Baumannii Endemic in New York City. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2003, 37, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Mar Tomás, M.; Beceiro, A.; Pérez, A.; Velasco, D.; Moure, R.; Villanueva, R.; Martínez-Beltrán, J.; Bou, G. Cloning and Functional Analysis of the Gene Encoding the 33- to 36-Kilodalton Outer Membrane Protein Associated with Carbapenem Resistance in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2005, 49, 5172–5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.I.; Jacobs, A.C.; Sayood, K.; Dunman, P.M.; Skaar, E.P. Acinetobacter Baumannii Increases Tolerance to Antibiotics in Response to Monovalent Cations. Antimicrob. Agents Chemother. 2010, 54, 1029–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of Resistance to Carbapenems in Multidrug-Resistant Clinical Strains of Acinetobacter Baumannii: Natural Insertional Inactivation of a Gene Encoding a Member of a Novel Family of Beta-Barrel Outer Membrane Proteins. Antimicrob. Agents Chemother. 2005, 49, 1432–1440. [Google Scholar] [CrossRef] [Green Version]
- Mussi, M.A.; Relling, V.M.; Limansky, A.S.; Viale, A.M. CarO, an Acinetobacter Baumannii Outer Membrane Protein Involved in Carbapenem Resistance, Is Essential for L-Ornithine Uptake. FEBS Lett. 2007, 581, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Siroy, A.; Molle, V.; Lemaître-Guillier, C.; Vallenet, D.; Pestel-Caron, M.; Cozzone, A.J.; Jouenne, T.; Dé, E. Channel Formation by CarO, the Carbapenem Resistance-Associated Outer Membrane Protein of Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2005, 49, 4876–4883. [Google Scholar] [CrossRef] [Green Version]
- Catel-Ferreira, M.; Coadou, G.; Molle, V.; Mugnier, P.; Nordmann, P.; Siroy, A.; Jouenne, T.; Dé, E. Structure-Function Relationships of CarO, the Carbapenem Resistance-Associated Outer Membrane Protein of Acinetobacter Baumannii. J. Antimicrob. Chemother. 2011, 66, 2053–2056. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.S.; Kwon, S.-O.; Moon, D.C.; Gurung, M.; Lee, J.H.; Kim, S.I.; Lee, J.C. Acinetobacter Baumannii Secretes Cytotoxic Outer Membrane Protein A via Outer Membrane Vesicles. PLoS ONE 2011, 6, e17027. [Google Scholar] [CrossRef]
- Smani, Y.; Fàbrega, A.; Roca, I.; Sánchez-Encinales, V.; Vila, J.; Pachón, J. Role of OmpA in the Multidrug Resistance Phenotype of Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2014, 58, 1806–1808. [Google Scholar] [CrossRef] [Green Version]
- Espinal, P.; Pantel, A.; Rolo, D.; Marti, S.; López-Rojas, R.; Smani, Y.; Pachón, J.; Vila, J.; Lavigne, J.-P. Relationship Between Different Resistance Mechanisms and Virulence in Acinetobacter Baumannii. Microb. Drug Resist. 2019, 25, 752–760. [Google Scholar] [CrossRef]
- Sánchez-Encinales, V.; Álvarez-Marín, R.; Pachón-Ibáñez, M.E.; Fernández-Cuenca, F.; Pascual, A.; Garnacho-Montero, J.; Martínez-Martínez, L.; Vila, J.; Tomás, M.M.; Cisneros, J.M.; et al. Overproduction of Outer Membrane Protein A by Acinetobacter Baumannii as a Risk Factor for Nosocomial Pneumonia, Bacteremia, and Mortality Rate Increase. J. Infect. Dis. 2017, 215, 966–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Unno, Y.; Kawakami, S.; Ubagai, T.; Ono, Y. Virulence Characteristics of Acinetobacter Baumannii Clinical Isolates Vary with the Expression Levels of Omps. J. Med. Microbiol. 2017, 66, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Eghbali, Z.; Mojtahedi, A.; Moien Ansar, M.; Fakhrieh Asl, S.; Aminian, K. Detection of 23SrRNA Mutations Strongly Related to Clarithromycin Resistance in Helicobacter Pylori Strains Isolated From Patients in the North of Iran. Jundishapur J. Microbiol. 2016, 9, e29694. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Kelesidis, T.; Tsiodras, S.; Hindler, J.; Humphries, R.M. The Emerging Problem of Linezolid-Resistant Staphylococcus. J. Antimicrob. Chemother. 2013, 68, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural Mechanism for Rifampicin Inhibition of Bacterial Rna Polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Floss, H.G.; Yu, T.-W. Rifamycin-Mode of Action, Resistance, and Biosynthesis. Chem. Rev. 2005, 105, 621–632. [Google Scholar] [CrossRef]
- Park, S.; Lee, K.M.; Yoo, Y.S.; Yoo, J.S.; Yoo, J.I.; Kim, H.S.; Lee, Y.S.; Chung, G.T. Alterations of GyrA, GyrB, and ParC and Activity of Efflux Pump in Fluoroquinolone-Resistant Acinetobacter Baumannii. Osong Public Health Res. Perspect. 2011, 2, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Ardebili, A.; Lari, A.R.; Beheshti, M.; Lari, E.R. Association between Mutations in GyrA and ParC Genes of Acinetobacter Baumannii Clinical Isolates and Ciprofloxacin Resistance. Iran. J. Basic Med. Sci. 2015, 18, 623–626. [Google Scholar]
- Hamed, S.M.; Elkhatib, W.F.; El-Mahallawy, H.A.; Helmy, M.M.; Ashour, M.S.; Aboshanab, K.M.A. Multiple Mechanisms Contributing to Ciprofloxacin Resistance among Gram Negative Bacteria Causing Infections to Cancer Patients. Sci. Rep. 2018, 8, 12268. [Google Scholar] [CrossRef]
- Hooper, D.C. Fluoroquinolone Resistance among Gram-Positive Cocci. Lancet Infect. Dis. 2002, 2, 530–538. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Hofmann, B.; Hansen, B.; Scheuring, S.; Lückefahr, M.; Klootwijk, M.; Verhoef, J.; Fluit, A.; Heinz, H.P.; Köhrer, K.; et al. Relationship between Ciprofloxacin, Ofloxacin, Levofloxacin, Sparfloxacin and Moxifloxacin (BAY 12-8039) MICs and Mutations in GrlA, GrlB, GyrA and GyrB in 116 Unrelated Clinical Isolates of Staphylococcus Aureus. J. Antimicrob. Chemother. 1998, 41, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Albornoz, E.; Tijet, N.; De Belder, D.; Gomez, S.; Martino, F.; Corso, A.; Melano, R.G.; Petroni, A. QnrE1, a Member of a New Family of Plasmid-Located Quinolone Resistance Genes, Originated from the Chromosome of Enterobacter Species. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Jacoby, G.A.; Macielag, M.; Abbanat, D.; Park, C.H.; Bush, K.; Hooper, D.C. Fluoroquinolone-Modifying Enzyme: A New Adaptation of a Common Aminoglycoside Acetyltransferase. Nat. Med. 2006, 12, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the Treatment of Multidrug-Resistant Pathogens: A Critical Review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, M.D.; Stern, R.C.; O’Riordan, M.A.; Blumer, J.L. The Pharmacokinetics of Colistin in Patients with Cystic Fibrosis. J. Clin. Pharmacol. 2001, 41, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Bialvaei, A.Z.; Samadi Kafil, H. Colistin, Mechanisms and Prevalence of Resistance. Curr. Med. Res. Opin. 2015, 31, 707–721. [Google Scholar] [CrossRef]
- Hejnar, P.; Kolár, M.; Hájek, V. Characteristics of Acinetobacter Strains (Phenotype Classification, Antibiotic Susceptibility and Production of Beta-Lactamases) Isolated from Haemocultures from Patients at the Teaching Hospital in Olomouc. Acta Univ. Palacki. Olomuc. Fac. Med. 1999, 142, 73–77. [Google Scholar]
- Adams, M.D.; Nickel, G.C.; Bajaksouzian, S.; Lavender, H.; Murthy, A.R.; Jacobs, M.R.; Bonomo, R.A. Resistance to Colistin in Acinetobacter Baumannii Associated with Mutations in the PmrAB Two-Component System. Antimicrob. Agents Chemother. 2009, 53, 3628–3634. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-Y.; Gregg, K.A.; Napier, B.A.; Ernst, R.K.; Weiss, D.S. A PmrB-Regulated Deacetylase Required for Lipid A Modification and Polymyxin Resistance in Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2015, 59, 7911–7914. [Google Scholar] [CrossRef] [Green Version]
- Koomanachai, P.; Kim, A.; Nicolau, D.P. Pharmacodynamic Evaluation of Tigecycline against Acinetobacter Baumannii in a Murine Pneumonia Model. J. Antimicrob. Chemother. 2009, 63, 982–987. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, M.; Mendes, R.E.; Jones, R.N. Update on Acinetobacter Species: Mechanisms of Antimicrobial Resistance and Contemporary in Vitro Activity of Minocycline and Other Treatment Options. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2014, 59 (Suppl. 6), S367–S373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, K.R.; Kim, M.J.; Choi, J.Y.; Kim, H.S.; Kang, C.-I.; Cho, Y.K.; Park, D.W.; Lee, H.J.; Lee, M.S.; Ko, K.S. In Vitro Time-Kill Studies of Antimicrobial Agents against Blood Isolates of Imipenem-Resistant Acinetobacter Baumannii, Including Colistin- or Tigecycline-Resistant Isolates. J. Med. Microbiol. 2012, 61, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, H.; Akduman, D.; Piskin, N.; Comert, F.; Horuz, E.; Terzi, A.; Kokturk, F.; Ornek, T.; Celebi, G. Colistin vs. the Combination of Colistin and Rifampicin for the Treatment of Carbapenem-Resistant Acinetobacter Baumannii Ventilator-Associated Pneumonia. Epidemiol. Infect. 2013, 141, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Menegucci, T.C.; Fedrigo, N.H.; Lodi, F.G.; Albiero, J.; Nishiyama, S.A.B.; Mazucheli, J.; Carrara-Marroni, F.E.; Voelkner, N.M.F.; Gong, H.; Sy, S.K.B.; et al. Pharmacodynamic Effects of Sulbactam/Meropenem/Polymyxin-B Combination Against Extremely Drug Resistant Acinetobacter Baumannii Using Checkerboard Information. Microb. Drug Resist. 2019, 25, 1266–1274. [Google Scholar] [CrossRef]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E.W. Host Defense Peptides: Front-Line Immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Falanga, A.; Lombardi, L.; Franci, G.; Vitiello, M.; Iovene, M.R.; Morelli, G.; Galdiero, M.; Galdiero, S. Marine Antimicrobial Peptides: Nature Provides Templates for the Design of Novel Compounds against Pathogenic Bacteria. Int. J. Mol. Sci. 2016, 17, 785. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Sun, J.; Zhou, M.; Zhou, J.; Lao, X.; Zheng, H.; Xu, H. DRAMP: A Comprehensive Data Repository of Antimicrobial Peptides. Sci. Rep. 2016, 6, 24482. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Govender, T.; Dawood, A.; Esterhuyse, A.J.; Katerere, D.R. Antimicrobial Properties of the Skin Secretions of Frogs. S. Afr. J. Sci. 2012, 108, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of Antimicrobial Peptides. A Review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Lan, X.-Q.; Du, Y.; Chen, P.-Y.; Zhao, J.; Zhao, F.; Lee, W.-H.; Zhang, Y. King Cobra Peptide OH-CATH30 as a Potential Candidate Drug through Clinic Drug-Resistant Isolates. Zool. Res. 2018, 39, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekan, Z.; Headey, S.J.; Scanlon, M.; Baldo, B.A.; Lee, T.-H.; Aguilar, M.-I.; Deuis, J.R.; Vetter, I.; Elliott, A.G.; Amado, M.; et al. Δ-Myrtoxin-Mp1a Is a Helical Heterodimer from the Venom of the Jack Jumper Ant That Has Antimicrobial, Membrane-Disrupting, and Nociceptive Activities. Angew. Chem. Int. Ed. Engl. 2017, 56, 8495–8499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The Antimicrobial Peptide ZY4 Combats Multidrug-Resistant Pseudomonas Aeruginosa and Acinetobacter Baumannii Infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajbakhsh, M.; Akhavan, M.M.; Fallah, F.; Karimi, A. A Recombinant Snake Cathelicidin Derivative Peptide: Antibiofilm Properties and Expression in Escherichia Coli. Biomolecules 2018, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, A.K.; Alamri, M.A.; Almasoudi, A.S.; Abbasmanthiri, R.; Mahfoud, M. Evaluation of the in Vitro Antimicrobial Activity of Selected Saudi Scorpion Venoms Tested against Multidrug-Resistant Micro-Organisms. J. Glob. Antimicrob. Resist. 2017, 10, 14–18. [Google Scholar] [CrossRef]
- Lin, C.-H.; Lee, M.-C.; Tzen, J.T.C.; Lee, H.-M.; Chang, S.-M.; Tu, W.-C.; Lin, C.-F. Efficacy of Mastoparan-AF Alone and in Combination with Clinically Used Antibiotics on Nosocomial Multidrug-Resistant Acinetobacter Baumannii. Saudi J. Biol. Sci. 2017, 24, 1023–1029. [Google Scholar] [CrossRef]
- Shamova, O.V.; Orlov, D.S.; Zharkova, M.S.; Balandin, S.V.; Yamschikova, E.V.; Knappe, D.; Hoffmann, R.; Kokryakov, V.N.; Ovchinnikova, T.V. Minibactenecins ChBac7.Nα and ChBac7. Nβ—Antimicrobial Peptides from Leukocytes of the Goat Capra Hircus. Acta Nat. 2016, 8, 136–146. [Google Scholar] [CrossRef]
- Hirsch, R.; Wiesner, J.; Marker, A.; Pfeifer, Y.; Bauer, A.; Hammann, P.E.; Vilcinskas, A. Profiling Antimicrobial Peptides from the Medical Maggot Lucilia Sericata as Potential Antibiotics for MDR Gram-Negative Bacteria. J. Antimicrob. Chemother. 2019, 74, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Domhan, C.; Uhl, P.; Kleist, C.; Zimmermann, S.; Umstätter, F.; Leotta, K.; Mier, W.; Wink, M. Replacement of L-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution. Molecules 2019, 24, 2987. [Google Scholar] [CrossRef] [Green Version]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial Activity of Selected Antimicrobial Peptides Against Planktonic Culture and Biofilm of Acinetobacter Baumannii. Probiotics Antimicrob. Proteins 2019, 11, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Long, H.; Liu, W.; Wu, Z.; Wang, T.; Zeng, Z.; Guo, G.; Wu, J. Antibacterial Mechanism of Peptide Cec4 against Acinetobacter Baumannii. Infect. Drug Resist. 2019, 12, 2417–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.M.; Ko, S.; Cheong, M.-J.; Bang, J.K.; Seo, C.H.; Luchian, T.; Park, Y. Myxinidin2 and Myxinidin3 Suppress Inflammatory Responses through STAT3 and MAPKs to Promote Wound Healing. Oncotarget 2017, 8, 87582–87597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordya, N.; Yakovlev, A.; Kruglikova, A.; Tulin, D.; Potolitsina, E.; Suborova, T.; Bordo, D.; Rosano, C.; Chernysh, S. Natural Antimicrobial Peptide Complexes in the Fighting of Antibiotic Resistant Biofilms: Calliphora Vicina Medicinal Maggots. PLoS ONE 2017, 12, e0173559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourtada, R.; Herce, H.D.; Yin, D.J.; Moroco, J.A.; Wales, T.E.; Engen, J.R.; Walensky, L.D. Design of Stapled Antimicrobial Peptides That Are Stable, Nontoxic and Kill Antibiotic-Resistant Bacteria in Mice. Nat. Biotechnol. 2019, 37, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Lapuebla, A.; Landman, D.; Quale, J. In Vitro and In Vivo Activity of a Novel Antisense Peptide Nucleic Acid Compound Against Multidrug-Resistant Acinetobacter Baumannii. Microb. Drug Resist. 2019, 25, 961–965. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T. Structural Features of Helical Antimicrobial Peptides: Their Potential to Modulate Activity on Model Membranes and Biological Cells. Biochim. Biophys. Acta 1999, 1462, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Starr, C.G.; Wimley, W.C. Antimicrobial Peptides Are Degraded by the Cytosolic Proteases of Human Erythrocytes. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2319–2326. [Google Scholar] [CrossRef]
- McPhee, J.B.; Hancock, R.E.W. Function and Therapeutic Potential of Host Defence Peptides. J. Pept. Sci. 2005, 11, 677–687. [Google Scholar] [CrossRef]
- Cantisani, M.; Finamore, E.; Mignogna, E.; Falanga, A.; Nicoletti, G.F.; Pedone, C.; Morelli, G.; Leone, M.; Galdiero, M.; Galdiero, S. Structural Insights into and Activity Analysis of the Antimicrobial Peptide Myxinidin. Antimicrob. Agents Chemother. 2014, 58, 5280–5290. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Jang, J.H.; Kim, S.C.; Cho, J.H. De Novo Generation of Short Antimicrobial Peptides with Enhanced Stability and Cell Specificity. J. Antimicrob. Chemother. 2014, 69, 121–132. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and Consequences of Bacterial Resistance to Antimicrobial Peptides. Drug Resist. Update Rev. Comment. Antimicrob. Anticancer Chemother. 2016, 26, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Xu, M.; Wang, T.; You, C.; Wang, X.; Ren, H.; Zhou, H.; Khan, A.; Han, C.; Li, P. Catechol Cross-Linked Antimicrobial Peptide Hydrogels Prevent Multidrug-Resistant Acinetobacter Baumannii Infection in Burn Wounds. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostorhazi, E.; Hoffmann, R.; Herth, N.; Wade, J.D.; Kraus, C.N.; Otvos, L.J. Advantage of a Narrow Spectrum Host Defense (Antimicrobial) Peptide Over a Broad Spectrum Analog in Preclinical Drug Development. Front. Chem. 2018, 6, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mant, C.T.; Jiang, Z.; Gera, L.; Davis, T.; Nelson, K.L.; Bevers, S.; Hodges, R.S. De Novo Designed Amphipathic α-Helical Antimicrobial Peptides Incorporating Dab and Dap Residues on the Polar Face To Treat the Gram-Negative Pathogen, Acinetobacter Baumannii. J. Med. Chem. 2019, 62, 3354–3366. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Robinson, A.; Davidov, Y.; Shulman, A.; Varvak, A.; Reyes, F.; Rahav, G.; Nissan, I. Delftibactin-A, a Non-Ribosomal Peptide With Broad Antimicrobial Activity. Front. Microbiol. 2019, 10, 2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri, B.; Mohammadi, M.; Momenzadeh, N.; Farshadzadeh, Z.; Roozbehani, M.; Dehghani, P.; Hajian, S.; Darvishi, S.; Shamseddin, J. Substitution of Lysine for Isoleucine at the Center of the Nonpolar Face of the Antimicrobial Peptide, Piscidin-1, Leads to an Increase in the Rapidity of Bactericidal Activity and a Reduction in Toxicity. Infect. Drug Resist. 2019, 12, 1629–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, D.; Roy, N.; Kulkarni, O.; Nanajkar, N.; Datey, A.; Ravichandran, S.; Thakur, C.; Sandeep, T.; Aprameya, I.V.; Sarma, S.P.; et al. Ω76: A Designed Antimicrobial Peptide to Combat Carbapenem- and Tigecycline-Resistant Acinetobacter Baumannii. Sci. Adv. 2019, 5, eaax1946. [Google Scholar] [CrossRef] [Green Version]
- Hacioglu, M.; Oyardi, O.; Bozkurt-Guzel, C.; Savage, P.B. Antibiofilm Activities of Ceragenins and Antimicrobial Peptides against Fungal-Bacterial Mono and Multispecies Biofilms. J. Antibiot. 2020. [Google Scholar] [CrossRef]
- Jenei, S.; Tiricz, H.; Szolomájer, J.; Tímár, E.; Klement, É.; Al Bouni, M.A.; Lima, R.M.; Kata, D.; Harmati, M.; Buzás, K.; et al. Potent Chimeric Antimicrobial Derivatives of the Medicago Truncatula NCR247 Symbiotic Peptide. Front. Microbiol. 2020, 11, 270. [Google Scholar] [CrossRef]
- Heulot, M.; Jacquier, N.; Aeby, S.; Le Roy, D.; Roger, T.; Trofimenko, E.; Barras, D.; Greub, G.; Widmann, C. The Anticancer Peptide TAT-RasGAP(317-326) Exerts Broad Antimicrobial Activity. Front. Microbiol. 2017, 8, 994. [Google Scholar] [CrossRef]
- Swedan, S.; Shubair, Z.; Almaaytah, A. Synergism of Cationic Antimicrobial Peptide WLBU2 with Antibacterial Agents against Biofilms of Multi-Drug Resistant Acinetobacter Baumannii and Klebsiella Pneumoniae. Infect. Drug Resist. 2019, 12, 2019–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-L.; Su, P.-Y.; Kuo, S.-C.; Lauderdale, T.-L.Y.; Shih, C. Adding a C-Terminal Cysteine (CTC) Can Enhance the Bactericidal Activity of Three Different Antimicrobial Peptides. Front. Microbiol. 2018, 9, 1440. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.M.; Zorgani, A.; Jalowicki, G.; Kerr, A.; Khaldi, N.; Martins, M. Unlocking NuriPep 1653 From Common Pea Protein: A Potent Antimicrobial Peptide to Tackle a Pan-Drug Resistant Acinetobacter Baumannii. Front. Microbiol. 2019, 10, 2086. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Gupta, K.; van Hoek, M.L. Characterization of Cimex Lectularius (Bedbug) Defensin Peptide and Its Antimicrobial Activity against Human Skin Microflora. Biochem. Biophys. Res. Commun. 2016, 470, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, S.H.; Murphy, R.A.; Juul-Madsen, K.; Fredborg, M.; Hvam, M.L.; Axelgaard, E.; Skovdal, S.M.; Meyer, R.L.; Sørensen, U.B.S.; Möller, A.; et al. The Immunomodulatory Drug Glatiramer Acetate Is Also an Effective Antimicrobial Agent That Kills Gram-Negative Bacteria. Sci. Rep. 2017, 7, 15653. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, C.L.; Broberg, C.A.; McCool, E.N.; Lee, W.J.; Chao, A.; McConnell, E.W.; Pritchard, D.A.; Hebert, M.; Fleeman, R.; Adams, J.; et al. The “PepSAVI-MS” Pipeline for Natural Product Bioactive Peptide Discovery. Anal. Chem. 2017, 89, 1194–1201. [Google Scholar] [CrossRef]
- Defraine, V.; Schuermans, J.; Grymonprez, B.; Govers, S.K.; Aertsen, A.; Fauvart, M.; Michiels, J.; Lavigne, R.; Briers, Y. Efficacy of Artilysin Art-175 against Resistant and Persistent Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2016, 60, 3480–3488. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, L.; Mahdi, N.; Al-Kakei, S.; Musafer, H.; Al-Joofy, I.; Essa, R.; Zwain, L.; Salman, I.; Mater, H.; Al-Alak, S.; et al. Treatment Strategy by Lactoperoxidase and Lactoferrin Combination: Immunomodulatory and Antibacterial Activity against Multidrug-Resistant Acinetobacter Baumannii. Microb. Pathog. 2018, 114, 147–152. [Google Scholar] [CrossRef]
- Morroni, G.; Simonetti, O.; Brenciani, A.; Brescini, L.; Kamysz, W.; Kamysz, E.; Neubauer, D.; Caffarini, M.; Orciani, M.; Giovanetti, E.; et al. In Vitro Activity of Protegrin-1, Alone and in Combination with Clinically Useful Antibiotics, against Acinetobacter Baumannii Strains Isolated from Surgical Wounds. Med. Microbiol. Immunol. 2019, 208, 877–883. [Google Scholar] [CrossRef]
- Vrancianu, C.O.; Popa, L.I.; Bleotu, C.; Chifiriuc, M.C. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Front. Microbiol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; Del Fiol, F.S.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Buckling, A.; Rainey, P.B. Antagonistic Coevolution between a Bacterium and a Bacteriophage. Proc. Biol. Sci. 2002, 269, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage Therapy: An Alternative to Antibiotics in the Age of Multi-Drug Resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G.J. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.R.; March, J.B. Bacteriophages and Biotechnology: Vaccines, Gene Therapy and Antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef]
- Międzybrodzki, R.; Borysowski, J.; Weber-Dąbrowska, B.; Fortuna, W.; Letkiewicz, S.; Szufnarowski, K.; Pawełczyk, Z.; Rogóż, P.; Kłak, M.; Wojtasik, E.; et al. Clinical Aspects of Phage Therapy. Adv. Virus Res. 2012, 83, 73–121. [Google Scholar] [CrossRef]
- Shen, G.-H.; Wang, J.-L.; Wen, F.-S.; Chang, K.-M.; Kuo, C.-F.; Lin, C.-H.; Luo, H.-R.; Hung, C.-H. Isolation and Characterization of Φkm18p, a Novel Lytic Phage with Therapeutic Potential against Extensively Drug Resistant Acinetobacter Baumannii. PLoS ONE 2012, 7, e46537. [Google Scholar] [CrossRef] [Green Version]
- Merabishvili, M.; Vandenheuvel, D.; Kropinski, A.M.; Mast, J.; De Vos, D.; Verbeken, G.; Noben, J.-P.; Lavigne, R.; Vaneechoutte, M.; Pirnay, J.-P. Characterization of Newly Isolated Lytic Bacteriophages Active against Acinetobacter Baumannii. PLoS ONE 2014, 9, e104853. [Google Scholar] [CrossRef] [Green Version]
- Ghajavand, H.; Esfahani, B.N.; Havaei, A.; Fazeli, H.; Jafari, R.; Moghim, S. Isolation of Bacteriophages against Multidrug Resistant Acinetobacter Baumannii. Res. Pharm. Sci. 2017, 12, 373–380. [Google Scholar] [CrossRef]
- Turner, D.; Wand, M.E.; Briers, Y.; Lavigne, R.; Sutton, J.M.; Reynolds, D.M. Characterisation and Genome Sequence of the Lytic Acinetobacter Baumannii Bacteriophage VB_AbaS_Loki. PLoS ONE 2017, 12, e0172303. [Google Scholar] [CrossRef]
- Hernandez-Morales, A.C.; Lessor, L.L.; Wood, T.L.; Migl, D.; Mijalis, E.M.; Cahill, J.; Russell, W.K.; Young, R.F.; Gill, J.J. Genomic and Biochemical Characterization of Acinetobacter Podophage Petty Reveals a Novel Lysis Mechanism and Tail-Associated Depolymerase Activity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.; Luo, T.; Yang, Y.; Dong, D.; Wang, R.; Wang, Y.; Xu, M.; Guo, X.; Hu, F.; He, P. Phage Therapy as a Promising New Treatment for Lung Infection Caused by Carbapenem-Resistant Acinetobacter Baumannii in Mice. Front. Microbiol. 2017, 8, 2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusradze, I.; Karumidze, N.; Rigvava, S.; Dvalidze, T.; Katsitadze, M.; Amiranashvili, I.; Goderdzishvili, M. Characterization and Testing the Efficiency of Acinetobacter Baumannii Phage VB-GEC_Ab-M-G7 as an Antibacterial Agent. Front. Microbiol. 2016, 7, 1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, S.; Huang, G.; Zhang, Y.; Jiang, B.; Yang, Z.; Dong, Z.; You, B.; Yuan, Z.; Hu, F.; Zhao, Y.; et al. Phage Abp1 Rescues Human Cells and Mice from Infection by Pan-Drug Resistant Acinetobacter Baumannii. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 2337–2345. [Google Scholar] [CrossRef]
- Cha, K.; Oh, H.K.; Jang, J.Y.; Jo, Y.; Kim, W.K.; Ha, G.U.; Ko, K.S.; Myung, H. Characterization of Two Novel Bacteriophages Infecting Multidrug-Resistant (MDR) Acinetobacter Baumannii and Evaluation of Their Therapeutic Efficacy in Vivo. Front. Microbiol. 2018, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Feng, Y.; Zong, Z. Two New Lytic Bacteriophages of the Myoviridae Family Against Carbapenem-Resistant Acinetobacter Baumannii. Front. Microbiol. 2018, 9, 850. [Google Scholar] [CrossRef]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A Novel Phage PD-6A3, and Its Endolysin Ply6A3, With Extended Lytic Activity Against Acinetobacter Baumannii. Front. Microbiol. 2018, 9, 3302. [Google Scholar] [CrossRef]
- Jeon, J.; Park, J.-H.; Yong, D. Efficacy of Bacteriophage Treatment against Carbapenem-Resistant Acinetobacter Baumannii in Galleria Mellonella Larvae and a Mouse Model of Acute Pneumonia. BMC Microbiol. 2019, 19, 70. [Google Scholar] [CrossRef]
- Jasim, H.N.; Hafidh, R.R.; Abdulamir, A.S. Formation of Therapeutic Phage Cocktail and Endolysin to Highly Multi-Drug Resistant Acinetobacter Baumannii: In Vitro and in Vivo Study. Iran. J. Basic Med. Sci. 2018, 21, 1100–1108. [Google Scholar] [CrossRef]
- WC, S. Bacteriophage:Early Research. In Bacteriophage: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2005; p. 510. [Google Scholar]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage Treatment of Human Infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter Baumannii Infection. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea With Two Coliphage Preparations: A Randomized Trial in Children From Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef] [Green Version]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.-A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R.; et al. Efficacy and Tolerability of a Cocktail of Bacteriophages to Treat Burn Wounds Infected by Pseudomonas Aeruginosa (PhagoBurn): A Randomised, Controlled, Double-Blind Phase 1/2 Trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Azam, A.H.; Tanji, Y. Bacteriophage-Host Arm Race: An Update on the Mechanism of Phage Resistance in Bacteria and Revenge of the Phage with the Perspective for Phage Therapy. Appl. Microbiol. Biotechnol. 2019, 103, 2121–2131. [Google Scholar] [CrossRef]
- Taylor, V.L.; Fitzpatrick, A.D.; Islam, Z.; Maxwell, K.L. The Diverse Impacts of Phage Morons on Bacterial Fitness and Virulence. Adv. Virus Res. 2019, 103, 1–31. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, L.; Li, X.; Tan, D.; Cong, C.; Xu, Y. Efficacy of a Phage Cocktail in Controlling Phage Resistance Development in Multidrug Resistant Acinetobacter Baumannii. Virus Res. 2019, 272, 197734. [Google Scholar] [CrossRef]
- Chadha, P.; Katare, O.P.; Chhibber, S. In Vivo Efficacy of Single Phage versus Phage Cocktail in Resolving Burn Wound Infection in BALB/c Mice. Microb. Pathog. 2016, 99, 68–77. [Google Scholar] [CrossRef]
- Sneppen, K.; Semsey, S.; Seshasayee, A.S.N.; Krishna, S. Restriction Modification Systems as Engines of Diversity. Front. Microbiol. 2015, 6, 528. [Google Scholar] [CrossRef]
- Van Belleghem, J.D.; Dąbrowska, K.; Vaneechoutte, M.; Barr, J.J.; Bollyky, P.L. Interactions between Bacteriophage, Bacteria, and the Mammalian Immune System. Viruses 2018, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, D.; Taha, M.; Yi, Q.; Ramirez-Arcos, S.; Diallo, J.-S.; Carli, A.; Abdelbary, H. Does Treatment Order Matter? Investigating the Ability of Bacteriophage to Augment Antibiotic Activity against Staphylococcus Aureus Biofilms. Front. Microbiol. 2018, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide Sequence of the Iap Gene, Responsible for Alkaline Phosphatase Isozyme Conversion in Escherichia Coli, and Identification of the Gene Product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-Y.; Kao, C.-Y.; Lin, W.-H.; Zheng, P.-X.; Yan, J.-J.; Wang, M.-C.; Teng, C.-H.; Tseng, C.-C.; Wu, J.-J. Characterization of CRISPR-Cas Systems in Clinical Klebsiella Pneumoniae Isolates Uncovers Its Potential Association With Antibiotic Susceptibility. Front. Microbiol. 2018, 9, 1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, A.B.; Henriksen, E.D.; Stout, E.; Brandt, K.; Barrangou, R. Characterizing the Activity of Abundant, Diverse and Active CRISPR-Cas Systems in Lactobacilli. Sci. Rep. 2018, 8, 11544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, A.; Shi, Y.; Kage-Nakadai, E.; Mitani, S.; Xue, D. Caspase-Dependent Conversion of Dicer Ribonuclease into a Death-Promoting Deoxyribonuclease. Science 2010, 328, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, F.C.; Hatoum-Aslan, A. Conjugation Assay for Testing CRISPR-Cas Anti-Plasmid Immunity in Staphylococci. Bio-Protocol 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR Interference Limits Horizontal Gene Transfer in Staphylococci by Targeting DNA. Science 2008, 322, 1843–1845. [Google Scholar] [CrossRef] [Green Version]
- Di Nocera, P.P.; Rocco, F.; Giannouli, M.; Triassi, M.; Zarrilli, R. Genome Organization of Epidemic Acinetobacter Baumannii Strains. BMC Microbiol. 2011, 11, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, Y.; Soler, C.; Jault, P.; Mérens, A.; Gérome, P.; Nab, C.M.; Trueba, F.; Bargues, L.; Thien, H.V.; Vergnaud, G.; et al. Diversity of Acinetobacter Baumannii in Four French Military Hospitals, as Assessed by Multiple Locus Variable Number of Tandem Repeats Analysis. PLoS ONE 2012, 7, e44597. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.R.; Lee, S.Y. CRISPR Technologies for Bacterial Systems: Current Achievements and Future Directions. Biotechnol. Adv. 2016, 34, 1180–1209. [Google Scholar] [CrossRef]
- Karah, N.; Samuelsen, Ø.; Zarrilli, R.; Sahl, J.W.; Wai, S.N.; Uhlin, B.E. CRISPR-Cas Subtype I-Fb in Acinetobacter Baumannii: Evolution and Utilization for Strain Subtyping. PLoS ONE 2015, 10, e0118205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-Guided Editing of Bacterial Genomes Using CRISPR-Cas Systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Citorik, R.J.; Mimee, M.; Lu, T.K. Sequence-Specific Antimicrobials Using Efficiently Delivered RNA-Guided Nucleases. Nat. Biotechnol. 2014, 32, 1141–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikard, D.; Euler, C.W.; Jiang, W.; Nussenzweig, P.M.; Goldberg, G.W.; Duportet, X.; Fischetti, V.A.; Marraffini, L.A. Exploiting CRISPR-Cas Nucleases to Produce Sequence-Specific Antimicrobials. Nat. Biotechnol. 2014, 32, 1146–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Cho, D.-H.; Park, M.; Chung, W.-J.; Shin, D.; Ko, K.S.; Kweon, D.-H. CRISPR/Cas9-Mediated Re-Sensitization of Antibiotic-Resistant Escherichia Coli Harboring Extended-Spectrum β-Lactamases. J. Microbiol. Biotechnol. 2016, 26, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; He, T.; Zhang, L.; Pang, M.; Zhang, Q.; Zhou, Y.; Bao, H.; Wang, R. Generation of Newly Discovered Resistance Gene Mcr-1 Knockout in Escherichia Coli Using the CRISPR/Cas9 System. J. Microbiol. Biotechnol. 2017, 27, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Jiang, Y.; Shao, L.; Yang, P.; Sun, B.; Yang, S.; Chen, D. CRISPR/Cas9-Based Efficient Genome Editing in Staphylococcus Aureus. Acta Biochim. Biophys. Sin. 2017, 49, 764–770. [Google Scholar] [CrossRef] [Green Version]
- So, Y.; Park, S.-Y.; Park, E.-H.; Park, S.-H.; Kim, E.-J.; Pan, J.-G.; Choi, S.-K. A Highly Efficient CRISPR-Cas9-Mediated Large Genomic Deletion in Bacillus Subtilis. Front. Microbiol. 2017, 8, 1167. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Xiang, H.; Mu, D.; Wang, D.; Wang, T. Exploiting a Conjugative CRISPR/Cas9 System to Eliminate Plasmid Harbouring the Mcr-1 Gene from Escherichia Coli. Int. J. Antimicrob. Agents 2019, 53, 1–8. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Chen, Y.; Hua, X.; Yu, Y.; Ji, Q. A Highly Efficient CRISPR-Cas9-Based Genome Engineering Platform in Acinetobacter Baumannii to Understand the H(2)O(2)-Sensing Mechanism of OxyR. Cell Chem. Biol. 2019, 26, 1732–1742.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangas, E.L.; Rubio, A.; Álvarez-Marín, R.; Labrador-Herrera, G.; Pachón, J.; Pachón-Ibáñez, M.E.; Divina, F.; Pérez-Pulido, A.J. Pangenome of Acinetobacter Baumannii Uncovers Two Groups of Genomes, One of Them with Genes Involved in CRISPR/Cas Defence Systems Associated with the Absence of Plasmids and Exclusive Genes for Biofilm Formation. Microb. Genom. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Karlapudi, A.P.; Venkateswarulu, T.C.; Tammineedi, J.; Srirama, K.; Kanumuri, L.; Prabhakar Kodali, V. In Silico SgRNA Tool Design for CRISPR Control of Quorum Sensing in Acinetobacter Species. Genes Dis. 2018, 5, 123–129. [Google Scholar] [CrossRef]
- Zhang, F.; Wen, Y.; Guo, X. CRISPR/Cas9 for Genome Editing: Progress, Implications and Challenges. Hum. Mol. Genet. 2014, 23, R40–R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozzuto, G.; Molinari, A. Liposomes as Nanomedical Devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taranejoo, S.; Liu, J.; Verma, P.; Hourigan, K. A Review of the Developments of Characteristics of PEI Derivatives for Gene Delivery Applications. J. Appl. Polym. Sci. 2015, 132. [Google Scholar] [CrossRef]
- Terwee, C.B.; Roorda, L.D.; Dekker, J.; Bierma-Zeinstra, S.M.; Peat, G.; Jordan, K.P.; Croft, P.; de Vet, H.C.W. Mind the MIC: Large Variation among Populations and Methods. J. Clin. Epidemiol. 2010, 63, 524–534. [Google Scholar] [CrossRef]
- Çağlan, E.; Nigiz, Ş.; Sancak, B.; Gür, D. Resistance and Heteroresistance to Colistin among Clinical Isolates of Acinetobacter Baumannii. Acta Microbiol. Immunol. Hung. 2019, 1–5. [Google Scholar] [CrossRef]
- Mouton, J.W.; Muller, A.E.; Canton, R.; Giske, C.G.; Kahlmeter, G.; Turnidge, J. MIC-Based Dose Adjustment: Facts and Fables. J. Antimicrob. Chemother. 2018, 73, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical Relevance of Bacteriostatic versus Bactericidal Mechanisms of Action in the Treatment of Gram-Positive Bacterial Infections. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, G.; Hoen, B.; Tornos, P.; Thuny, F.; Prendergast, B.; Vilacosta, I.; Moreillon, P.; de Jesus Antunes, M.; Thilen, U.; Lekakis, J.; et al. Guidelines on the Prevention, Diagnosis, and Treatment of Infective Endocarditis (New Version 2009): The Task Force on the Prevention, Diagnosis, and Treatment of Infective Endocarditis of the European Society of Cardiology (ESC). Endorsed by the Europea. Eur. Heart J. 2009, 30, 2369–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nau, R.; Djukic, M.; Spreer, A.; Ribes, S.; Eiffert, H. Bacterial Meningitis: An Update of New Treatment Options. Expert Rev. Anti-Infect. Ther. 2015, 13, 1401–1423. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C.P.; Van Anglen, L.J.; Dretler, R.H.; Adams, J.S.; Prokesch, R.C.; Luu, Q.; Krinsky, A.H. Outpatient Treatment of Osteomyelitis with Telavancin. Int. J. Antimicrob. Agents 2017, 50, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Rafailidis, P.I.; Kouranos, V.D.; Christodoulou, C.; Falagas, M.E. Linezolid for Patients with Neutropenia: Are Bacteriostatic Agents Appropriate? Expert Rev. Anti-Infect. Ther. 2009, 7, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Nau, R.; Eiffert, H. Modulation of Release of Proinflammatory Bacterial Compounds by Antibacterials: Potential Impact on Course of Inflammation and Outcome in Sepsis and Meningitis. Clin. Microbiol. Rev. 2002, 15, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Halfawy, O.M.; Valvano, M.A. Antimicrobial Heteroresistance: An Emerging Field in Need of Clarity. Clin. Microbiol. Rev. 2015, 28, 191–207. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.E.; Sherman, E.X.; Weiss, D.S.; Rather, P.N. Aminoglycoside Heteroresistance in Acinetobacter Baumannii AB5075. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-Y.; Chen, C.-L.; Wang, S.-B.; Su, L.-H.; Chen, S.-H.; Liu, S.-Y.; Wu, T.-L.; Lin, T.-Y.; Chiu, C.-H. Imipenem Heteroresistance Induced by Imipenem in Multidrug-Resistant Acinetobacter Baumannii: Mechanism and Clinical Implications. Int. J. Antimicrob. Agents 2011, 37, 302–308. [Google Scholar] [CrossRef]
- Hung, K.-H.; Wang, M.-C.; Huang, A.-H.; Yan, J.-J.; Wu, J.-J. Heteroresistance to Cephalosporins and Penicillins in Acinetobacter Baumannii. J. Clin. Microbiol. 2012, 50, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Rayner, C.R.; Nation, R.L.; Owen, R.J.; Spelman, D.; Tan, K.E.; Liolios, L. Heteroresistance to Colistin in Multidrug-Resistant Acinetobacter Baumannii. Antimicrob. Agents Chemother. 2006, 50, 2946–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sola, C.; Lamberghini, R.O.; Ciarlantini, M.; Egea, A.L.; Gonzalez, P.; Diaz, E.G.; Huerta, V.; Gonzalez, J.; Corso, A.; Vilaro, M.; et al. Heterogeneous Vancomycin-Intermediate Susceptibility in a Community-Associated Methicillin-Resistant Staphylococcus Aureus Epidemic Clone, in a Case of Infective Endocarditis in Argentina. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, J.S.; Murray, C.K.; Jorgensen, J.H. Colistin Heteroresistance in Acinetobacter and Its Association with Previous Colistin Therapy. Antimicrob. Agents Chemother. 2008, 52, 351–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| A. baumannii Isolates Number | AR Profile | Minimal Inhibitory Concentration (MIC) Range | Resistant Isolates (%) | Hospital Wards/Origin | References |
|---|---|---|---|---|---|
| 164 | GEN, AMK | 256–≥1024 mg/L | – – 32.1%–83.8% | – – Endotracheal aspirates, tracheal secretion, wound tampon, pus, sputum, catheters/ ICUs (Intensive Units Care) | [14] |
| Cephalosporins, Carbapenems | – U a | ||||
| 44 | PIP, CTX, CAZ/ATM, IMP | 0.5–256 mg/L | 79.5% | U | [15] |
| 121 | CIP, AMK, AMP–SUL, ATM, CTX, GEN, NET, PIP, TIM, TOB, IMP | 0.5–256 mg/L | 0%–92.6% | Bronchial cultures, burns, blood culture, catheters/ ICUs | [16] |
| 375 | AMP, PIP, PTZ, CAZ, CTX, IMP, MEM, GEN, AMK, CIP | 0.5–>256 mg/L | 0%–100% | Sputum, wounds/ ICUs | [17] |
| 23 | CAZ, CTX, FEP, PIP, TZP b, ATM, IMP, MEM, CIP, AMK, GEN, SXT | 0.75–>256 (µg mL−1) b | 100% | U | [32] |
| 72 | IMP, MEM, FEP, CAZ, SUL, CFP–SUL, PIP, TZP, ATM, CIP, AMK, TIG | 4–≥512 mg/L | 77.8% | ICUs – | [36] |
| 100 | IMP, MEM, CAZ, CST, TET, TIG, AMP–SUL | 0.25–256 (µg/mL) b | 0%–100% | Sputum, wounds, blood culture, urine, fluids, hemodialysis catheters; ACH (Acute Care Hospital) | [37] |
| 204 | TIC, TIM, CAZ, FEP, ATM, IMP, TOB, KM, GEN, AMK, PFX, LVX, OFX, CST, TET, FOS | U | 92% for β-lactams; 47% for IMP | Puncture, pus, blood culture samples/ICUs | [41] |
| 20 | TIC, CAZ, IMP, MEM | 4–>256 µg/mL | 90%–100% | Tracheal aspirate, bile, urine, burns, respiratory tract, blood culture, sputum/ICUs | [38] |
| 100 | PIP, AMP/SUL, CIP, AMK, IMP, CTX, FEP, CRO, TET, GEN | U | 45%–100% | Burns, sputum, tracheal secretion, pleural fluid, blood culture, urine, cerebrospinal fluid/ ICU | [42] |
| 834 | AMC, CIP, GEN, AMK, CFP–SUL, IMP, MEM | U | 71.7%–96.8% | Tracheal aspirate, blood culture, urine, wounds/ICU | [43] |
| 15 | IMP, MEM, GEN, GEN/NMP, CIP, CIP/NMP, CAZ | 0.25–≥256 mg/L | U | Gastroenterology | [44] |
| 155 | CIP, FEP, TZP, CAZ, IMP, MEM, AMP/SUL | U | 93.5%–94.4% | Blood respiratory tract secretions, catheter, urine | [39] |
| 65 | AMK, AMP/SUL, FEP, CST, SXT, DOX, IMP, LVX, MEM, MIN, SUL, TIG, TOB | 0.125–256 mg/L | 1.5%–96.9% | Respiratory tract secretions | [24] |
| 59 | TIC, PIP, AMP/SUL, TZP, CAZ, FEP, IMP, MEM, CST, GEN, TOB, AMK, MIN, CIP, LVX, SXT | U | 10.17%–96.61% | CCNU (critical care nursing unit); UPU (emergency unit); RE (resuscitation); IM (internal medicine); NE (Nephrology); HO (hematology/Oncology); TC (thoracic cardiology); GS (general surgery); NEO (neonatology) | [25] |
| 15 | TIC, TIM, PIP, TZP, CAZ, CTX, FEP, MEM, NAL, LVX, CIP, GEN, TOB, AMK, SXT, IMP | U (exception MEM and IMP–32 mg/L) | 40%–100% | Burns, pleural fluid, urine, bronchoalveolar lavage, pus, blood culture | [26] |
| 40 | CAZ IMP GEN | U | 92.5% 85% 70% | ICUs, IM, GS | [27] |
| 41 | AMP/SUL, TZP, CAZ, CRO, FEP, IMP, MEM, GEN, CIP, TIG, AMK | 4–≥128 mg/L | 34.1%–100% | Tracheal aspirate, burns, urine, blood, exudate/ICU | [28] |
| 84 | GEN, AMK, CRO, FEP, CIP, LVX, CAZ, IMP, MEM, PMB, CST, AMP, TET, TIG, ATM | U | 0%–100% | Wounds | [30] |
| 35 | Class III Cephalosporins Class II Cephalosporins Carbapenems Aminoglycosides Fluoroquinolones | U | >90% 65% >90% 60% 90% | Tracheal secretion, urine, cerebrospinal fluid/ICU | [31] |
| 8 | CAZ, FEP, CTX, CRO, IMP MEM, TZP, AMP/SUL, GEN, CIP, TIG, CST | 1–>128 mg/L | 71.4%–100% | Blood cultures, urine samples, aspirate sputum, bronchoalveolar lavage fluid, wound swab, pus | [21] |
| 41 | Aminoglycosides Β-lactams Quinolones Tetracyclines | U | 95% | Tracheal aspirate, peritoneal fluid, bronchial lavage/ ICU; UPU; NICU (Neonatal Intensive Care Unit) | [22] |
| Organism | AMP | Type of Study | Animal Model | Main Results | References |
|---|---|---|---|---|---|
| NA | ZY4 cathelicidin-BF-15 derived | in vitro; in vivo | mouse septicemia infection model | Antibacterial activity in plasma; biofilm inhibition; kills persister cells; inhibition of infection and inflammation in vivo | [244] |
| NA | epsilon-poly L-lysine (EPL)-catechol | in vitro; in vivo | mouse burn wounds infection model | Reducing bacterial burden in vivo | [263] |
| Vespa affinis | mastoparan-AF | in vitro | NA | Potent antimicrobial activity | [247] |
| NA | chex1-Arg20 amide (ARV-1502) | in vivo | Mouse infection model | Reduction of bacterial load | [264] |
| NA | α-helical -26 AMP residues | in vitro | NA | Great antimicrobial activity | [265] |
| Delftia spp. | delfibactin A | in vitro | NA | Great inhibitory effects | [266] |
| Capra hircus | mini-ChBac7.5Nα mini-ChBac7.5Nβ | in vitro | NA | Significant antimicrobial activity; induce membrane damages; | [248] |
| Hybrid striped bass Morone saxatilis × M. chrysops | I16 K-piscidine-1 analog | in vitro; in vivo | Sepsis mouse model | Strong bactericidal activity; high survival rate of infected mice; | [267] |
| Musca domestica | cecropin-4 | in vitro | NA | Great bactericidal activity against MRAB and PRAB; inhibits biofilm formation | [252] |
| NA | Ω17 and Ω76 family peptides | in vitro; in vivo | Mouse peritoneal infection model | Disrupt bacterial membranes; induce small-molecule leakage; rapid bactericidal activity; | [268] |
| NA | ceragenins (AMP synthetic mimics) | in vitro | NA | Antibiofilm activity; inhibitory effects | [269] |
| Medicago truncatula | nodule-specific cysteine-rich (NCR) peptide and its derivatives | in vitro | NA | Potent killer of pathogenic bacteria | [270] |
| NA | TAT-RasGAP317−326 anticancer peptide | in vitro; in vivo | Mousel model of lethal peritonitis | Growth inhibition effects; broad-spectrum antimicrobial activity; great efficacy in vivo | [271] |
| NA | WLBU2-cationic amphipathic peptide | in vitro | NA | Eradicating bacterial biofilms; | [272] |
| Myxine glutinosa L. | myxinidin 2; myxinidin 3 | in vitro, in vivo | Mouse skin wounds infection model | Antibiofilm activity; anti-inflammatory activity; enhance wound healing; | [253] |
| Hepatitis B virus | D-150–177C, HBcARD derivative peptide | in vivo | Mouse sepsis infection model | Strong bactericidal activity; 90% of mice protected from death; | [273] |
| Pisum sativum | nuripep 1653 | in vitro | NA | Significant antimicrobial activity; | [274] |
| Cimex lectularius (bedbug) | CL defensin | in vitro | NA | Inducing membrane depolarization and pore forming; bactericidal action | [275] |
| Bungarus fasciatus | cathelicidin—BF derivate (Cath-A) | in vitro | NA | Bacterial growth inhibition | [245] |
| Lucilia sericata | LS-sarcotoxin and LS-stomoxyn | in vitro; in vivo | Mouse model infection | Strong activity against GRAM-NEGATIVE; | [249] |
| Leiurus quinquestriatus | venom cocktail proteins | in vitro | NA | Broad-spectrum antimicrobial activity; growth inhibition; | [246] |
| Myrmecia pilosula | Δ-Myrtoxin-Mp1a (Mp1a) heterodimeric peptide | in vitro; in vivo | Mouse model | Antibacterial activity; significant potency; nociceptive pain upon injection into mice | [243] |
| NA | glatiramer acetate | in vitro | NA | Efficient killing of clinical isolates | [276] |
| King cobra | OH-CATH30 D-OH-CATH30 | in vitro; in vivo | Mouse model | Strong inhibition activity; low toxicity, great immunogenicity; | [242] |
| NA | stapled AMP Mag(i+4)1,15(A9 K, B21A, N22 K, S23 K) | in vitro; in vivo | Mouse peritonitis sepsis model | Great bactericidal activity; 88% of mice cured after intraperitoneal injection; | [255] |
| Viola odorata | Cy02 (cyclotide) | in vitro | NA | Strong bactericidal action | [277] |
| P. aeruginosa bacteriophage | artilysin 175 | in vitro | NA | High, rapid and broad antibacterial activity against MRAB | [278] |
| Calliphora vicina | FLIP 7 | in vitro | NA | Antibiofilm activity | [254] |
| Camel (colostrum milk) | lactoperoxidase lactoferrin | in vitro; in vivo | Acute pneumonia mouse model | Major inhibition effects; significant clearance of A. baumannii in lung and blood culture; | [279] |
| Rana catesbeiana | ranalexin danalexin | in vitro | NA | Strong antimicrobial activity | [250] |
| NA | PNA (RXR)4 XB | in vitro; in vivo | Galleria mellonella sepsis model | Excellent bactericidal activity in vitro; high dose of PNA conjugate required in sepsis model | [256] |
| NA | protegrin-1 | in vitro | NA | Good activity against MRAB; no antibiofilm activity; | [280] |
| NA | aurein 1.2, CAMEL, citropin 1.1., LL-37, omiganan, r-omiganan, pexiganan and temporin A | in vitro | NA | CAMEL and pexiganan displayed the highest antibacterial activity | [251] |
| Phages | Family | Isolation Source | Type of Study | Number of Tested Strains | % of Susceptible Strains | Animal Model Application | References |
|---|---|---|---|---|---|---|---|
| økm18p | Corticoviridae | hospital sewage | in vitro | 34 MDR, 16 of those XDRAB | 44.1% | NA | [288] |
| Acibel004 | Myoviridae | wastewater sample | in vitro | 34 MDR | 82.3% | NA | [289] |
| Acibel007 | Podoviridae | wastewater sample | in vitro | 34 MDR | 82.3% | NA | |
| IsfAB78 | Myoviridae | water sample | in vitro | 43 MDR | 27.9% | NA | [290] |
| IsfAB39 | Podoviridae | water sample | in vitro | 43 MDR | 25.5% | NA | |
| vB_AbaS_Loki | Siphoviridae | sludge | in vitro | 34 | 5.8% | NA | [291] |
| Petty phage | Podoviridae | sewage | in vitro | 40, 25 of those MDR | 10% | NA | [292] |
| SH-Ab 15599 | Myoviridae | sewage | in vitro | 48 CRAB | 27% | NA | [293] |
| SH-Ab15708 | Myoviridae | sewage | in vitro | 48 CRAB | 29.1% | NA | |
| SH-Ab15497 | Siphoviridae | sewage | in vitro | 48 CRAB | 29.1% | NA | |
| SH-Ab15519 | Podoviridae | sewage | in vivo | 48 CRAB | 16.6% | Mouse model—lung infection; 90% survival rate | |
| vBGEC_AbM-G7was (phiG7) | Myoviridae | sewage | in vivo | 200 | 68% | Rats wound model; 100% survival rate | [294] |
| Abp1 | Moraxelaceae | sewage | in vitro | 20 | NA | Hella cells infection protection assay; 100% protection and survival rate of Hella cells. | [295] |
| in vivo | 20 | Mouse local and systemic infection model; 100% survival rate. | |||||
| PB AB08 | Myoviridae | Bacteriophage Bank of Korea | in vivo | 14 MDR | 35.7% | Mice model—intranasal phage cocktail; 35% survival rate | [296] |
| PBAB25 | Myoviridae | Bacteriophage Bank of Korea | in vivo | 14 MDR | 7.1% | Mice model—ntranasal phage cocktail; 35% survival rate. | |
| WCHABP1 | Myoviridae | hospital sewage | in vivo | 2 CRAB | NA | Galleria mellonela infection model; 75% survival rate after phage administration | [297] |
| WCHABP12 | Myoviridae | hospital sewage | in vivo | NA | |||
| PD-6A3 | Podoviridae | sewage | in vivo | 552 MDR | 32.4% | Sepsis mouse model; intraperitoneal administration; endolysin therapy, endolysin + phage therapy, phage therapy and phage cocktail; 70%, 70%, 60% and 50% survival rate. | [298] |
| Bϕ-R2096 | Myoviridae | hospital sewage | in vivo | 20 CRAB | NA | Galleria mellonella infection model; 80% and 50% survival rate at 96 and 48 h. | [299] |
| in vivo | NA | Mouse model acute pneumonia; 100%, 60% and 30% survival rate at day 12, with MOI 10, 1 and 0.1 | |||||
| AB3P1 | NA | sewage, farm soil, feces of sheep, chicken litter, swab for surgical lounge. | in vivo | 23 | 78.2% | Mice model; intraperitoneal administration of AB3 phages; 100% survival rate; | [300] |
| AB3P2 | NA | in vivo | |||||
| AB3P3 | NA | in vivo | |||||
| AB3P4 | NA | in vivo |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii. Microorganisms 2020, 8, 935. https://doi.org/10.3390/microorganisms8060935
Vrancianu CO, Gheorghe I, Czobor IB, Chifiriuc MC. Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii. Microorganisms. 2020; 8(6):935. https://doi.org/10.3390/microorganisms8060935
Chicago/Turabian StyleVrancianu, Corneliu Ovidiu, Irina Gheorghe, Ilda Barbu Czobor, and Mariana Carmen Chifiriuc. 2020. "Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii" Microorganisms 8, no. 6: 935. https://doi.org/10.3390/microorganisms8060935