Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol with Rhodosporidiobolus fluvialis Using a Two-Stage Batch-Cultivation Strategy with Separate Optimization of Each Stage

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain and Inoculum Preparation
2.2. Substrates Used for Lipid Production
2.3. Lipid Production from STH and CG by Two-Stage Batch Cultivation
2.3.1. Optimization of Cell Mass Production from STH in the First Stage
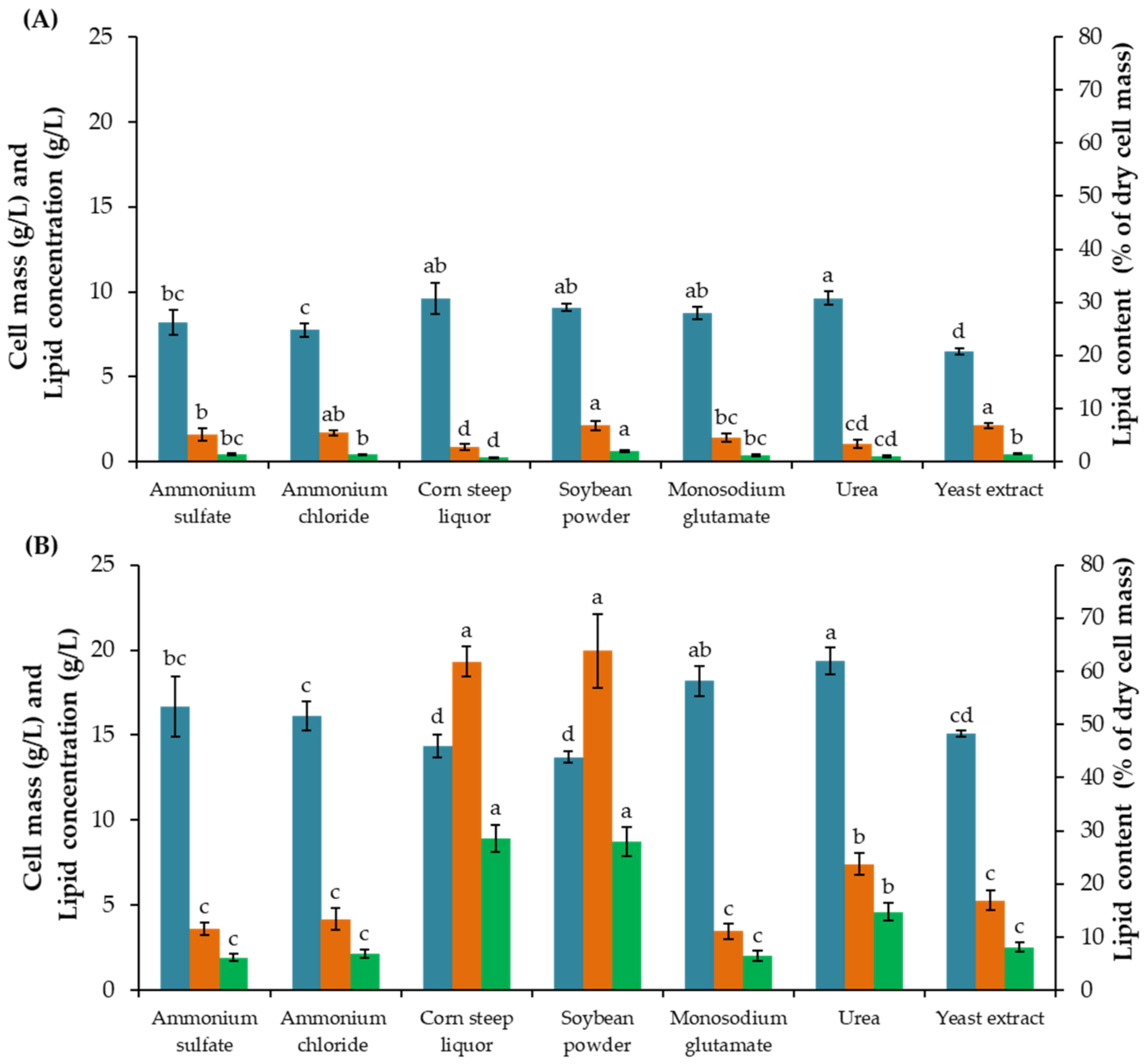
Effect of Additional Nitrogen Source
Screening of Significant Factors for Cell Mass
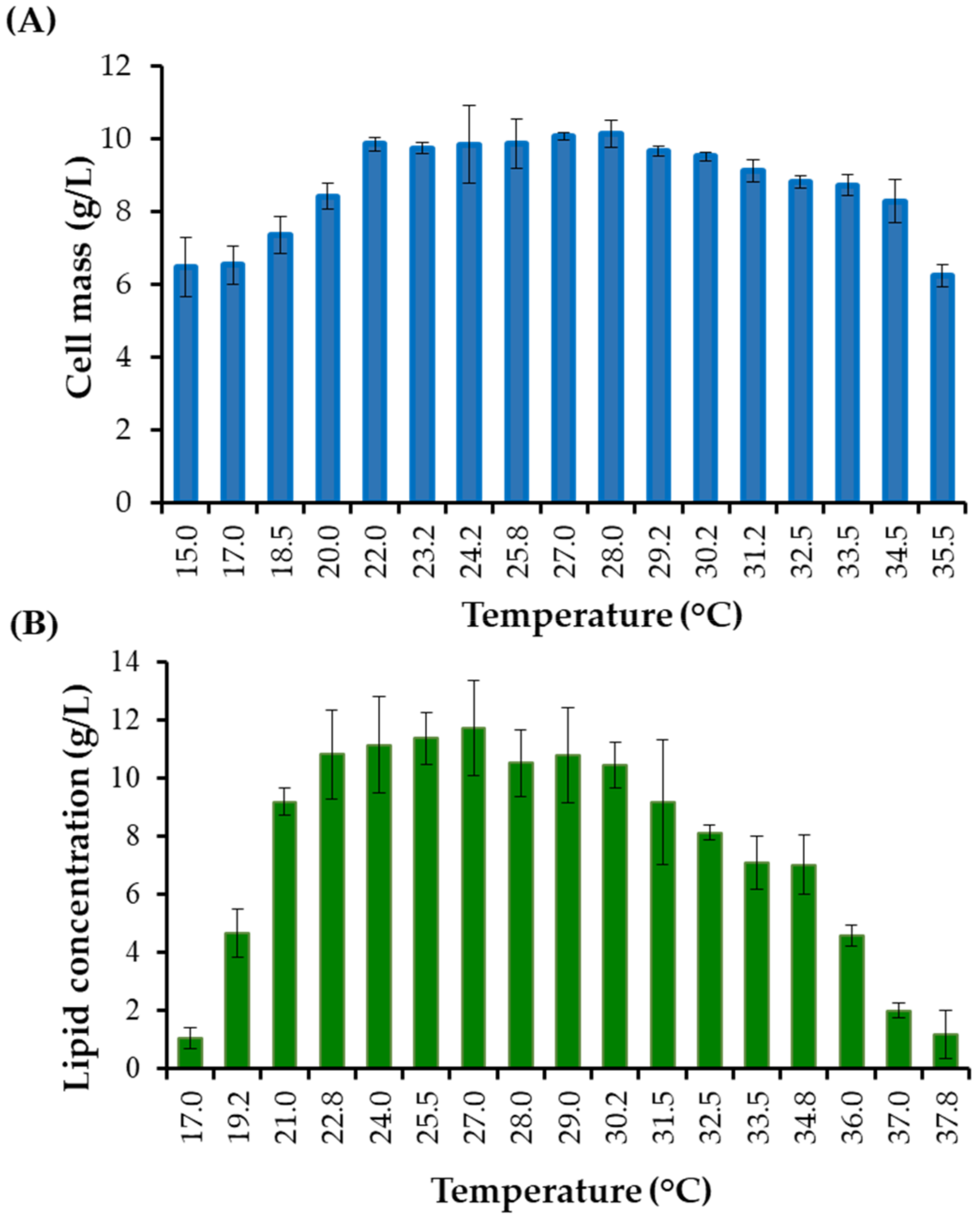
Optimization Temperature by the One-Factor-At-a-Time Methodology
Optimization of the Concentration of (NH4)2SO4 and KH2PO4, and Shaking Speed by Box-Behnken Design
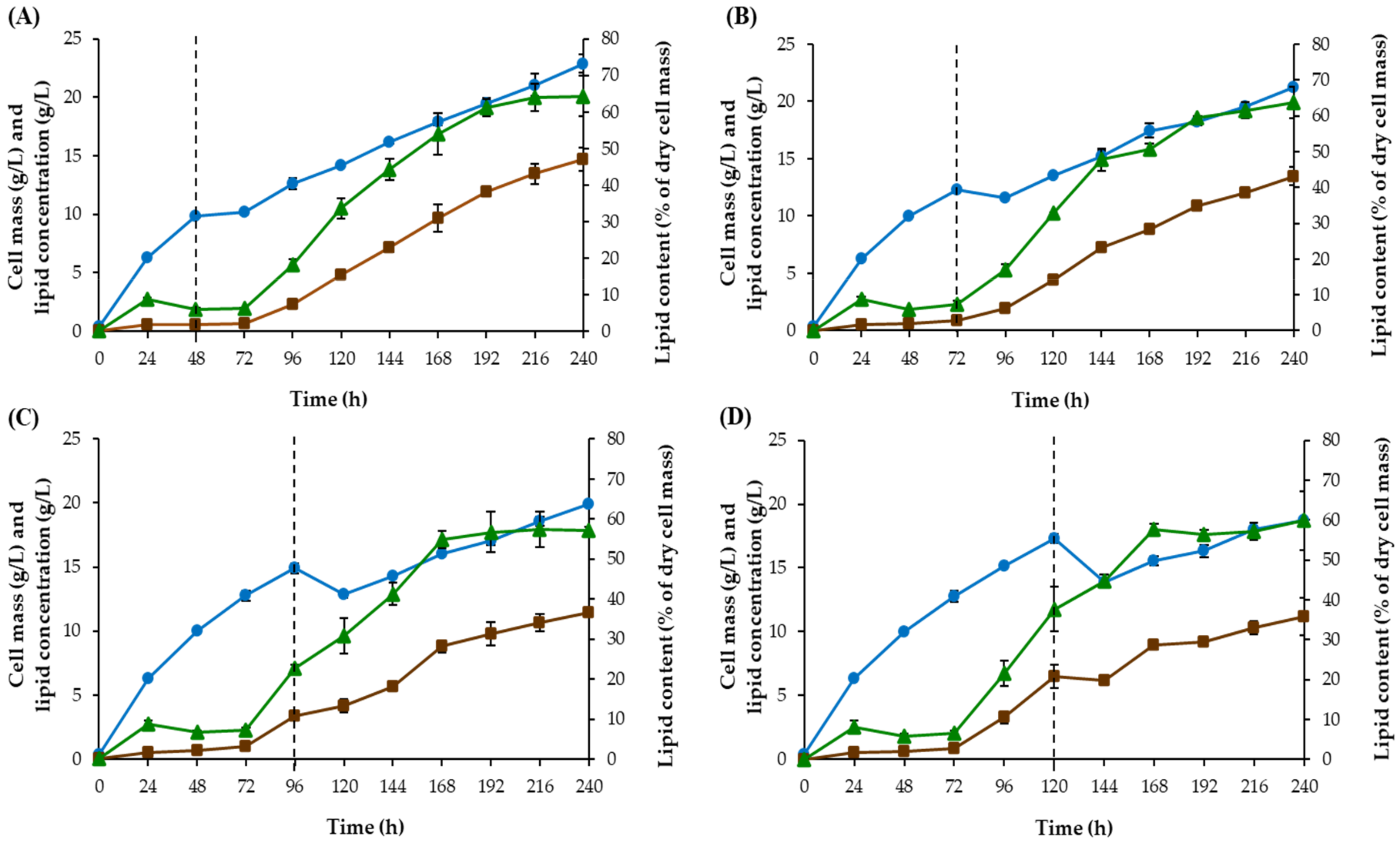
Shift Time Improvement
Initial TRS Concentration
2.3.2. Optimization of Lipid Production from CG in the Second Stage
2.4. Analytical Methods
3. Results and Discussion
3.1. Optimization of Cell Mass Production from STH in the First Stage
3.1.1. Effect of Additional Nitrogen Source
3.1.2. Screening of Significant Factors
3.1.3. Optimization of Temperature by the One-Factor-At-a-Time Method
3.1.4. Optimization of the Concentration of (NH4)2SO4, KH2PO4, and Shaking Speed by BBD
3.1.5. Shift Time Improvement
3.1.6. TRS Concentration
3.2. Optimization of Lipid Production from CG in the Second Stage by the One-Factor-At-a-Time Technique and BBD
3.3. Fatty Acid Compositions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Knothe, G.; Razon, L.F. Biodiesel fuels. Prog. Energy Combust. Sci. 2017, 58, 36–59. [Google Scholar] [CrossRef]
- Mohd Noor, C.W.; Noor, M.M.; Mamat, R. Biodiesel as alternative fuel for marine diesel engine applications: A review. Renew. Sustain. Energy Rev. 2018, 94, 127–142. [Google Scholar] [CrossRef]
- Knothe, G.; Krahl, J.; Van Gerpen, J. The Biodiesel Handbook, Second Edition; Elsevier Inc., Academic Press and AOCS Press: Cambridge, MA, USA, 2010; Current Status of the Biodiesel Industry, 494; pp. 299–399. [Google Scholar]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Valorization of Waste Frying Oils and Animal Fats for Biodiesel Production. In Advanced Biofuels and Bioproducts; Lee, J.W., Ed.; Springer: New York, NY, USA, 2013; pp. 671–693. [Google Scholar] [CrossRef]
- Parsons, S.; Raikova, S.; Chuck, C.J. The viability and desirability of replacing palm oil. Nat. Sustain. 2020. [Google Scholar] [CrossRef]
- Magdouli, S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Heterotrophic microorganisms: A promising source for biodiesel production. Crit. Rev. Environ. Sci. Technol. 2014, 44, 416–453. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Sestric, R.; Ignatia, L.; Levin, D.; German, J.B.; Gillies, L.A.; Almada, L.A.G.; Boundy-Mills, K.L. Manipulation of culture conditions alters lipid content and fatty acid profiles of a wide variety of known and new oleaginous yeast species. Bioresour. Technol. 2013, 144, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Amaretti, A.; Raimondi, S.; Sala, M.; Roncaglia, L.; De Lucia, M.; Leonardi, A.; Rossi, M. Single cell oils of the cold-adapted oleaginous yeast Rhodotorula glacialis DBVPG 4785. Microb. Cell Fact. 2010, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Chardot, T.; Nicaud, J.-M. Yarrowia lipolytica: A model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 2009, 91, 692–696. [Google Scholar] [CrossRef]
- Bellou, S.; Triantaphyllidou, I.E.; Mizerakis, P.; Aggelis, G. High lipid accumulation in Yarrowia lipolytica cultivated under double limitation of nitrogen and magnesium. J. Biotechnol. 2016, 234, 116–126. [Google Scholar] [CrossRef]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 2017, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-H.; Chang, K.-S.; Lee, C.-F.; Hsu, C.-L.; Huang, C.-W.; Jang, H.-D. Microbial lipid production by oleaginous yeast Cryptococcus sp. in the batch cultures using corncob hydrolysate as carbon source. Biomass Bioenergy 2015, 72, 95–103. [Google Scholar] [CrossRef]
- Xiong, L.; Huang, C.; Yang, X.-Y.; Lin, X.-Q.; Chen, X.-F.; Wang, C.; Wang, B.; Zeng, X.-A.; Chen, X.-D. Beneficial effect of corncob acid hydrolysate on the lipid production by oleaginous yeast Trichosporon dermatis. Prep. Biochem. Biotechnol. 2015, 45, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Matsakas, L.; Bonturi, N.; Miranda, E.A.; Rova, U.; Christakopoulos, P. High concentrations of dried sorghum stalks as a biomass feedstock for single cell oil production by Rhodosporidium toruloides. Biotechnol. Biofuels 2015, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.H.; de Andrade, C.C.P.; Ghiselli, G.; Maugeri, F. Exploration of Brazilian biodiversity and selection of a new oleaginous yeast strain cultivated in raw glycerol. Bioresour. Technol. 2013, 138, 377–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-H.; Chang, K.-S.; Hsu, C.-L.; Chuang, L.-T.; Chen, C.-Y.; Huang, F.-Y.; Jang, H.-D. A comparative study on batch and fed-batch cultures of oleaginous yeast Cryptococcus sp. in glucose-based media and corncob hydrolysate for microbial oil production. Fuel 2013, 105, 711–717. [Google Scholar] [CrossRef]
- Leiva-Candia, D.E.; Tsakona, S.; Kopsahelis, N.; García, I.L.; Papanikolaou, S.; Dorado, M.P.; Koutinas, A.A. Biorefining of by-product streams from sunflower-based biodiesel production plants for integrated synthesis of microbial oil and value-added co-products. Bioresour. Technol. 2015, 190, 57–65. [Google Scholar] [CrossRef]
- Poontawee, R.; Limtong, S. Feeding strategies of two-stage fed-batch cultivation processes for microbial lipid production from sugarcane top hydrolysate and crude glycerol by the oleaginous red yeast Rhodosporidiobolus fluvialis. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and carotenoid production by Rhodotorula glutinis under irradiation/high-temperature and dark/low-temperature cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Polburee, P.; Yongmanitchai, W.; Honda, K.; Ohashi, T.; Yoshida, T.; Fujiyama, K.; Limtong, S. Lipid production from biodiesel-derived crude glycerol by Rhodosporidium fluviale DMKU-RK253 using temperature shift with high cell density. Biochem. Eng. J. 2016, 112, 208–218. [Google Scholar] [CrossRef]
- Lin, J.; Shen, H.; Tan, H.; Zhao, X.; Wu, S.; Hu, C.; Zhao, Z.K. Lipid production by Lipomyces starkeyi cells in glucose solution without auxiliary nutrients. J. Biotechnol. 2011, 152, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Calvey, C.H.; Su, Y.-K.; Willis, L.B.; McGee, M.; Jeffries, T.W. Nitrogen limitation, oxygen limitation, and lipid accumulation in Lipomyces starkeyi. Bioresour. Technol. 2016, 200, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Liu, H.; Zhang, J.a. Enhanced lipid production with undetoxified corncob hydrolysate by Rhodotorula glutinis using a high cell density culture strategy. Bioresour. Technol. 2015, 180, 32–39. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, B.; Korstad, J. Utilization of lignocellulosic biomass by oleaginous yeast and bacteria for production of biodiesel and renewable diesel. Renew. Sustain. Energy Rev. 2017, 73, 654–671. [Google Scholar] [CrossRef]
- López, I.; Aranda, E.M.; Juárez, J.A.R.; Mendoza, G.D. Nutritional evaluation of eight sugarcane varieties with forage potential. Cuban J. Agric. Sci. 2003, 36, 375–380. [Google Scholar]
- Poontawee, R.; Yongmanitchai, W.; Limtong, S. Efficient oleaginous yeasts for lipid production from lignocellulosic sugars and effects of lignocellulose degradation compounds on growth and lipid production. Process Biochem. 2017, 53, 44–60. [Google Scholar] [CrossRef]
- Poontawee, R.; Yongmanitchai, W.; Limtong, S. Lipid production from a mixture of sugarcane top hydrolysate and biodiesel-derived crude glycerol by the oleaginous red yeast, Rhodosporidiobolus fluvialis. Process Biochem. 2018, 66, 150–161. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, Y.; Trushenski, J.; Blackburn, J.W. Converting crude glycerol derived from yellow grease to lipids through yeast fermentation. Bioresour. Technol. 2010, 101, 7581–7586. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Louhasakul, Y. Industrial wastes as a promising renewable source for production of microbial lipid and direct transesterification of the lipid into biodiesel. Bioresour. Technol. 2013, 142, 329–337. [Google Scholar] [CrossRef]
- Kitcha, S.; Cheirsilp, B. Enhancing lipid production from crude glycerol by newly isolated oleaginous yeasts: Strain selection, process optimization, and fed-batch strategy. Bioenergy Res. 2013, 6, 300–310. [Google Scholar] [CrossRef]
- Yang, X.; Jin, G.; Gong, Z.; Shen, H.; Bai, F.; Zhao, Z.K. Recycling biodiesel-derived glycerol by the oleaginous yeast Rhodosporidium toruloides Y4 through the two-stage lipid production process. Biochem. Eng. J. 2014, 91, 86–91. [Google Scholar] [CrossRef]
- Kraisintu, P.; Yongmanitchai, W.; Limtong, S. Selection and optimization for lipid production of a newly isolated oleaginous yeast, Rhodosporidium toruloides DMKU3-TK16. Kasetsart J. 2010, 44, 436–445. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Holub, B.J.; Skeaff, C.M. Nutritional regulation of cellular phosphatidylinositol. In Methods in Enzymology; Academic Press: Chambridge, MA, USA, 1987; Volume 141, pp. 234–244. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sushma, C.; Anand, A.P.; Veeranki, V.D. Enhanced production of glutaminase free L-asparaginase II by Bacillus subtilis WB800N through media optimization. Korean J. Chem. Eng. 2017, 34, 2901–2915. [Google Scholar] [CrossRef]
- Dai, C.-c.; Tao, J.; Xie, F.; Dai, Y.-j.; Zhao, M. Biodiesel generation from oleaginous yeast Rhodotorula glutinis with xylose assimilating capacity. Afr. J. Biotechnol. 2007, 6, 2130–2134. [Google Scholar]
- Huang, C.; Chen, X.-f.; Xiong, L.; Yang, X.-y.; Chen, X.-d.; Ma, L.-l.; Chen, Y. Microbial oil production from corncob acid hydrolysate by oleaginous yeast Trichosporon coremiiforme. Biomass Bioenergy 2013, 49, 273–278. [Google Scholar] [CrossRef]
- Kolouchová, I.; Maťátková, O.; Sigler, K.; Masák, J.; Řezanka, T. Lipid accumulation by oleaginous and non-oleaginous yeast strains in nitrogen and phosphate limitation. Folia Microbiol. 2016, 61, 431–438. [Google Scholar] [CrossRef]
- Bandhu, S.; Dasgupta, D.; Akhter, J.; Kanaujia, P.; Suman, S.K.; Agrawal, D.; Kaul, S.; Adhikari, D.K.; Ghosh, D. Statistical design and optimization of single cell oil production from sugarcane bagasse hydrolysate by an oleaginous yeast Rhodotorula sp. IIP-33 using response surface methodology. SpringerPlus 2014, 3, 691. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, Z.; Bai, F. High-density cultivation of oleaginous yeast Rhodosporidium toruloides Y4 in fed-batch culture. Enzyme Microb. Technol. 2007, 41, 312–317. [Google Scholar] [CrossRef]
- Trumbly, R.J. Glucose repression in the yeast Saccharomyces cerevisiae. Mol. Microbiol. 1992, 6, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Prabhu, A.A.; Gadela, R.; Bharali, B.; Deshavath, N.N.; Dasu, V.V. Development of high biomass and lipid yielding medium for newly isolated Rhodotorula mucilaginosa. Fuel 2019, 239, 874–885. [Google Scholar] [CrossRef]
- Fakas, S.; Makri, A.; Bellou, S.; Aggelis, G. Pathways to aerobic glycerol catabolism and their regulation. In Microbial Conversions of Raw Glycerol; Aggelis, G., Ed.; Nova Science Publishers Inc: New York, NY, USA, 2009; pp. 9–18. [Google Scholar]
- Capusoni, C.; Rodighiero, V.; Cucchetti, D.; Galafassi, S.; Bianchi, D.; Franzosi, G.; Compagno, C. Characterization of lipid accumulation and lipidome analysis in the oleaginous yeasts Rhodosporidium azoricum and Trichosporon oleaginosus. Bioresour. Technol. 2017, 238, 281–289. [Google Scholar] [CrossRef]
- Christophe, G.; Deo, J.L.; Kumar, V.; Nouaille, R.; Fontanille, P.; Larroche, C. Production of oils from acetic acid by the oleaginous yeast Cryptococcus curvatus. Appl. Biochem. Biotechnol. 2012, 167, 1270–1279. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential use of oleaginous red yeast Rhodotorula glutinis for the bioconversion of crude glycerol from biodiesel plant to lipids and carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Huang, X.-F.; Liu, J.-N.; Lu, L.-J.; Peng, K.-M.; Yang, G.-X.; Liu, J. Culture strategies for lipid production using acetic acid as sole carbon source by Rhodosporidium toruloides. Bioresour. Technol. 2016, 206, 141–149. [Google Scholar] [CrossRef]
- Fei, Q.; O’Brien, M.; Nelson, R.; Chen, X.; Lowell, A.; Dowe, N. Enhanced lipid production by Rhodosporidium toruloides using different fed-batch feeding strategies with lignocellulosic hydrolysate as the sole carbon source. Biotechnol. Biofuels 2016, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Awad, D.; Bohnen, F.; Mehlmer, N.; Brueck, T. Multi-factorial-guided media optimization for enhanced biomass and lipid formation by the oleaginous yeast Cutaneotrichosporon oleaginosus. Front. Bioeng. Biotechnol. 2019, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Giakoumis, E.G. Analysis of 22 vegetable oils’ physico-chemical properties and fatty acid composition on a statistical basis, and correlation with the degree of unsaturation. Renew. Energy 2018, 126, 403–419. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Cell Mass | Lipid Concentration | Lipid Content | |||
|---|---|---|---|---|---|---|
| T-Value | p-Value | T-Value | p-Value | T-Value | p-Value | |
| X1 | 1.8151 | 0.1671 | 2.0491 | 0.1329 | 1.8485 | 0.1617 |
| X2 | 1.3260 | 0.2767 | 2.2091 | 0.1142 | 2.3541 | 0.0999 |
| X3 | 2.7722 | 0.0694 | −1.0801 | 0.3592 | −4.3845 | 0.0220 |
| X4 | −2.6963 | 0.0740 | −2.6091 | 0.0797 | −2.6770 | 0.0752 |
| X5 | −0.1623 | 0.8814 | −0.9734 | 0.4021 | −1.4697 | 0.2380 |
| X6 | −0.9086 | 0.4305 | 0.3334 | 0.7608 | 0.6194 | 0.5795 |
| X7 | −2.6626 | 0.0762 | −1.4446 | 0.2443 | −0.6970 | 0.5359 |
| X8 | 4.3660 | 0.0222 | 4.1826 | 0.0249 | 3.9307 | 0.0293 |
| Source | SS | DF | MS | Cell Mass | |
|---|---|---|---|---|---|
| F-Value | p-Value | ||||
| Model | 116.85 | 9 | 12.98 | 69.42 | <0.0001 |
| X1 | 59.34 | 1 | 59.34 | 317.30 | <0.0001 |
| X2 | 0.17 | 1 | 0.17 | 0.93 | 0.3677 |
| X3 | 15.71 | 1 | 15.71 | 84.00 | <0.0001 |
| X1X2 | 0.00 | 1 | 0.00 | 0.01 | 0.9112 |
| X1X3 | 11.79 | 1 | 11.79 | 63.03 | <0.0001 |
| X2X3 | 0.01 | 1 | 0.01 | 0.05 | 0.8334 |
| X12 | 29.15 | 1 | 29.15 | 155.85 | <0.0001 |
| X22 | 0.03 | 1 | 0.03 | 0.13 | 0.7249 |
| X32 | 0.27 | 1 | 0.27 | 1.44 | 0.2684 |
| Lack of Fit | 0.90 | 3 | 0.30 | 2.92 | 0.1636 |
| R2 | 0.9889 | ||||
| Adjust R2 | 0.9747 | ||||
| TRS (g/L) | TRSC (% of Initial TRS) | Cell Mass (g/L) | Lipid Concentration (g/L) | Lipid Content (% of Dry Cell Mass) |
|---|---|---|---|---|
| 43.18 | 69.54 a | 10.61 ± 0.22 a | 0.65 ± 0.09 b | 5.66 ± 0.71 c |
| 52.21 | 54.22 b | 10.54 ± 0.16 a | 0.76 ± 0.08 a | 7.21 ± 0.82 b |
| 62.87 | 45.55 ab | 10.46 ± 0.13 a | 0.81 ± 0.08 a | 7.70 ± 0.69 ab |
| 72.26 | 39.05 b | 9.94 ± 0.22 b | 0.85 ± 0.05 a | 8.58 ± 0.65 a |
| 80.40 | 34.21 b | 9.93 ± 0.28 b | 0.87 ± 0.02 a | 8.70 ± 0.40 a |
| Source | DF | Cell Mass Production (Y1) | Lipid Concentration (Y2) | Lipid Content (Y3) | |||
|---|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | ||
| Model | 9 | 8.44 | 0.0151 | 14.66 | 0.0006 | 10.98 | 0.0084 |
| X1 | 1 | 27.08 | 0.0035 | 0.71 | 0.4240 | 13.14 | 0.0151 |
| X2 | 1 | 1.45 | 0.2817 | 0.15 | 0.7071 | 0.24 | 0.6480 |
| X3 | 1 | 0.60 | 0.474 | 0.76 | 0.4098 | 0.08 | 0.7876 |
| X1 X2 | 1 | 3.68 | 0.1133 | 0.22 | 0.6482 | 1.39 | 0.2920 |
| X1 X3 | 1 | 4.96 | 0.0765 | 17.23 | 0.0032 | 4.50 | 0.0873 |
| X2 X3 | 1 | 0.0057 | 0.9428 | 68.91 | <0.0001 | 52.49 | 0.0008 |
| X12 | 1 | 8.96 | 0.0303 | - | - | 1.28 | 0.3097 |
| X22 | 1 | 0.18 | 0.6907 | - | - | 1.34 | 0.2985 |
| X32 | 1 | 26.60 | 0.0036 | - | - | 23.94 | 0.0045 |
| Lack of Fit | 3 | 2.72 | 0.2806 | 0.91 | 0.6079 | 4.85 | 0.1758 |
| R2 | 0.9383 | 0.9167 | 0.9519 | ||||
| Adjust R2 | 0.8272 | 0.8541 | 0.8652 | ||||
| Stage (Cultivation Time) | TRSC (% of Initial TRS) | CGC (% of Initial CG) | Cell Mass (g/L) | Lipid Concentration (g/L) | Yx (g/g) | YL (g/g) |
|---|---|---|---|---|---|---|
| 1st stage (48 h) | 69.54 | - | 10.61 ± 0.22 | 0.65 ± 0.09 | 0.35 | 0.02 |
| 2nd stage (192 h) | - | 76.43 | 11.09 ± 0.23 | 15.20 ± 0.54 | 0.17 | 0.23 |
| Yeast | Carbon Sources | Cell Mass (g/L) | Lipid Concentration (g/L) | Lipid Content (% of Dry Cell Mass) | Lipid Productivity (g/L/h) | Culture Mode (Volume) | References |
|---|---|---|---|---|---|---|---|
| Rhodosporidiobolus fluvialis DMKU-SP314 | sugarcane top hydrolysate and crude glycerol | 21.7 | 15.8 | 73.0 | 0.066 | Two-stage batch flasks (100 mL) | This study |
| Rhodosporidiobolus fluvialis DMKU-SP314 | sugarcane top hydrolysate and crude glycerol | 38.5 | 23.6 | 61.9 | 0.098 | Two-stage fed-batch bioreactor (3 L) | [21] |
| Cryptococcus curvatus ATCC 20509 | crude glycerol | 32.9 | 17.4 | 53.0 | 0.060 | Two-stage fed-batch bioreactor (1.5 L) | [31] |
| Cryptococcus curvatus MUCL 29819 | glucose and acetate | 4.0 | 1.0 | 47.3 | 0.011 | Two-stage batch flasks (250 mL) | [50] |
| Rhodosporidiobolus fluvialis DMKU-RK253 | crude glycerol | 22.4 | 16.0 | 70.0 | 0.074 | Two-stage batch flasks (100 mL) | [23] |
| Rhodotorula glutinis TISTR 5159 | palm oil mill effluent | 8.8 | 4.6 | 51.9 | 0.064 | Two-stage semi-continuous bioreactor (1 L) | [51] |
| Rhodosporidium toruloides AS 2.1389 | glucose and acetate | 6.8 | 2.1 | 50.1 | 0.011 | Two-stage batch flasks (50 mL) | [52] |
| Rhodosporidium toruloides Y4 | glucose and glycerol | 20.3 | 8.6 | 42.5 | 0.051 | Two-stage batch flasks (50 mL) | [34] |
| Trichosporonoides spathulata JU4–57 | crude glycerol | 13.8 | 7.8 | 56.4 | 0.065 | Two-stage fed-batch bioreactor (2 L) | [33] |
| Yarrowia lipolytica TISTR 5151 | serum latex and glycerol | 7–8 | 3.4 | 44.5 | 0.024 | Two-stage batch flasks (50 mL) | [32] |
| Source | C14:0 | C16:0 | C18:0 | C18:1 | C18:2 | C18:3 |
|---|---|---|---|---|---|---|
| Rhodosporidiobolus fluvialis DMKU-SP314 | 1.7 | 31.0 | 5.4 | 36.7 | 20.7 | 4.4 |
| Canola | - | 4.5 | 2.0 | 60.4 | 21.2 | 9.4 |
| Castor | - | 1.4 | 1.1 | 3.4 | 4.8 | 0.6 |
| Olive | 0.1 | 11.3 | 2.8 | 74.5 | 9.8 | 0.5 |
| Palm | 1.1 | 42.3 | 4.3 | 40.9 | 10.1 | 0.3 |
| Peanut | - | 10.3 | 2.8 | 49.6 | 31.5 | 0.6 |
| Rapeseed | - | 4.1 | 1.5 | 62.3 | 20.6 | 8.7 |
| Soybean | 0.1 | 11.5 | 4.1 | 23.5 | 53.3 | 6.8 |
| Sunflower | - | 6.4 | 3.9 | 20.9 | 67.6 | 0.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonyarit, J.; Polburee, P.; Khaenda, B.; Zhao, Z.K.; Limtong, S. Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol with Rhodosporidiobolus fluvialis Using a Two-Stage Batch-Cultivation Strategy with Separate Optimization of Each Stage. Microorganisms 2020, 8, 453. https://doi.org/10.3390/microorganisms8030453
Boonyarit J, Polburee P, Khaenda B, Zhao ZK, Limtong S. Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol with Rhodosporidiobolus fluvialis Using a Two-Stage Batch-Cultivation Strategy with Separate Optimization of Each Stage. Microorganisms. 2020; 8(3):453. https://doi.org/10.3390/microorganisms8030453
Chicago/Turabian StyleBoonyarit, Jeerapan, Pirapan Polburee, Bongkot Khaenda, Zongbao K. Zhao, and Savitree Limtong. 2020. "Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol with Rhodosporidiobolus fluvialis Using a Two-Stage Batch-Cultivation Strategy with Separate Optimization of Each Stage" Microorganisms 8, no. 3: 453. https://doi.org/10.3390/microorganisms8030453