Bridging the Gap: A Role for Campylobacter jejuni Biofilms


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Motility and Chemotaxis
3. Glycobiology
4. Metabolism and Environmental Factors
5. Quorum Sensing
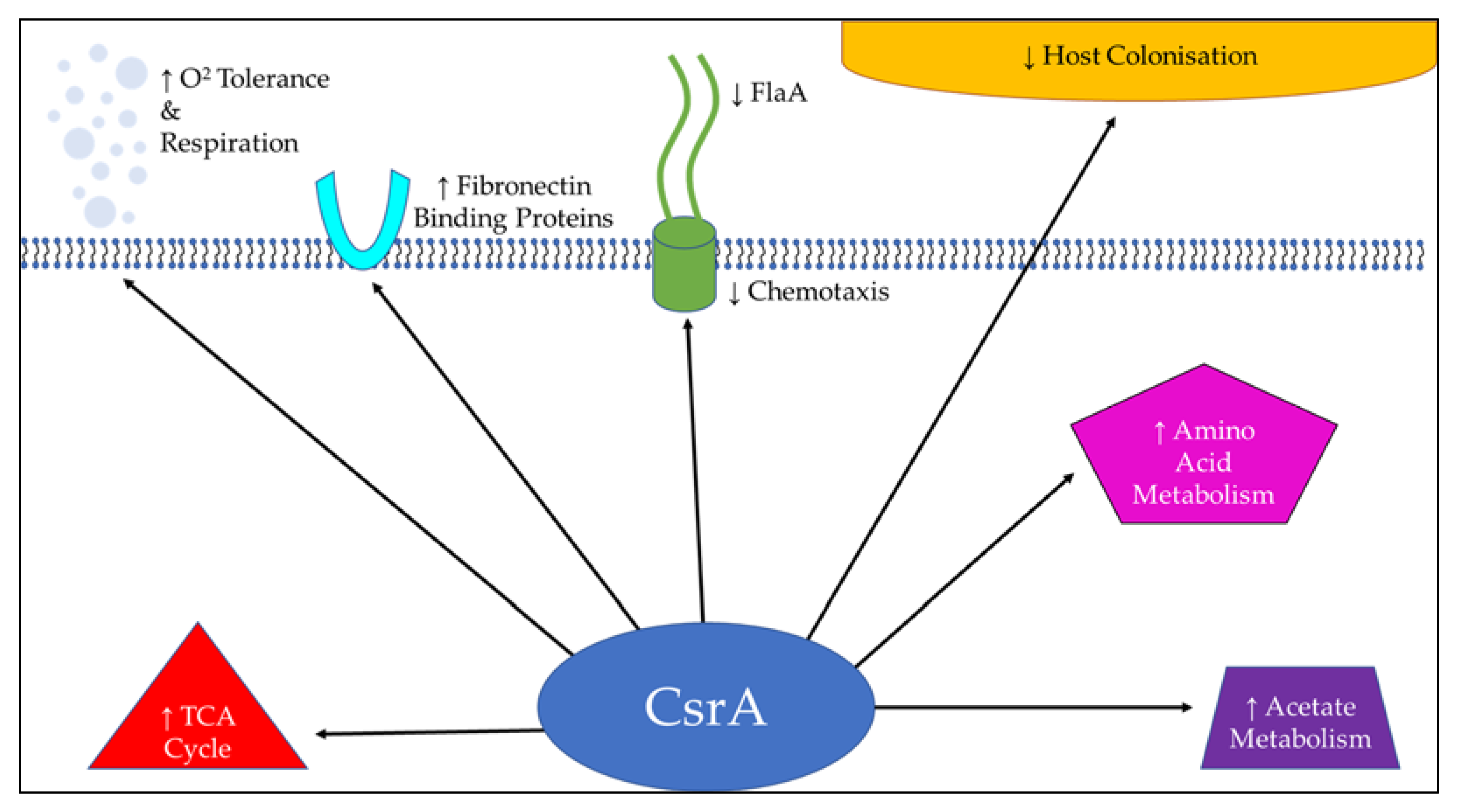
6. Stress Response Regulators
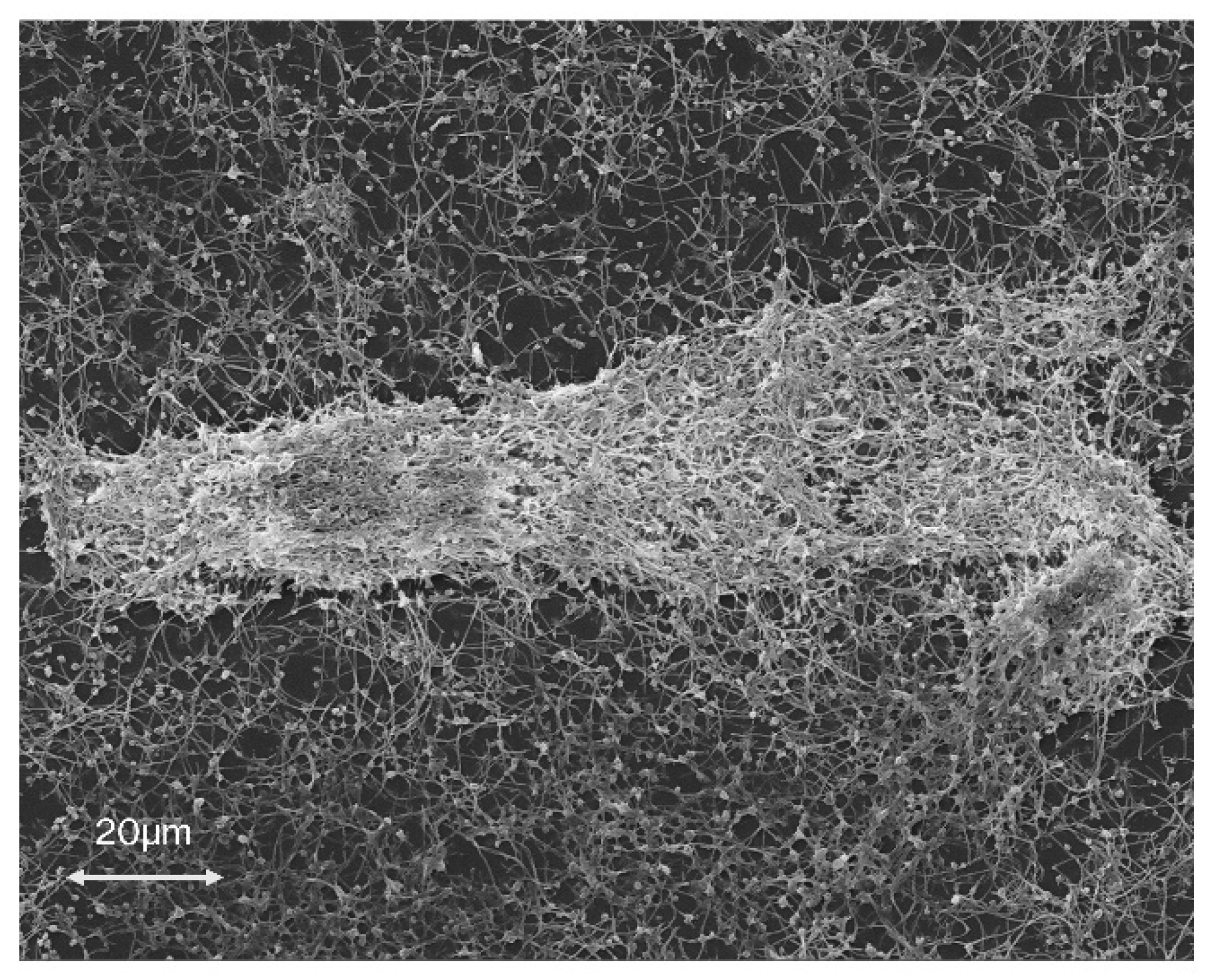
7. The Role of C. jejuni Biofilms
Author Contributions
Funding
Conflicts of Interest
References
- Lopez, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar]
- Hermans, D.; Pasmans, F.; Messens, W.; Martel, A.; Van Immerseel, F.; Rasschaert, G.; Heyndrickx, M.; Van Deun, K.; Haesebrouck, F. Poultry as a host for the zoonotic pathogen Campylobacter jejuni. Vector Borne Zoonotic Dis. 2012, 12, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moe, K.K.; Mimura, J.; Ohnishi, T.; Wake, T.; Yamazaki, W.; Nakai, M.; Misawa, N. The mode of biofilm formation on smooth surfaces by Campylobacter jejuni. Vet. Med. Sci. 2010, 72, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.T.; Fegan, N.; Turner, M.S.; Dykes, G.A. Role of attachment to surfaces on the prevalence and survival of Campylobacter through food systems. J. Food Prot. 2012, 75, 195–206. [Google Scholar] [CrossRef]
- Bae, J.; Jeon, B. Increased emergence of fluoroquinolone-resistant Campylobacter jejuni in biofilm. Antimicrob. Agents Chemother. 2013, 57, 5195–5196. [Google Scholar] [CrossRef] [Green Version]
- Melo, R.T.; Mendonça, E.P.; Monteiro, G.P.; Siqueira, M.C.; Pereira, C.B.; Peres, P.A.B.M.; Fernandez, H.; Rossi, D.A. Intrinsic and Extrinsic Aspects on Campylobacter jejuni Biofilms. Front. Microbiol. 2017, 8, 1332. [Google Scholar] [CrossRef] [Green Version]
- Turonova, H.; Neu, T.R.; Ulbrich, P.; Pazlarova, J.; Tresse, O. The biofilm matrix of Campylobacter jejuni determined by fluorescence lectin-binding analysis. Biofouling 2016, 32, 597–608. [Google Scholar] [CrossRef]
- McLennan, M.K.; Ringoir, D.D.; Frirdich, E.; Svensson, S.L.; Wells, D.H.; Jarrell, H.; Szymanski, C.M.; Gaynor, E.C. Campylobacter jejuni biofilms up-regulated in the absence of the stringent response utilize a calcofluor white-reactive polysaccharide. J. Bacteriol. 2008, 190, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.L.; Hanman, K.; Reuter, M.; Betts, R.P.; van Vliet, A.H. Campylobacter jejuni biofilms contain extracellular DNA and are sensitive to DNase I treatment. Front. Microbiol. 2015, 6, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, H.L.; Reuter, M.; Hanman, K.; Betts, R.P.; van Vliet, A.H. Prevention of biofilm formation and removal of existing biofilms by extracellular DNases of Campylobacter jejuni. PLoS ONE 2015, 10, e0121680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, G.H.; Lim, E.S.; Woo, M.A.; Lee, J.Y.; Kim, J.S.; Paik, H.D. Inverse Correlation between Extracellular DNase Activity and Biofilm Formation among Chicken-Derived Campylobacter Strains. J. Microbiol. Biotechnol. 2017, 27, 1942–1951. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Heredia, N.; Garcia, S. 2(5H)-Furanone, epigallocatechin gallate, and a citric-based disinfectant disturb quorum-sensing activity and reduce motility and biofilm formation of Campylobacter jejuni. Folia Microbiol. 2015, 60, 89–95. [Google Scholar] [CrossRef]
- Salamaszynska-Guz, A.; Rose, S.; Lykkebo, C.A.; Taciak, B.; Bacal, P.; Uspienski, T.; Douthwaite, S. Biofilm Formation and Motility Are Promoted by Cj0588-Directed Methylation of rRNA in Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2017, 7, 533. [Google Scholar] [CrossRef]
- Dwivedi, R.; Nothaft, H.; Garber, J.; Xin Kin, L.; Stahl, M.; Flint, A.; van Vliet, A.H.; Stintzi, A.; Szymanski, C.M. L-fucose influences chemotaxis and biofilm formation in Campylobacter jejuni. Mol. Microbiol. 2016, 101, 575–589. [Google Scholar] [CrossRef] [Green Version]
- Hartley-Tassell, L.E.; Shewell, L.K.; Day, C.J.; Wilson, J.C.; Sandhu, R.; Ketley, J.M.; Korolik, V. Identification and characterization of the aspartate chemosensory receptor of Campylobacter jejuni. Mol. Microbiol. 2010, 75, 710–730. [Google Scholar] [CrossRef]
- Rahman, H.; King, R.M.; Shewell, L.K.; Semchenko, E.A.; Hartley-Tassell, L.E.; Wilson, J.C.; Day, C.J.; Korolik, V. Characterisation of a multi-ligand binding chemoreceptor CcmL (Tlp3) of Campylobacter jejuni. PLoS Pathog. 2014, 10, e1003822. [Google Scholar] [CrossRef] [Green Version]
- Day, C.J.; King, R.M.; Shewell, L.K.; Tram, G.; Najnin, T.; Hartley-Tassell, L.E.; Wilson, J.C.; Fleetwood, A.D.; Zhulin, I.B.; Korolik, V. A direct-sensing galactose chemoreceptor recently evolved in invasive strains of Campylobacter jejuni. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Lux, R.; Shi, W. Chemotaxis-guided Movements in Bacteria. Crit. Rev. Oral Biol. Med. 2004, 15, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, M.K.; Paul, K.; Blair, D. Chemotaxis signaling protein CheY binds to the rotor protein FliN to control the direction of flagellar rotation in Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 9370–9375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Park, C.; Kim, Y.J. Role of flgA for Flagellar Biosynthesis and Biofilm Formation of Campylobacter jejuni NCTC11168. J. Microbiol. Biotechnol. 2015, 25, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Mallet, A.; Pearson, B.M.; van Vliet, A.H. Biofilm formation by Campylobacter jejuni is increased under aerobic conditions. Appl. Environ. Microbiol. 2010, 76, 2122–2128. [Google Scholar] [CrossRef] [Green Version]
- Kalmokoff, M.; Lanthier, P.; Tremblay, T.L.; Foss, M.; Lau, P.C.; Sanders, G.; Austin, J.; Kelly, J.; Szymanski, C. Proteomic analysis of Campylobacter jejuni 11168 biofilms reveals a role for the motility complex in biofilm formation. J. Bacteriol. 2006, 188, 4312–4320. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekhar, K.; Gangaiah, D.; Pina-Mimbela, R.; Kassem, I.I.; Jeon, B.H.; Rajashekara, G. Transducer like proteins of Campylobacter jejuni 81–176: Role in chemotaxis and colonization of the chicken gastrointestinal tract. Front. Cell. Infect. Microbiol. 2015, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Tram, G.; Klare, W.P.; Cain, J.A.; Mourad, B.; Cordwell, S.J.; Korolik, V.; Day, C.J. Assigning a role for chemosensory signal transduction in Campylobacter jejuni biofilms using a combined omics approach. Sci. Rep. Unpublished.
- Misawa, N.; Blaser, M.J. Detection and characterization of autoagglutination activity by Campylobacter jejuni. Infect. Immun. 2000, 68, 6168–6175. [Google Scholar] [CrossRef]
- Howard, S.L.; Jagannathan, A.; Soo, E.C.; Hui, J.P.; Aubry, A.J.; Ahmed, I.; Karlyshev, A.; Kelly, J.F.; Jones, M.A.; Stevens, M.P.; et al. Campylobacter jejuni glycosylation island important in cell charge, legionaminic acid biosynthesis, and colonization of chickens. Infect. Immun. 2009, 77, 2544–2556. [Google Scholar] [CrossRef] [Green Version]
- Teren, M.; Turonova Michova, H.; Vondrakova, L.; Demnerova, K. Molecules Autoinducer 2 and cjA and Their Impact on Gene Expression in Campylobacter jejuni. J. Mol. Microbiol. Biotechnol. 2018, 28, 207–215. [Google Scholar] [CrossRef]
- El Abbar, F.M.; Li, J.; Owen, H.C.; Daugherty, C.L.; Fulmer, C.A.; Bogacz, M.; Thompson, S.A. RNA Binding by the Campylobacter jejuni Post-transcriptional Regulator CsrA. Front. Microbiol. 2019, 10, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simunovic, K.; Ramic, D.; Xu, C.; Smole Mozina, S. Modulation of Campylobacter jejuni Motility, Adhesion to Polystyrene Surfaces, and Invasion of INT407 Cells by Quorum-Sensing Inhibition. Microorganisms 2020, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Karlyshev, A.V.; Ketley, J.M.; Wren, B.W. The Campylobacter jejuni glycome. FEMS Microbiol. Rev. 2005, 29, 377–390. [Google Scholar]
- Semchenko, E.A.; Day, C.J.; Wilson, J.C.; Grice, I.D.; Moran, A.P.; Korolik, V. Temperature-dependent phenotypic variation of Campylobacter jejuni lipooligosaccharides. BMC Microbiol. 2010, 10, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, C.J.; Semchenko, E.A.; Korolik, V. Glycoconjugates play a key role in Campylobacter jejuni Infection: Interactions between host and pathogen. Front. Cell. Infect. Microbiol. 2012, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frirdich, E.; Whitfield, C. Lipopolysaccharide inner core oligosaccharide structure and outer membrane stability in human pathogens belonging to the Enterobacteriaceae. J. Endotoxin Res. 2005, 11, 133–144. [Google Scholar] [PubMed]
- Naito, M.; Frirdich, E.; Fields, J.A.; Pryjma, M.; Li, J.; Cameron, A.; Gilbert, M.; Thompson, S.A.; Gaynor, E.C. Effects of sequential Campylobacter jejuni 81–176 lipooligosaccharide core truncations on biofilm formation, stress survival, and pathogenesis. J. Bacteriol. 2010, 192, 2182–2192. [Google Scholar] [CrossRef] [Green Version]
- Wacker, M.; Linton, D.; Hitchen, P.G.; Nita-Lazar, M.; Haslam, S.M.; North, S.J.; Aebi, M. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 2002, 298, 1790–1793. [Google Scholar] [CrossRef]
- Alemka, A.; Nothaft, H.; Zheng, J.; Szymanski, C.M. N-glycosylation of Campylobacter jejuni surface proteins promotes bacterial fitness. Infect. Immun. 2013, 81, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- Cain, J.A.; Dale, A.L.; Niewold, P.; Klare, W.P.; Man, L.; White, M.Y.; Scott, N.E.; Cordwell, S.J. Proteomics Reveals Multiple Phenotypes Associated with N-linked Glycosylation in Campylobacter jejuni. Mol. Cell. Proteom. 2019, 18, 715–734. [Google Scholar] [CrossRef]
- Thibault, P.; Logan, S.M.; Kelly, J.F.; Brisson, J.R.; Ewing, C.P.; Trust, T.J.; Guerry, P. Identification of the carbohydrate moieties and glycosylation motifs in Campylobacter jejuni flagellin. J. Biol. Chem. 2001, 276, 34862–34870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, J.; Pirinccioglu, N.; Oldfield, N.J.; Carlsohn, E.; Stoof, J.; Aslam, A.; Self, T.; Cawthraw, S.A.; Petrovska, L.; Colborne, N.; et al. A novel O-linked glycan modulates Campylobacter jejuni major outer membrane protein-mediated adhesion to human histo-blood group antigens and chicken colonization. Open Biol. 2014, 4, 130202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerry, P.; Ewing, C.P.; Schirm, M.; Lorenzo, M.; Kelly, J.; Pattarini, D.; Majam, G.; Thibault, P.; Logan, S. Changes in flagellin glycosylation affect Campylobacter autoagglutination and virulence. Mol. Microbiol. 2006, 60, 299–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, C.M.; Michael, F.S.; Jarrell, H.C.; Li, J.; Gilbert, M.; Larocque, S.; Vinogradov, E.; Brisson, J.R. Detection of conserved N-linked glycans and phase-variable lipooligosaccharides and capsules from Campylobacter cells by mass spectrometry and high resolution magic angle spinning NMR spectroscopy. J. Biol. Chem. 2003, 278, 24509–24520. [Google Scholar] [CrossRef] [Green Version]
- Karlyshev, A.V.; Champion, O.L.; Churcher, C.; Brisson, J.R.; Jarrell, H.C.; Gilbert, M.; Brochu, D.; St Michael, F.; Li, J.; Wakarchuk, W.W.; et al. Analysis of Campylobacter jejuni capsular loci reveals multiple mechanisms for the generation of structural diversity and the ability to form complex heptoses. Mol. Microbiol. 2005, 55, 90–103. [Google Scholar] [CrossRef]
- Bacon, D.J.; Szymanski, C.S.; Burr, D.H.; Silver, R.P.; Alm, R.A.; Guerry, P. A phase-variable capsule is involved in virulence of Campylobacter jejuni 81–176. Mol. Microbiol. 2001, 40, 769–777. [Google Scholar] [CrossRef]
- Wong, A.; Lange, D.; Houle, S.; Arbatsky, N.P.; Valvano, M.A.; Knirel, Y.A.; Dozois, C.M.; Creuzenet, C. Role of capsular modified heptose in the virulence of Campylobacter jejuni. Mol. Microbiol. 2015, 96, 1136–1158. [Google Scholar] [CrossRef] [Green Version]
- Candon, H.L.; Allan, B.J.; Fraley, C.D.; Gaynor, E.C. Polyphosphate kinase 1 is a pathogenesis determinant in Campylobacter jejuni. J. Bacteriol. 2007, 189, 8099–8108. [Google Scholar] [CrossRef] [Green Version]
- Gangaiah, D.; Liu, Z.; Arcos, J.; Kassem, I.I.; Sanad, Y.; Torrelles, J.B.; Rajashekara, G. Polyphosphate kinase 2: A novel determinant of stress responses and pathogenesis in Campylobacter jejuni. PLoS ONE 2010, 5, e12142. [Google Scholar] [CrossRef]
- Drozd, M.; Gangaiah, D.; Liu, Z.; Rajashekara, G. Contribution of TAT system translocated PhoX to Campylobacter jejuni phosphate metabolism and resilience to environmental stresses. PLoS ONE 2011, 6, e26336. [Google Scholar] [CrossRef] [Green Version]
- Stetsenko, V.V.; Efimochkina, N.R.; Pichugina, T.V. Growth and Persistence of Campylobacter jejuni in Foodstuffs. Bull. Exp. Biol. Med. 2019, 166, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Sulaeman, S.; Hernould, M.; Schaumann, A.; Coquet, L.; Bolla, J.M.; De, E.; Tresse, O. Enhanced adhesion of Campylobacter jejuni to abiotic surfaces is mediated by membrane proteins in oxygen-enriched conditions. PLoS ONE 2012, 7, e46402. [Google Scholar] [CrossRef] [PubMed]
- Gundogdu, O.; da Silva, D.T.; Mohammad, B.; Elmi, A.; Mills, D.C.; Wren, B.W.; Dorrell, N. The Campylobacter jejuni MarR-like transcriptional regulators RrpA and RrpB both influence bacterial responses to oxidative and aerobic stresses. Front. Microbiol. 2015, 21, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turonova, H.; Briandet, R.; Rodrigues, R.; Hernould, M.; Hayek, N.; Stintzi, A.; Pazlarova, J.; Tresse, O. Biofilm spatial organization by the emerging pathogen Campylobacter jejuni: Comparison between NCTC 11168 and 81–176 strains under microaerobic and oxygen-enriched conditions. Front. Microbiol. 2015, 13, 709. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Jeon, B. Role of alkyl hydroperoxide reductase (AhpC) in the biofilm formation of Campylobacter jejuni. PLoS ONE 2014, 9, e87312. [Google Scholar] [CrossRef] [Green Version]
- Reeser, R.J.; Medler, R.T.; Billington, S.J.; Jost, B.H.; Joens, L.A. Characterization of Campylobacter jejuni biofilms under defined growth conditions. Appl. Environ. Microbiol. 2007, 73, 1908–1913. [Google Scholar] [CrossRef] [Green Version]
- Teh, A.; Lee, S.; Dykes, G. Growth in the presence of specific antibiotics induces biofilm formation by a Campylobacter jejuni strain sensitive to them but not in resistant strains. J. Glob. Antimicrob. Resist. 2019, 18, 55–58. [Google Scholar] [CrossRef]
- Day, C.J.; Tiralongo, J.; Hartnell, R.D.; Logue, C.A.; Wilson, J.C.; von Itzstein, M.; Korolik, V. Differential Carbohydrate Recognition by Campylobacter jejuni Strain 11168: Influences of Temperature and Growth Conditions. PLoS ONE 2009, 4, e4927. [Google Scholar] [CrossRef] [Green Version]
- Stahl, M.; Friis, L.M.; Nothaft, H.; Liu, X.; Li, J.; Szymanski, C.M.; Stintzi, A. L-fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc. Natl. Acad. Sci. USA 2011, 108, 7194–7199. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.Q.; Boothe, D.H.; Frank, J.F.; Arnold, J.W. Culture and detection of Campylobacter jejuni within mixed microbial populations of biofilms on stainless steel. J. Food Prot. 2007, 70, 1379–1385. [Google Scholar] [CrossRef]
- Teh, A.; Lee, S.; Dykes, G. Association of some Campylobacter jejuni with Pseudomonas aeruginosa biofilms increases attachment under conditions mimicking those in the environment. PLoS ONE 2019, 14, e0215275. [Google Scholar] [CrossRef] [PubMed]
- Engebrecht, J.; Silverman, M. Identification of genes and gene products necessary for bacterial bioluminescence. Proc. Natl. Acad. Sci. USA 1984, 81, 4154–4158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: Isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef]
- Higgins, D.A.; Pomianek, M.E.; Kraml, C.M.; Taylor, R.K.; Semmelhack, M.F.; Bassler, B.L. The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 2007, 450, 883–886. [Google Scholar] [CrossRef]
- Passador, L.; Cook, J.M.; Gambello, M.J.; Rust, L.; Iglewski, B.H. Expression of Pseudomonas aeruginosa virulence genes requires cell-to-cell communication. Science 1993, 260, 1127–1130. [Google Scholar] [CrossRef]
- De Kievit, T.R. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Jain, V.; Dongre, M. Identification of a constitutively active variant of LuxO that affects production of HA/protease and biofilm development in a non-O1, non-O139 Vibrio cholerae O110. Gene 2006, 369, 126–133. [Google Scholar] [CrossRef]
- Yarwood, J.M.; Schlievert, P.M. Quorum sensing in Staphylococcus infections. J. Clin. Investig. 2003, 112, 1620–1625. [Google Scholar] [CrossRef]
- Elvers, K.T.; Park, S.F. Quorum sensing in Campylobacter jejuni: Detection of a luxS encoded signalling molecule. Microbiology 2002, 148, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.; Itoh, K.; Misawa, N.; Ryu, S. Effects of quorum sensing on flaA transcription and autoagglutination in Campylobacter jejuni. Microbiol. Immunol. 2003, 48, 833–839. [Google Scholar] [CrossRef]
- Ligowska, M.; Cohn, M.T.; Stabler, R.A.; Wren, B.W.; Brondsted, L. Effect of chicken meat environment on gene expression of Campylobacter jejuni and its relevance to survival in food. Int. J. Food Microbiol. 2011, 145, S111–S115. [Google Scholar] [CrossRef] [PubMed]
- Fields, J.A.; Li, J.Q.; Gulbronson, C.J.; Hendrixson, D.R.; Thompson, S.A. Campylobacter jejuni CsrA Regulates Metabolic and Virulence Associated Proteins and Is Necessary for Mouse Colonization. PLoS ONE 2016, 11, e0156932. [Google Scholar] [CrossRef] [PubMed]
- Fields, J.A.; Thompson, S.A. Campylobacter jejuni CsrA mediates oxidative stress responses, biofilm formation, and host cell invasion. J. Bacteriol. 2008, 190, 3411–3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gulbronson, C.J.; Bogacz, M.; Hendrixson, D.R.; Thompson, S.A. FliW controls growth-phase expression of Campylobacter jejuni flagellar and non-flagellar proteins via the post-transcriptional regulator CsrA. Microbiology 2018, 164, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Svensson, S.L.; Davis, L.M.; MacKichan, J.K.; Allan, B.J.; Pajaniappan, M.; Thompson, S.A.; Gaynor, E.C. The CprS sensor kinase of the zoonotic pathogen Campylobacter jejuni influences biofilm formation and is required for optimal chick colonization. Mol. Microbiol. 2009, 71, 253–272. [Google Scholar] [CrossRef] [Green Version]
- Teh, A.; Lee, S.; Dykes, G. Does Campylobacter jejuni Form Biofilms in Food-Related Environments? Appl. Environ. Microbiol. 2014, 80, 5154–5160. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.H.; Rambaugh, K.; Passador, L.; Davies, D.G.; Hamood, A.N.; Iglewski, B.H.; Kornberg, A. Polyphosphate kinase is essential for biofilm development, quorum sensing, and virulence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 9636–9641. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.L.; Reuter, M.; Salt, L.J.; Cross, K.L.; Betts, R.P.; van Vliet, A.H. Chicken juice enhances surface attachment and biofilm formation of Campylobacter jejuni. Appl. Environ. Microbiol. 2014, 80, 7053–7060. [Google Scholar] [CrossRef] [Green Version]
- Karki, A.B.; Wells, H.; Fakhr, M.K. Retail liver juices enhance the survivability of Campylobacter jejuni and Campylobacter coli at low temperatures. Sci. Rep. 2019, 9, 2733. [Google Scholar] [CrossRef]
- Hanning, I.; Donoghue, D.J.; Jarquin, R.; Kumar, G.S.; Aguiar, V.F.; Metcalf, J.H.; Reyes-Herrera, I.; Slavik, M. Campylobacter biofilm phenotype exhibits reduced colonization potential in young chickens and altered in vitro virulence. Poult. Sci. 2009, 88, 1102–1107. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tram, G.; Day, C.J.; Korolik, V. Bridging the Gap: A Role for Campylobacter jejuni Biofilms. Microorganisms 2020, 8, 452. https://doi.org/10.3390/microorganisms8030452
Tram G, Day CJ, Korolik V. Bridging the Gap: A Role for Campylobacter jejuni Biofilms. Microorganisms. 2020; 8(3):452. https://doi.org/10.3390/microorganisms8030452
Chicago/Turabian StyleTram, Greg, Christopher J. Day, and Victoria Korolik. 2020. "Bridging the Gap: A Role for Campylobacter jejuni Biofilms" Microorganisms 8, no. 3: 452. https://doi.org/10.3390/microorganisms8030452