Association between Gut Microbiota and Infant’s Temperament in the First Year of Life in a Chinese Birth Cohort



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Setting, and Participants
2.2. Data Collection
2.3. Statistical Methods
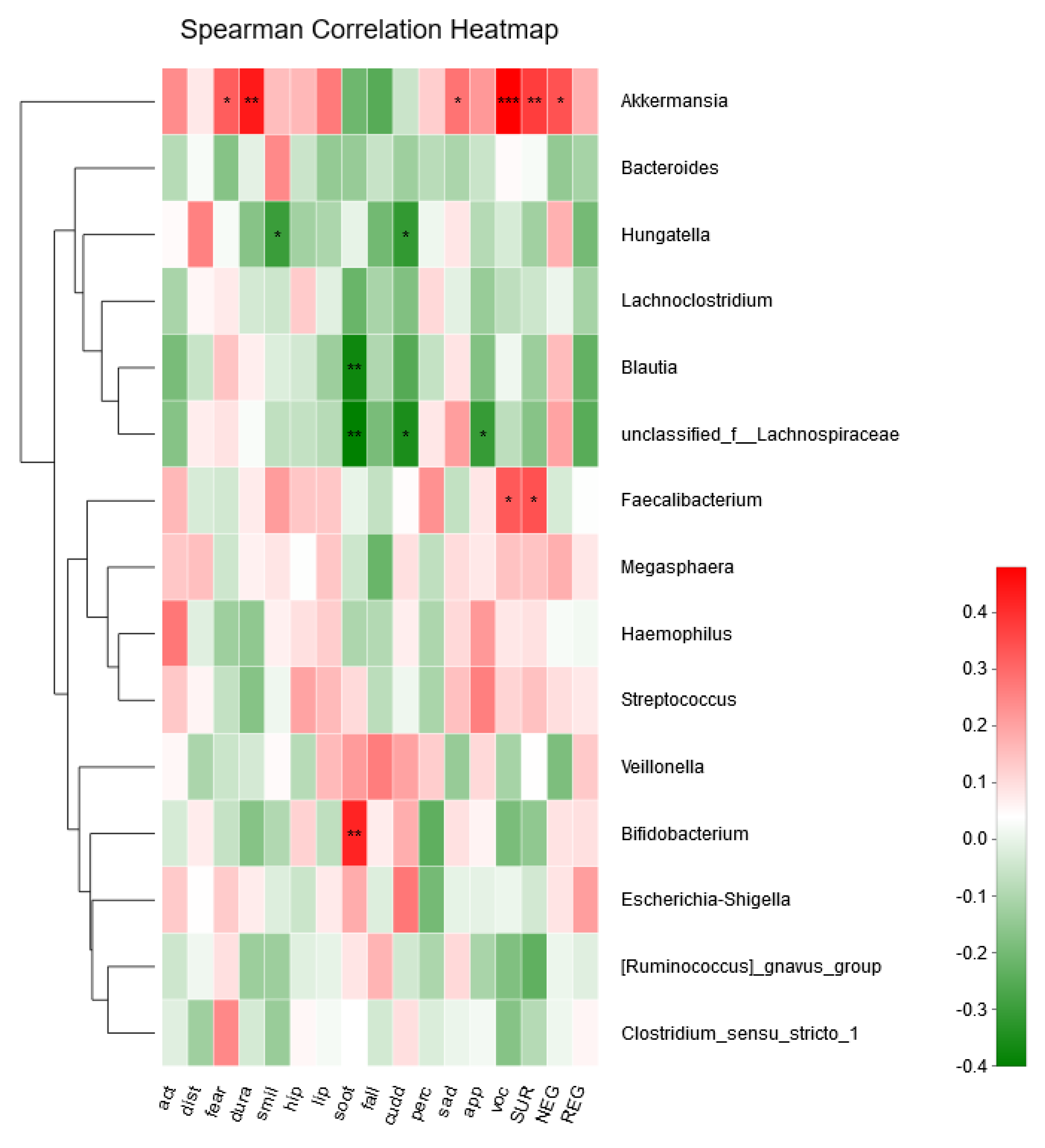
3. Results
3.1. Participant Characteristics
3.2. Infant Temperament at 12 Months Old
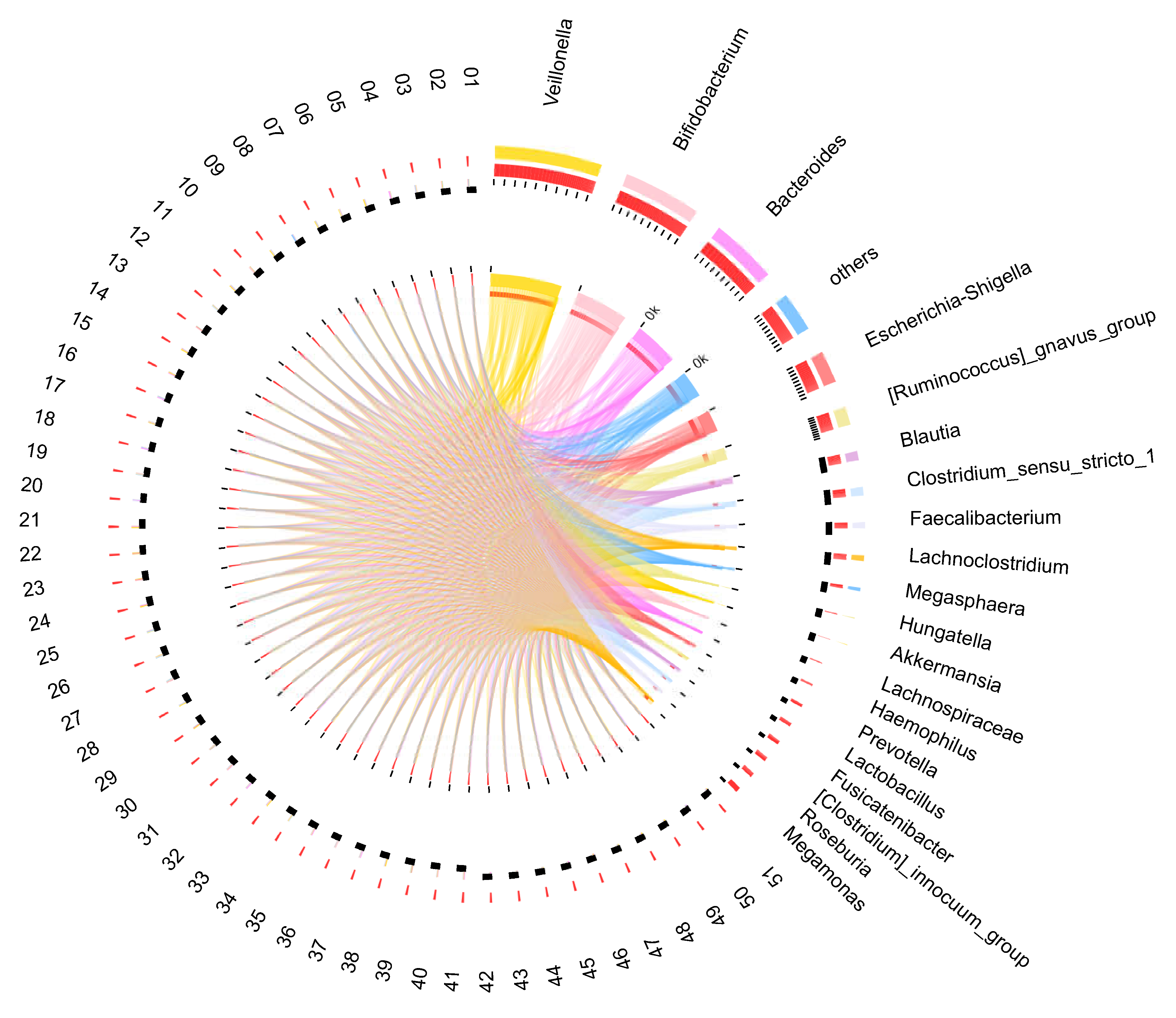
3.3. Infant Gut Microbiota at 12 Months Old
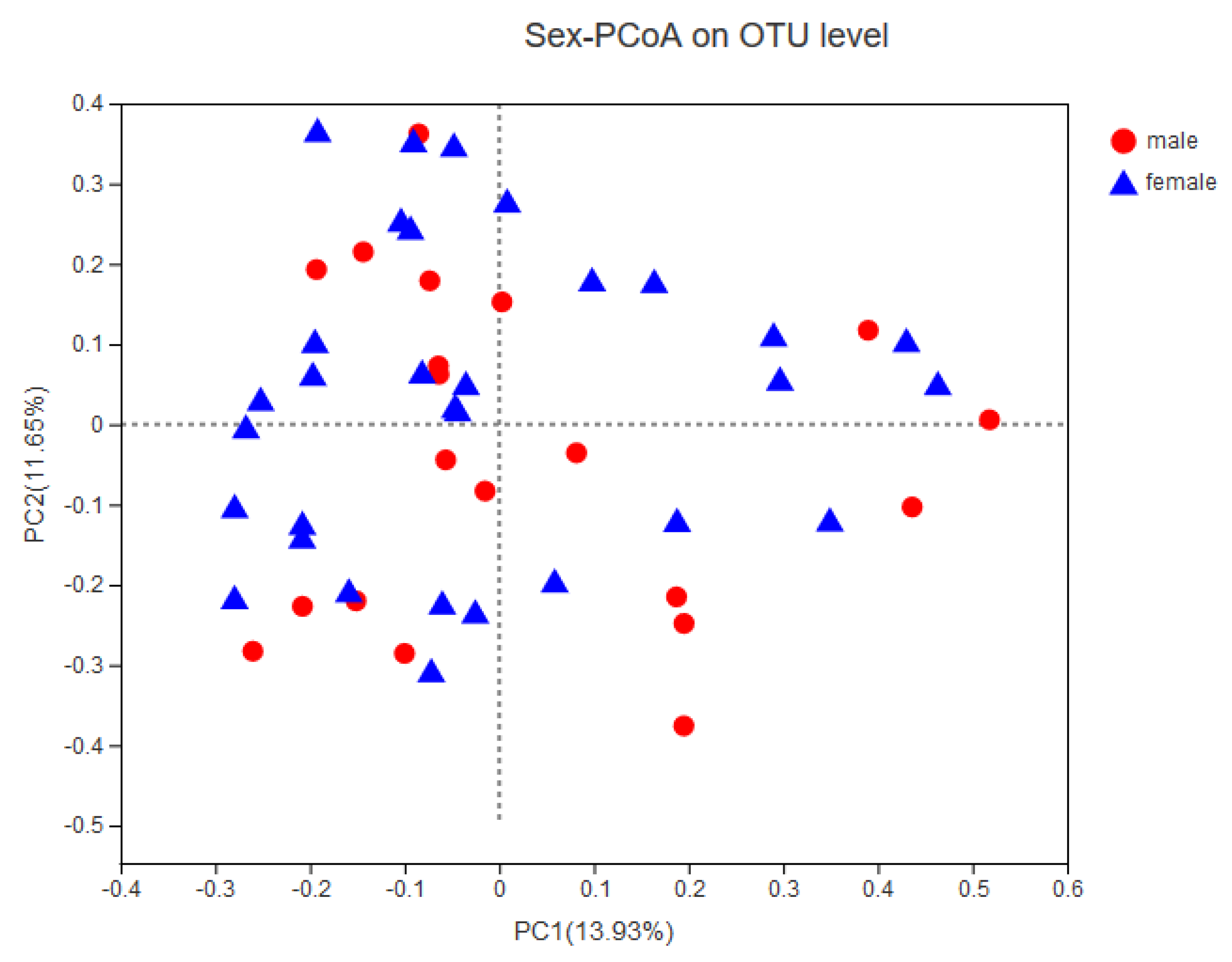
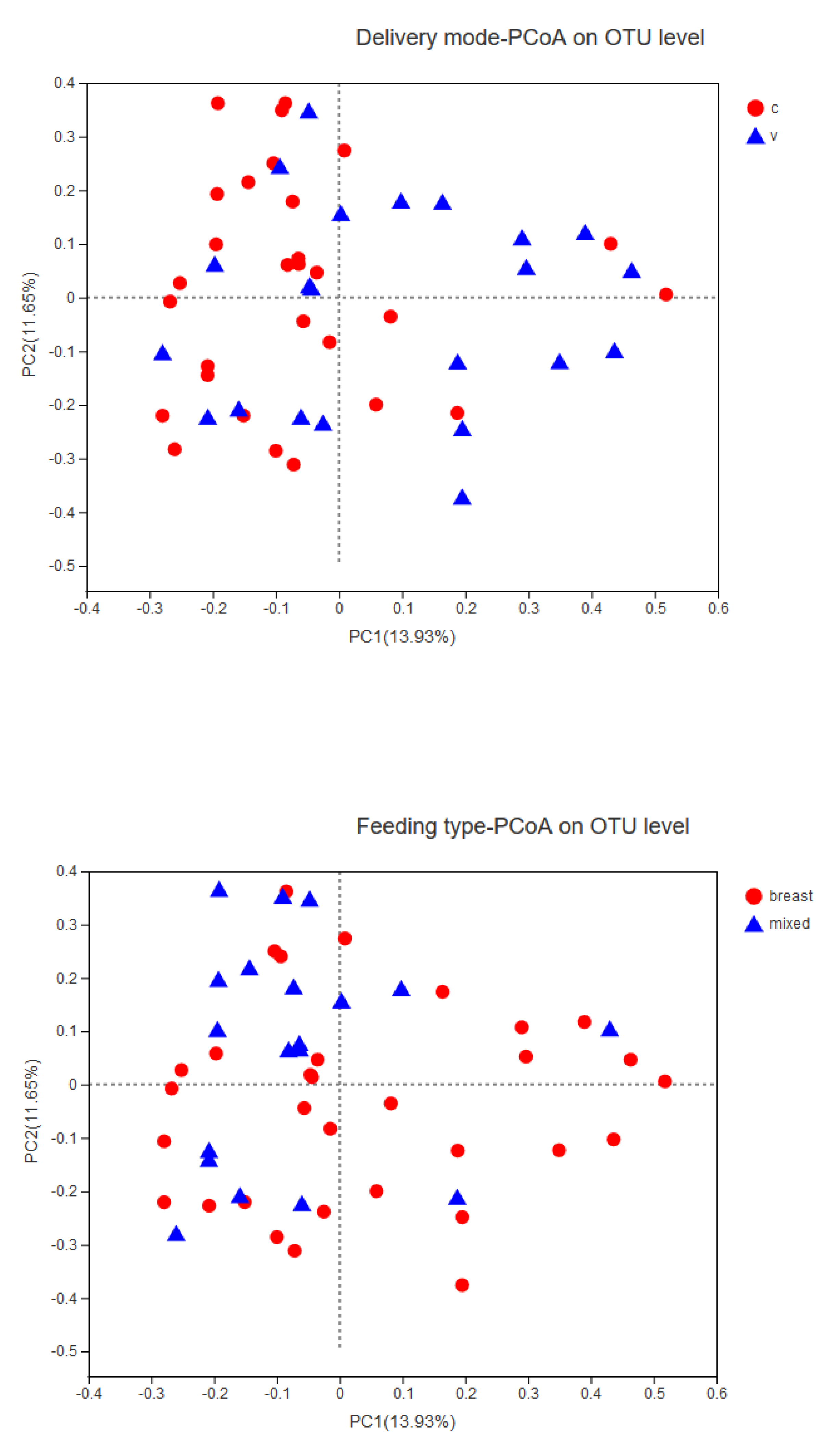
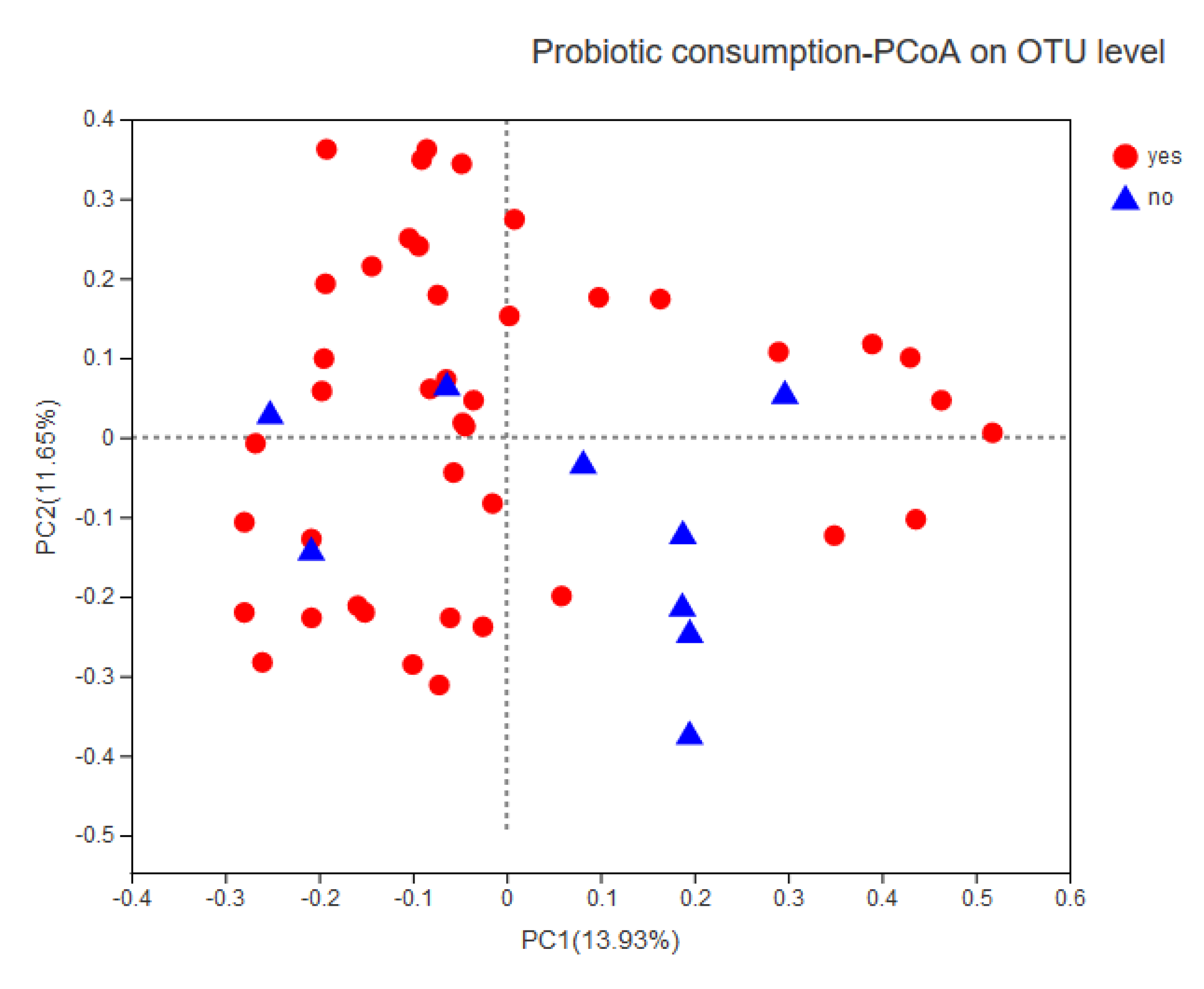
3.4. Association between Infant Temperament and the Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rothbart, M.K. Measurement of Temperament in Infancy. Child Dev. 1981, 52, 569–578. [Google Scholar] [CrossRef]
- Gartstein, M.A.; Rothbart, M.K. Studying infant temperament via the Revised Infant Behavior Questionnaire. Infant Behav. Dev. 2003, 26, 64–86. [Google Scholar] [CrossRef]
- Cozzi, P.; Putnam, S.P.; Menesini, E.; Gartstein, M.A.; Aureli, T.; Calussi, P.; Montirosso, R. Studying cross-cultural differences in temperament in toddlerhood: United States of America (US) and Italy. Infant Behav. Dev. 2013, 36, 480–483. [Google Scholar] [CrossRef]
- Enlow, M.B.; White, M.T.; Hails, K.; Cabrera, I.; Wright, R.J. The Infant Behavior Questionnaire-Revised: Factor structure in a culturally and sociodemographically diverse sample in the United States. Infant Behav. Dev. 2016, 43, 24–35. [Google Scholar] [CrossRef] [Green Version]
- de Lauzon-Guillain, B.; Wijndaele, K.; Clark, M.; Acerini, C.L.; Hughes, I.A.; Dunger, D.B.; Wells, J.C.; Ong, K.K. Breastfeeding and Infant Temperament at Age Three Months. PLoS ONE 2012, 7, e29326. [Google Scholar] [CrossRef] [Green Version]
- Gartstein, M.A.; Gonzalez, C.; Carranza, J.A.; Ahadi, S.A.; Ye, R.; Rothbart, M.K.; Yang, S.W. Studying Cross-cultural Differences in the Development of Infant Temperament: People’s Republic of China, the United States of America, and Spain. Child Psychiatry Hum. Dev. 2006, 37, 145–161. [Google Scholar] [CrossRef]
- Gartstein, M.A.; Putnick, D.; Kwak, K.; Hahn, C.S.; Bornstein, M.H. Stability of temperament in South Korean infants from 6 to 12 to 18 months: Moderation by age, gender, and birth order. Infant Behav. Dev. 2015, 40, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Turkheimer, E.; Waldron, M. Nonshared environment: A theoretical, methodological, and quantitative review. Psychol. Bull. 2000, 126, 78–108. [Google Scholar] [CrossRef]
- Keightley, P.; Pavli, P.; Platten, J.; Looi, J.C.L. Gut feelings 1. Mind, mood and gut in irritable bowel syndrome: Approaches to psychiatric care. Australas. Psychiatry 2015, 23, 403–406. [Google Scholar] [CrossRef]
- Keightley, P.; Pavli, P.; Platten, J.; Looi, J.C.L. Gut feelings 2. Mind, mood and gut in inflammatory bowel disease: Approaches to psychiatric care. Australas. Psychiatry 2015, 23, 407–410. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2016. [Google Scholar] [CrossRef]
- Foster, J.; Neufeld, K.A. Gut-brain axis: How the microbiome influences anxiety and depression. Int. J. Neuropsychopharmacol. 2014, 17, 27. [Google Scholar] [CrossRef]
- Steenbergen, L.; Sellaro, R.; van Hemert, S.; Bosch, J.A.; Colzato, L.S. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain Behav. Immun. 2015, 48, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, M.; Violle, N.; Bisson, J.F.; Desor, D.; Javelot, H.; Rougeot, C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2011, 2, 256–261. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [Green Version]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain-Raspaud, S.; Trotin, B.; Naliboff, B.; et al. Consumption of fermented milk product with probiotic modulates brain activity. Gastroenterology 2013, 144, 1394–1401. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Salzwedel, A.P.; Carlson, A.L.; Xia, K.; Azcarate-Peril, M.A.; Styner, M.A.; Thompson, A.L.; Geng, X.; Goldman, B.D.; Gilmore, J.H.; et al. Gut microbiome and brain functional connectivity in infants-a preliminary study focusing on the amygdala. Psychopharmacology 2019. [Google Scholar] [CrossRef]
- Kim, H.; Park, Y.J. The Association between Temperament and Microbiota in Healthy Individuals: A Pilot Study. Psychosom. Med. 2017, 79, 898. [Google Scholar] [CrossRef]
- Christian, L.M.; Galley, J.D.; Hade, E.M.; Schoppe-Sullivan, S.; Dush, C.K.; Bailey, M.T. Gut microbiome composition is associated with temperament during early childhood. Brain Behav. Immun. 2015, 45, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Putnam, S.P.; Rothbart, M.K.; Gartstein, M.A. Homotypic and heterotypic continuity of fine-grained temperament during infancy, toddlerhood, and early childhood. Infant Child Dev. 2008, 17, 387–405. [Google Scholar] [CrossRef]
- Li, Y.; Faden, H.S.; Zhu, L. The Response of the Gut Microbiota to Dietary Changes in the First Two Years of Life. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Y.; Bai, J.; Chen, X. The Effect of Maternal Postpartum Practices on Infant Gut Microbiota: A Chinese Cohort Study. Microorganisms 2019, 7, 511. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.W.; Tang, L.Q.; Zhao, H.; Stenvang, J.; Li, Y.L.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 12 January 2019).
- Lerner, R.M. Handbook of Child Psychology and Developmental Science; WILEY: Hoboken, NJ, USA, 2015. [Google Scholar]
- Coll, C.T.G.; Halpern, L.F.; Vohr, B.R.; Seifer, R.; Oh, W. Stability and Correlates of Change of Early Temperament in Preterm and Full-Term Infants. Infant Behav. Dev. 1992, 15, 137–153. [Google Scholar] [CrossRef]
- Livas-Dlott, A.; Fuller, B.; Stein, G.L.; Bridges, M.; Mangual Figueroa, A.; Mireles, L. Commands, competence, and carino: Maternal socialization practices in Mexican American families. Dev. Psychol. 2010, 46, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Friend, M.; DeAnda, S.; Arias-Trejo, N.; Poulin-Dubois, D.; Zesiger, P. Developmental changes in maternal education and minimal exposure effects on vocabulary in English- and Spanish-learning toddlers. J. Exp. Child Psychol. 2017, 164, 250–259. [Google Scholar] [CrossRef]
- Lai, J. The Effects of Socioeconomic Status and Parent-Infant Interactions on Early Lexical Development of 12-to 24-Month-Old Infants. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2013. [Google Scholar]
- Tamis-LeMonda, C.S.; Kuchirko, Y.; Song, L.L. Why Is Infant Language Learning Facilitated by Parental Responsiveness? Curr. Dir. Psychol. Sci. 2014, 23, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Meyrose, A.K.; Klasen, F.; Otto, C.; Gniewosz, G.; Lampert, T.; Ravens-Sieberer, U. Benefits of maternal education for mental health trajectories across childhood and adolescence. Soc. Sci. Med. 2018, 202, 170–178. [Google Scholar] [CrossRef]
- Thompson, A.L.; Adair, L.S.; Bentley, M.E. Maternal Characteristics and Perception of Temperament Associated With Infant TV Exposure. Pediatrics 2013, 131, E390–E397. [Google Scholar] [CrossRef] [Green Version]
- Deanda, S.; Arias-Trejo, N.; Poulin-Dubois, D.; Zesiger, P.; Friend, M. Minimal second language exposure, SES, and early word comprehension: New evidence from a direct assessment. Biling.-Lang. Cogn. 2016, 19, 162–180. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.; Greene, M.M.; Patel, A.L.; Meier, P. Maternal Education Level Predicts Cognitive, Language, and Motor Outcome in Preterm Infants in the Second Year of Life. Am. J. Perinatol. 2016, 33, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Putnam, S.P.; Gartstein, M.A.; Rothbart, M.K. Measurement of fine-grained aspects of toddler temperament: The early childhood behavior questionnaire. Infant Behav. Dev. 2006, 29, 386–401. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sanchez, B.; Margolles, A. Bifidobacteria and Their Molecular Communication with the Immune System. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Kamei, A.; Xiao, J.Z.; Iwatsuki, K.; Abe, K. Bifidobacterium breve B-3 exerts metabolic syndrome-suppressing effects in the liver of diet-induced obese mice: A DNA microarray analysis. Benef. Microbes 2013, 4, 247–251. [Google Scholar] [CrossRef]
- Wang, Z.H.; Gao, Q.Y.; Fang, J.Y. Meta-analysis of the efficacy and safety of Lactobacillus-containing and Bifidobacterium-containing probiotic compound preparation in Helicobacter pylori eradication therapy. J. Clin. Gastroenterol. 2013, 47, 25–32. [Google Scholar] [CrossRef]
- Kondo, J.; Xiao, J.Z.; Shirahata, A.; Baba, M.; Abe, A.; Ogawa, K.; Shimoda, T. Modulatory effects of Bifidobacterium longum BB536 on defecation in elderly patients receiving enteral feeding. World J. Gastroenterol. 2013, 19, 2162–2170. [Google Scholar] [CrossRef]
- Sampson, T.R.; Mazmanian, S.K. Control of Brain Development, Function, and Behavior by the Microbiome. Cell Host Microbe 2015, 17, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Savignac, H.M.; Kiely, B.; Dinan, T.G.; Cryan, J.F. Bifidobacteria exert strain-specific effects on stress-related behavior and physiology in BALB/c mice. Neurogastroenterol. Motil. 2014, 26, 1615–1627. [Google Scholar] [CrossRef]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Sugahara, H.; Shimada, K.; Mitsuyama, E.; Kuhara, T.; Yasuoka, A.; Kondo, T.; Abe, K.; Xiao, J.Z. Therapeutic potential of Bifidobacterium breve strain A1 for preventing cognitive impairment in Alzheimer’s disease. Sci. Rep. 2017, 7, 13510. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Kobayashi, Y.; Mori, N.; Sakagawa, M.; Xiao, J.Z.; Moritani, T.; Sakane, N.; Nagai, N. Effect of combined bifidobacteria supplementation and resistance training on cognitive function, body composition and bowel habits of healthy elderly subjects. Benef. Microbes 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.P.; Hutch, W.; Borre, Y.E.; Kennedy, P.J.; Temko, A.; Boylan, G.; Murphy, E.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Bifidobacterium longum 1714 as a translational psychobiotic: Modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 2016, 6, e939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohara, T. Identification of the microbial diversity after fecal microbiota transplantation therapy for chronic intractable constipation using 16s rRNA amplicon sequencing. PLoS ONE 2019, 14, e0214085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Yawar, M.; Kumar, P.A.; Suresh, K. Hungatella effluvii gen. nov., sp. nov., an obligately anaerobic bacterium isolated from an effluent treatment plant, and reclassification of Clostridium hathewayi as Hungatella hathewayi gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 710–718. [Google Scholar] [CrossRef]
- Huang, L.S.; Gao, R.Y.; Yan, X.B. Structure analysis of the gut microbiota in chronic functional constipation patients. Chin. J. Colorectal Dis. 2017, 6, 121–126. [Google Scholar]
- Zao, A.B. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Ph.D. Thesis, Nanjing Medical University, Nanjing, China, 2018. [Google Scholar]
- Wei, L. Diversity of Gut Flora in Navy Officers and Soldiers Involved in Long Voyage. Ph.D. Thesis, The Second Military Medical University, Shanghai, China, 2017. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | N (%)/Median (IQR) |
|---|---|
| Maternal education | |
| <12 a | 4 (7.8%) |
| 12 a | 31 (60.8%) |
| >12 a | 16 (31.4%) |
| Sex | |
| Male | 20 (39.2%) |
| Female | 31 (60.8%) |
| Mode of delivery | |
| Caesarean | 29 (56.9%) |
| Vaginal | 22 (43.1%) |
| Feeding mode | |
| Breast | 32 (62.7%) |
| Mixed | 19 (37.3%) |
| Probiotic consumption | |
| Yes | 42 (82.4%) |
| No | 9 (17.6%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, X.; Yu, Y.; Liu, Y.; Zhang, Q.; Bai, J. Association between Gut Microbiota and Infant’s Temperament in the First Year of Life in a Chinese Birth Cohort. Microorganisms 2020, 8, 753. https://doi.org/10.3390/microorganisms8050753
Wang Y, Chen X, Yu Y, Liu Y, Zhang Q, Bai J. Association between Gut Microbiota and Infant’s Temperament in the First Year of Life in a Chinese Birth Cohort. Microorganisms. 2020; 8(5):753. https://doi.org/10.3390/microorganisms8050753
Chicago/Turabian StyleWang, Ying, Xiaoli Chen, Yun Yu, Yanqun Liu, Qing Zhang, and Jinbing Bai. 2020. "Association between Gut Microbiota and Infant’s Temperament in the First Year of Life in a Chinese Birth Cohort" Microorganisms 8, no. 5: 753. https://doi.org/10.3390/microorganisms8050753