Lanthanide-Dependent Methanol and Formaldehyde Oxidation in Methylobacterium aquaticum Strain 22A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Construction of Mutant Strains
2.3. Genome Resequencing of ΔxoxF1Sup and re-mxaFSup
2.4. Expression Vector Construction
2.5. qPCR
2.6. Purification of Recombinant XoxF1
2.7. MDH Activity and Enzyme Kinetics
2.8. S-hydroxymethyl Glutathione Dehydrogenase Assay
2.9. Statistical Analysis
3. Results
3.1. Phylogenetic Analysis MDH-Like Proteins of M. aquaticum Strain 22A
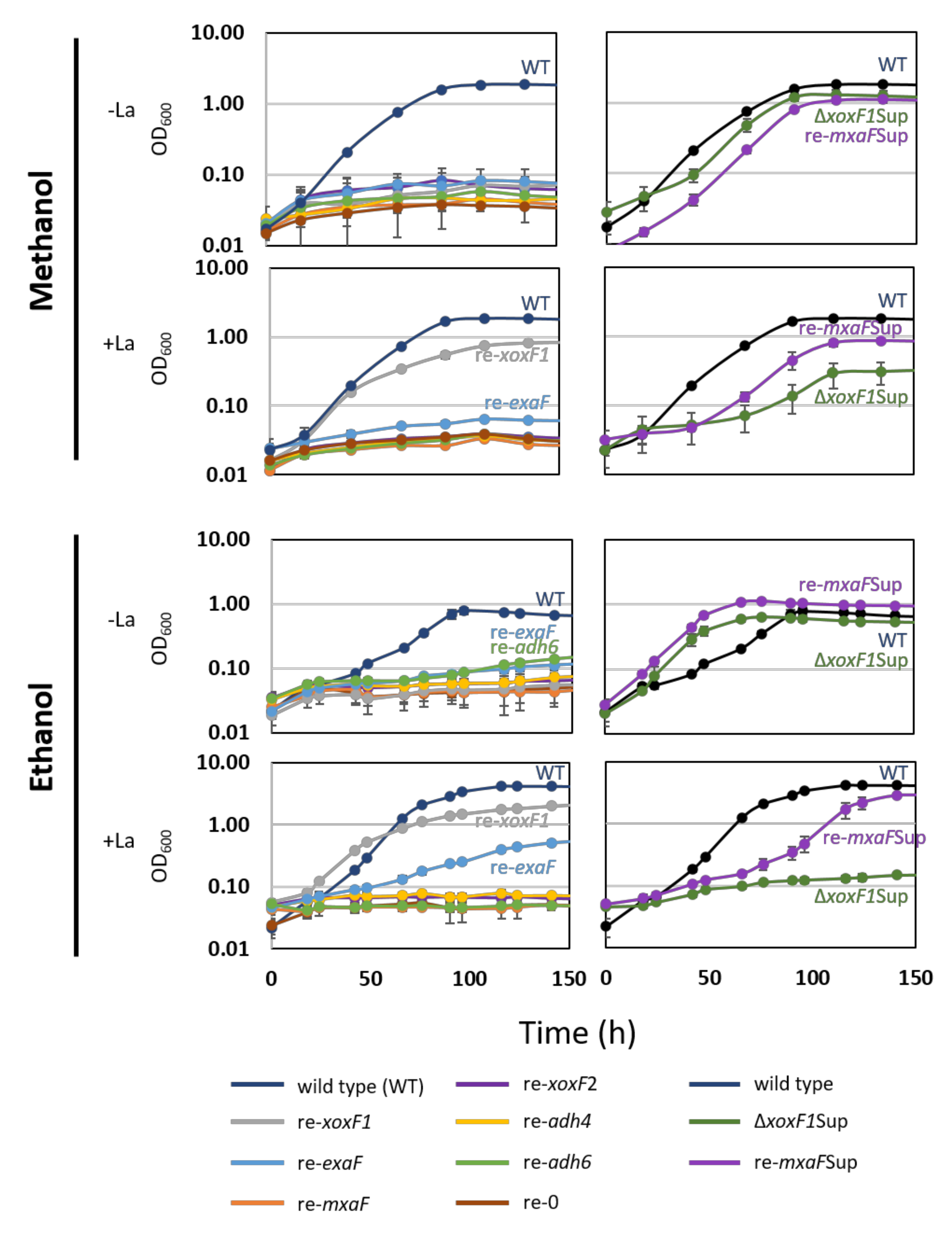
3.2. The Phenotype of MDH-Like Gene Mutants
3.3. Mutations Detected in the Suppression Mutants
3.4. ΔmxbD Phenotype
3.5. The Recombinant XoxF1-His Oxidizes Methanol and Formaldehyde
3.6. hgd Encodes S-hydroxymethylglutathione Dehydrogenase
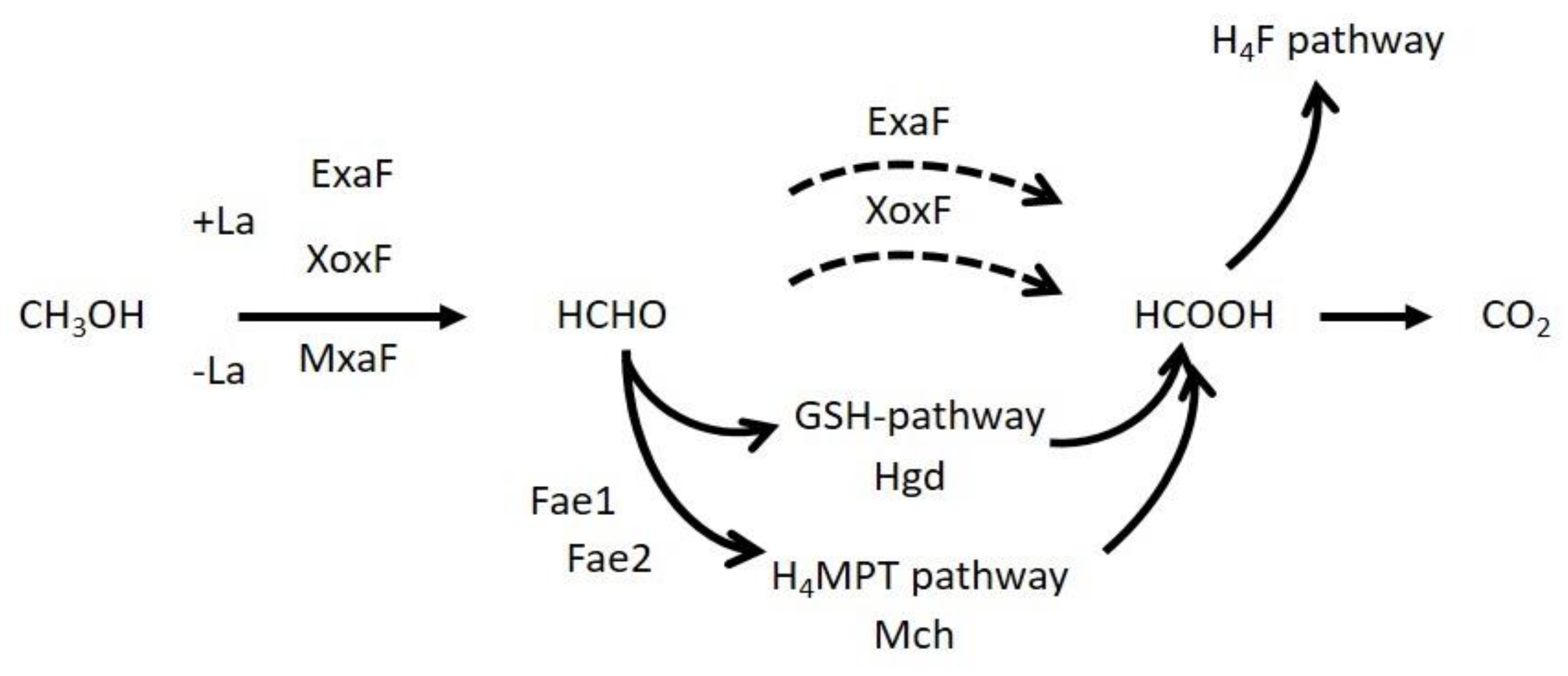
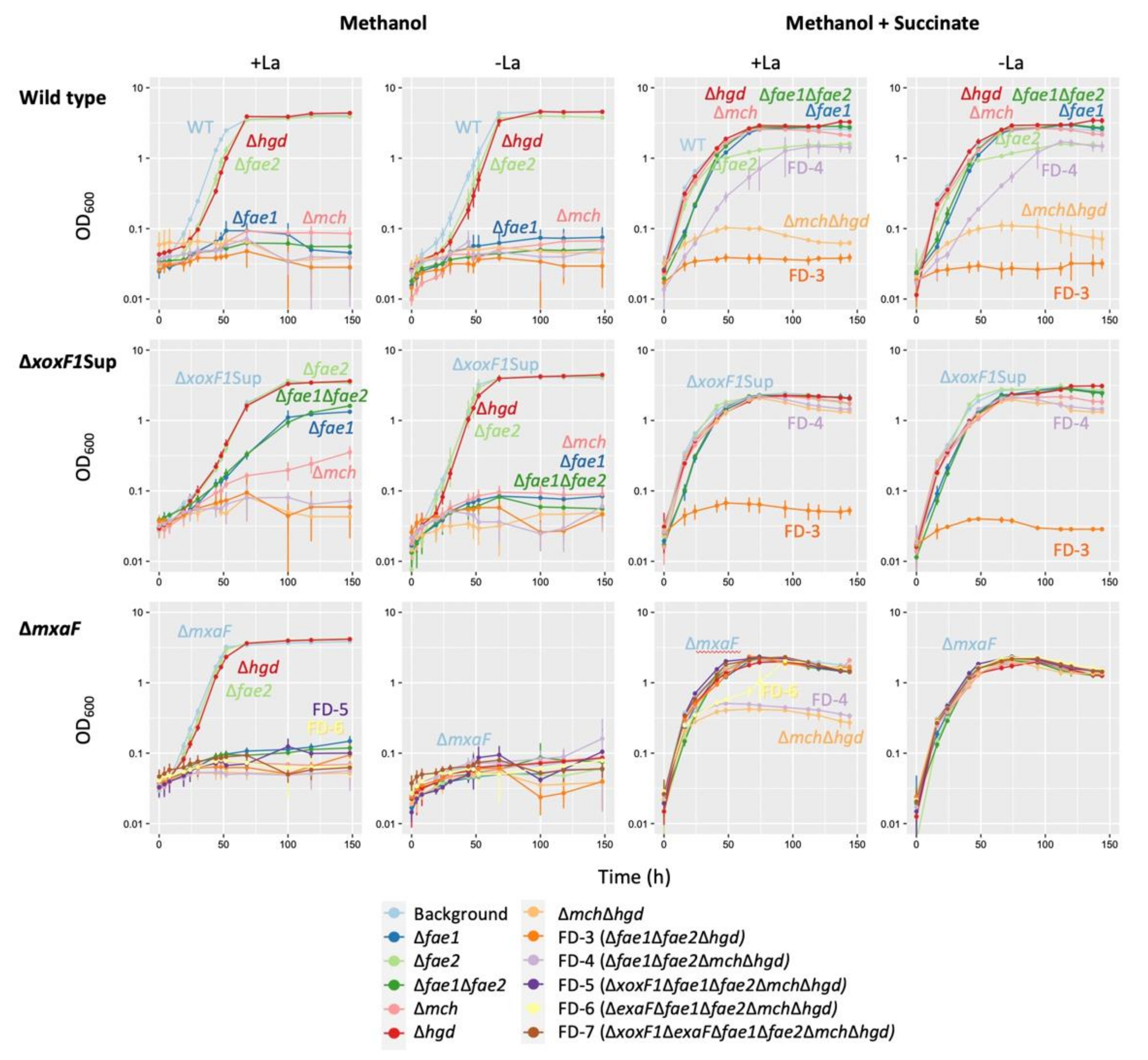
3.7. Characterization of the Mutants in Formaldehyde Oxidation Pathways
4. Discussion
4.1. Identification of PQQ-ADH Genes for Methylotrophy in Strain 22A
4.2. The Mutation in MxbD de-repress mxaF
4.3. Formaldehyde Oxidation by XoxF1 and ExaF
4.4. The Role of the GSH Pathway
4.5. The Role of H4MPT Pathway in Strain 22A
4.6. Overall Methanol Metabolism in Strain 22A
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fall, R.; Benson, A.A. Leaf methanol the simplest natural product from plants. Trends Plant Sci. 1996, 1, 296–301. [Google Scholar] [CrossRef]
- Anthony, C. The Biochemistry of Methylotrophs; Academic Press: London, UK, 1982. [Google Scholar]
- Schmidt, S.; Christen, P.; Kiefer, P.; Vorholt, J.A. Functional investigation of methanol dehydrogenase-like protein XoxF in Methylobacterium extorquens AM1. Microbiology 2010, 156, 2575–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of methylotrophs with plants and other heterotrophic bacteria. Microorganisms 2015, 3, 137–151. [Google Scholar] [CrossRef]
- Green, P.N.; Ardley, J.K. Review of the genus Methylobacterium and closely related organisms: A proposal that some Methylobacterium species be reclassified into a new genus, Methylorubrum gen. Nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 2727–2748. [Google Scholar] [CrossRef]
- Chistoserdova, L.; Chen, S.-W.; Lapidus, A.; Lidstrom, M.E. Methylotrophy in Methylobacterium extorquens AM1 from a genomic point of view. J. Bacteriol. 2003, 185, 2980–2987. [Google Scholar] [CrossRef] [Green Version]
- Anthony, C. How half a century of research was required to understand bacterial growth on C1 and C2 compounds; the story of the serine cycle and the ethylmalonyl-CoA pathway. Sci. Prog. 2011, 94, 109–137. [Google Scholar] [CrossRef]
- Anthony, C.; Williams, P. The structure and mechanism of methanol dehydrogenase. BBA-Proteins Proteom. 2003, 1647, 18–23. [Google Scholar] [CrossRef]
- Marx, C.J.; Chistoserdova, L.; Lidstrom, M.E. Formaldehyde-detoxifying role of the tetrahydromethanopterin-linked pathway in Methylobacterium extorquens AM1. J. Bacteriol. 2003, 185, 7160–7168. [Google Scholar] [CrossRef] [Green Version]
- Peyraud, R.; Schneider, K.; Kiefer, P.; Massou, S.; Vorholt, J.A.; Portais, J.-C. Genome-scale reconstruction and system level investigation of the metabolic network of Methylobacterium extorquens AM1. BMC Syst. Biol. 2011, 5, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A.; Marx, C.J.; Lidstrom, M.E.; Thauer, R.K. Novel formaldehyde-activating enzyme in Methylobacterium extorquens AM1 required for growth on methanol. J. Bacteriol. 2000, 182, 6645–6650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowther, G.J.; Kosaly, G.; Lidstrom, M.E. Formate as the main branch point for methylotrophic metabolism in Methylobacterium extorquens AM1. J. Bacteriol. 2008, 190, 5057–5062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, M.; Anthony, C.; Harlos, K.; Goodwin, M.G.; Blake, C. The refined structure of the quinoprotein methanol dehydrogenase from Methylobacterium extorquens at 1.94 Å. Structure 1995, 3, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Hibi, Y.; Asai, K.; Arafuka, H.; Hamajima, M.; Iwama, T.; Kawai, K. Molecular structure of La3+-induced methanol dehydrogenase-like protein in Methylobacterium radiotolerans. J. Biosci. Bioeng. 2011, 111, 547–549. [Google Scholar] [CrossRef]
- Nakagawa, T.; Mitsui, R.; Tani, A.; Sasa, K.; Tashiro, S.; Iwama, T.; Hayakawa, T.; Kawai, K. A catalytic role of XoxF1 as La3+-dependent methanol dehydrogenase in Methylobacterium extorquens strain AM1. PLoS ONE 2012, 7, e50480. [Google Scholar] [CrossRef] [Green Version]
- Pol, A.; Barends, T.R.M.; Dietl, A.; Khadem, A.F.; Eygensteyn, J.; Jetten, M.S.M.; Op den Camp, H.J.M. Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environ. Microbiol. 2014, 16, 255–264. [Google Scholar] [CrossRef]
- Good, N.M.; Moore, R.S.; Suriano, C.J.; Martinez-Gomez, N.C. Contrasting in vitro and in vivo methanol oxidation activities of lanthanide-dependent alcohol dehydrogenases XoxF1 and ExaF from Methylobacterium extorquens AM1. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Masuda, S.; Fujitani, Y.; Sahin, N.; Tani, A. Oharaeibacter diazotrophicus gen. Nov., sp. Nov., a diazotrophic and facultatively methylotrophic bacterium, isolated from rice rhizosphere. Int. J. Syst. Evol. Microbiol. 2017, 67, 576–582. [Google Scholar] [CrossRef]
- Lv, H.; Sahin, N.; Tani, A. Isolation and genomic characterization of Novimethylophilus kurashikiensis gen. Nov. Sp. Nov., a new lanthanide-dependent methylotrophic species of methylophilaceae. Environ. Microbiol. 2018, 20, 1204–1223. [Google Scholar] [CrossRef]
- Lv, H.; Tani, A. Genomic characterization of methylotrophy of Oharaeibacter diazotrophicus strain SM30T. J. Biosci. Bioeng. 2018, 126, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Fitriyanto, N.A.; Fushimi, M.; Matsunaga, M.; Pertiwiningrum, A.; Iwama, T.; Kawai, K. Molecular structure and gene analysis of Ce3+-induced methanol dehydrogenase of Bradyrhizobium sp. MAFF211645. J. Biosci. Bioeng. 2011, 111, 613–617. [Google Scholar] [CrossRef]
- Howat, A.M.; Vollmers, J.; Taubert, M.; Grob, C.; Dixon, J.L.; Todd, J.D.; Chen, Y.; Kaster, A.-K.; Murell, J.C. Comparative genomics and mutational analysis reveals a novel XoxF-utilizing methylotroph in the Roseobacter group isolated from the marine environment. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Farhan Ul Haque, M.; Kalidass, B.; Bandow, N.; Turpin, E.A.; DiSpirito, A.A.; Semrau, J.D. Cerium regulates expression of alternative methanol dehydrogenase in Methylosinus trichosporium OB3b. Appl. Environ. Microb. 2015, 81, 7546–7552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, F.; Lidstrom, M.E. XoxF acts as the predominant methanol dehydrogenase in the type I methanotroph Methylomicrobium buryatense. J. Bacteriol. 2016, 198, 1317–1325. [Google Scholar] [CrossRef] [Green Version]
- Vu, H.N.; Subuyuj, G.A.; Vijayakumar, S.; Good, N.M.; Martinez-Gomez, N.C.; Skovran, E. Lanthanide-dependent regulation of methanol oxidation systems in Methylobacterium extorquens AM1 and their contribution to methanol growth. J. Bacteriol. 2016, 198, 1250–1259. [Google Scholar] [CrossRef] [Green Version]
- Skovran, E.; Palmer, A.D.; Rountree, A.M.; Good, N.M.; Lidstrom, M.E. XoxF is required for expression of methanol dehydrogenase in Methylobacterium extorquens AM1. J. Bacteriol. 2011, 193, 6032–6038. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Huang, J.; Zhao, F.; Chistoserdova, L. Physiological effect of XoxG(4) on lanthanide-dependent methanotrophy. mBio 2018, 9, e02430-17. [Google Scholar] [CrossRef] [Green Version]
- Ochsner, A.M.; Hemmerle, L.; Vonderach, T.; Nüssli, R.; Bortfeld-Miller, M.; Hattendorf, B.; Vorholt, J.A. Use of rare-earth elements in the phyllosphere colonizer Methylobacterium extorquens PA1. Mol. Microbiol. 2019, 111, 1152–1166. [Google Scholar] [CrossRef] [Green Version]
- Roszczenko-Jasińska, P.; Vu, H.N.; Subuyuj, G.A.; Crisostomo, R.V.; Cai, J.; Raghuraman, C.; Ayala, E.M.; Clippard, E.J.; Lien, N.F.; Ngo, R.T.; et al. Lanthanide transport, storage, and beyond: Genes and processes contributing to XoxF function in Methylorubrum extorquens AM1. bioRxiv 2019. [Google Scholar] [CrossRef]
- Good, N.M.; Vu, H.N.; Suriano, C.J.; Subuyuj, G.A.; Skovran, E.; Martinez-Gomez, N.C. Pyrroloquinoline quinone ethanol dehydrogenase in Methylobacterium extorquens AM1 extends lanthanide-dependent metabolism to multicarbon substrates. J. Bacteriol. 2016, 198, 3109–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keltjens, J.T.; Pol, A.; Reimann, J.; Op den Camp, H.J.M. PQQ-dependent methanol dehydrogenases: Rare-earth elements make a difference. Appl. Microbiol. Biotechnol. 2014, 98, 6163–6183. [Google Scholar] [CrossRef] [PubMed]
- Chistoserdova, L. Modularity of methylotrophy, revisited. Environ. Microbiol. 2011, 13, 2603–2622. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.L.; Wessels, H.J.C.T.; Pol, A.; Op den Camp, H.J.M.; Jetten, M.S.M.; van Niftrik, L.; Keltjens, J.T. XoxF-type methanol dehydrogenase from the anaerobic methanotroph “Candidatus methylomirabilis oxyfera”. Appl. Environ. Microbiol. 2015, 81, 1442–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, A.; Akita, M.; Murase, H.; Kimbara, K. Culturable bacteria in hydroponic cultures of moss Racomitrium japonicum and their potential as biofertilizers for moss production. J. Biosci. Bioeng. 2011, 112, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Tani, A.; Takai, Y.; Suzukawa, I.; Akita, M.; Murase, H.; Kimbara, K. Practical application of methanol-mediated mutualistic symbiosis between Methylobacterium species and a roof greening moss, Racomitrium japonicum. PLoS ONE 2012, 7, e33800. [Google Scholar] [CrossRef] [Green Version]
- Masuda, S.; Suzuki, Y.; Fujitani, Y.; Mitsui, R.; Nakagawa, T.; Shintani, M.; Tani, A. Lanthanide-dependent regulation of methylotrophy in Methylobacterium aquaticum strain 22A. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Alamgir, K.M.; Masuda, S.; Fujitani, Y.; Fukuda, F.; Tani, A. Production of ergothioneine by Methylobacterium species. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.; Harris, S.R.; Otto, T.D.; Berriman, M.; Parkhill, J.; McQuillan, J.A. BamView: Visualizing and interpretation of next-generation sequencing read alignments. Brief. Bioinform. 2012, 14, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarczyk, A.; Vorholt, J.A.; Francez-Charlot, A. Cumate-inducible gene expression system for sphingomonads and other alphaproteobacteria. Appl. Environ. Microbiol. 2013, 79, 6795–6802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, C.J.; Lidstrom, M.E. Development of improved versatile broad-host-range vectors for use in methylotrophs and other gram-negative bacteria. Microbiology 2001, 147, 2065–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Uotila, L.; Koivusalo, M. [42] formaldehyde dehydrogenase. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1981; pp. 314–320. [Google Scholar] [CrossRef]
- Good, N.M.; Walser, O.N.; Moore, R.S.; Suriano, C.J.; Huff, A.F.; Martínez-Gómez, N.C. Investigation of lanthanide-dependent methylotrophy uncovers complementary roles for alcohol dehydrogenase enzymes. bioRxiv 2018. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Suganuma, S.; Hibino, A.; Mitsui, R.; Tani, A.; Matsumoto, T.; Ebihara, A.; Fitriyanto, N.A.; Pertiwiningrum, A.; Shimada, M.; et al. Lanthanide-dependent methanol dehydrogenase from the legume symbiotic nitrogen-fixing bacterium Bradyrhizobium diazoefficiens strain USDA110. Enzyme Microb. Technol. 2019, 130, 109371. [Google Scholar] [CrossRef]
- Goenrich, M.; Bartoschek, S.; Hagemeier, C.H.; Griesinger, C.; Vorholt, J.A. A glutathione-dependent formaldehyde-activating enzyme (Gfa) from Paracoccus denitrificans detected and purified via two-dimensional proton exchange NMR spectroscopy. J. Biol. Chem. 2002, 277, 3069–3072. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yu, Z.; Chistoserdova, L. Lanthanide-dependent methanol dehydrogenases of XoxF4 and XoxF5 clades are differentially distributed among methylotrophic bacteria and they reveal different biochemical properties. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Chistoserdova, L. C1 transfer enzymes and coenzymes linking methylotrophic bacteria and methanogenic archaea. Science 1998, 281, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.G.; Hristova, K.R.; Lidstrom, M.E.; Chistoserdova, L. Characterization of a novel methanol dehydrogenase in representatives of Burkholderiales: Implications for environmental detection of methylotrophy and evidence for convergent evolution. J. Bacteriol. 2008, 190, 3817–3823. [Google Scholar] [CrossRef] [Green Version]
- Wehrmann, M.; Berthelot, C.; Billard, P.; Klebensberger, J. The PedS2/PedR2 two-component system is crucial for the rare earth element switch in Pseudomonas putida KT2440. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravind, L.; Ponting, C.P. The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signalling proteins. FEMS Microbiol. Lett. 1999, 176, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; Hager, K.R.; Miller, S.I. HAMP domain rotation and tilting movements associated with signal transduction in the PhoQ sensor kinase. mBio 2015, 6. [Google Scholar] [CrossRef]
- Šmejkalová, H.; Erb, T.J.; Fuchs, G. Methanol assimilation in Methylobacterium extorquens AM1: Demonstration of all enzymes and their regulation. PLoS ONE 2010, 5, e13001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurimoto, H.; Kato, N.; Sakai, Y. Assimilation, dissimilation, and detoxification of formaldehyde, a central metabolic intermediate of methylotrophic metabolism. Chem. Rec. 2005, 5, 367–375. [Google Scholar] [CrossRef]
- Marx, C.J.; Miller, J.A.; Chistoserdova, L.; Lidstrom, M.E. Multiple formaldehyde oxidation/detoxification pathways in Burkholderia fungorum LB400. J. Bacteriol. 2004, 186, 2173–2178. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Gomez, N.C.; Good, N.M.; Lidstrom, M.E. Methenyl-dephosphotetrahydromethanopterin is a regulatory signal for acclimation to changes in substrate availability in Methylobacterium extorquens AM1. J. Bacteriol. 2015, 197, 2020–2026. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanpirat, P.; Nakatsuji, Y.; Hiraga, S.; Fujitani, Y.; Izumi, T.; Masuda, S.; Mitsui, R.; Nakagawa, T.; Tani, A. Lanthanide-Dependent Methanol and Formaldehyde Oxidation in Methylobacterium aquaticum Strain 22A. Microorganisms 2020, 8, 822. https://doi.org/10.3390/microorganisms8060822
Yanpirat P, Nakatsuji Y, Hiraga S, Fujitani Y, Izumi T, Masuda S, Mitsui R, Nakagawa T, Tani A. Lanthanide-Dependent Methanol and Formaldehyde Oxidation in Methylobacterium aquaticum Strain 22A. Microorganisms. 2020; 8(6):822. https://doi.org/10.3390/microorganisms8060822
Chicago/Turabian StyleYanpirat, Patcha, Yukari Nakatsuji, Shota Hiraga, Yoshiko Fujitani, Terumi Izumi, Sachiko Masuda, Ryoji Mitsui, Tomoyuki Nakagawa, and Akio Tani. 2020. "Lanthanide-Dependent Methanol and Formaldehyde Oxidation in Methylobacterium aquaticum Strain 22A" Microorganisms 8, no. 6: 822. https://doi.org/10.3390/microorganisms8060822