Liver Transudate, a Potential Alternative to Detect Anti-Hepatitis E Virus Antibodies in Pigs and Wild Boars (Sus scrofa)

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Samples
2.2. Detection of Anti-HEV Antibodies by ELISA
2.3. Detection of HEV RNA by RT-qPCR
2.4. Statistical Analysis
3. Results
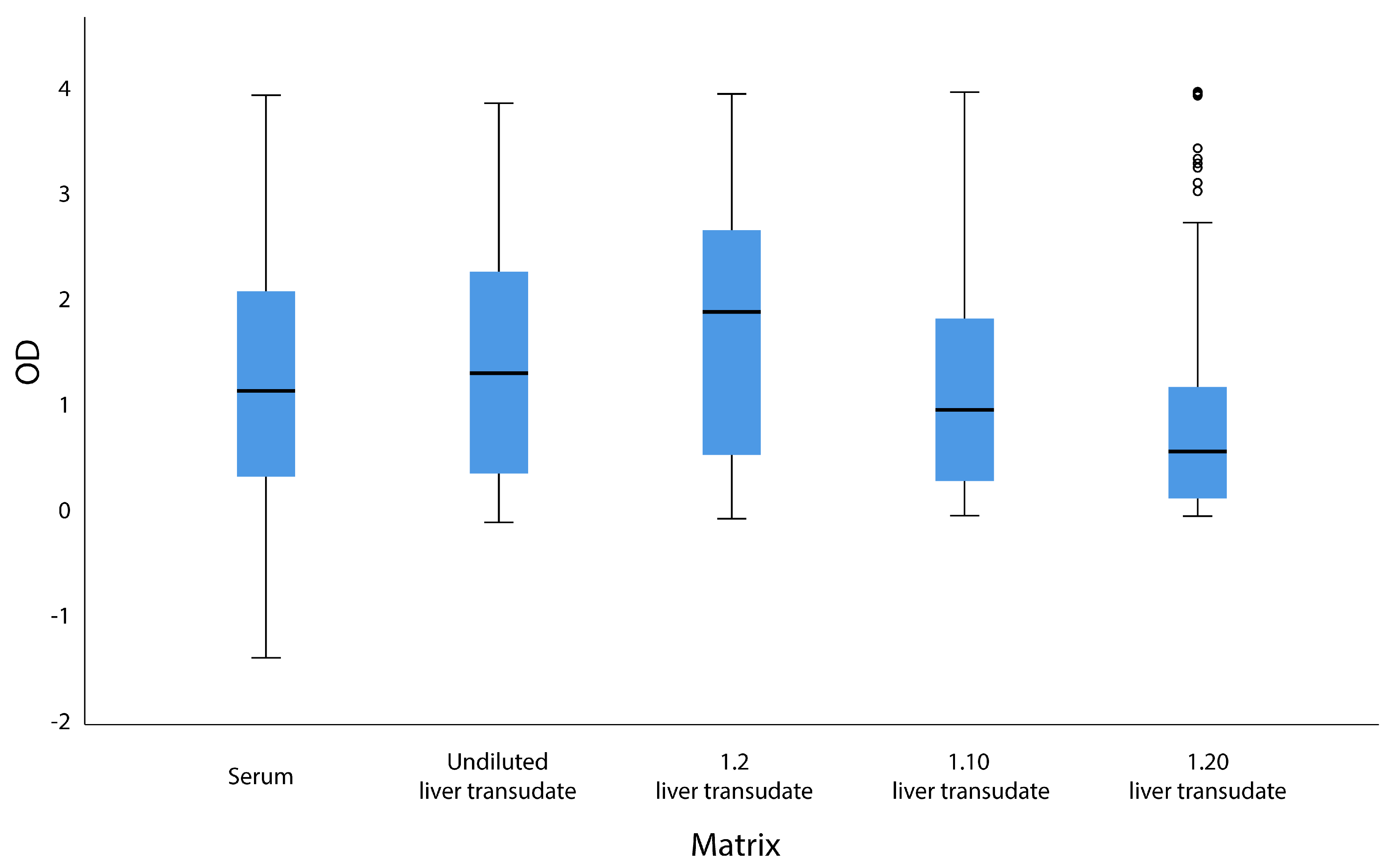
3.1. Serological Results
3.2. RT-qPCR Detection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Balayan, M.S.; Andjaparidze, A.G.; Savinskaya, S.S.; Ketiladze, E.S.; Braginsky, D.M.; Savinov, A.P.; Poleschuk, V.F. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology 1983, 20, 23–31. [Google Scholar] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Martinez-Peinado, A.; Risalde, M.A.; Rodriguez-Cano, D.; Camacho, A.; García-Bocanegra, I.; Cuenca-López, F.; Gomez-Villamandos, J.C.; Rivero, A. Familial Hepatitis E Outbreak Linked to Wild Boar Meat Consumption. Zoonoses Public Health. 2017, 64, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Pavio, N.; Meng, X.J.; Renou, C. Zoonotic hepatitis E: Animal reservoirs and emerging risks. Vet Res. 2010, 41, 46. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Istrate, C.; Santos-Ferreira, N.L.; Ferreira, A.S.; Abreu-Silva, J.; Veiga, J.; van der Poel, W.H.M.; Nascimento, M.S.J. Short communication: Detection and molecular characterization of hepatitis E virus in domestic animals of Sao Tome and Principe. Trop. Anim. Health Prod. 2019, 51, 481–485. [Google Scholar] [CrossRef]
- Hammerschmidt, F.; Schwaiger, K.; Dahnert, L.; Vina-Rodriguez, A.; Hoper, D.; Gareis, M.; Groschup, M.H.; Eiden, M. Hepatitis E virus in wild rabbits and European brown hares in Germany. Zoonoses Public Health. 2017, 64, 612–622. [Google Scholar] [CrossRef]
- Kukielka, D.; Rodriguez-Prieto, V.; Vicente, J.; Sanchez-Vizcaino, J.M. Constant Hepatitis E Virus (HEV) Circulation in Wild Boar and Red Deer in Spain: An Increasing Concern Source of HEV Zoonotic Transmission. Transbound. Emerg. Dis. 2016, 63, e360–e368. [Google Scholar] [CrossRef] [Green Version]
- Spahr, C.; Ryll, R.; Knauf-Witzens, T.; Vahlenkamp, T.W.; Ulrich, R.G.; Johne, R. Serological evidence of hepatitis E virus infection in zoo animals and identification of a rodent-borne strain in a Syrian brown bear. Vet. Microbiol. 2017, 212, 87–92. [Google Scholar] [CrossRef]
- Rui, P.; Zhao, F.; Yan, S.; Wang, C.; Fu, Q.; Hao, J.; Zhou, X.; Zhong, H.; Tang, M.; Hui, W.; et al. Detection of hepatitis E virus genotypes 3 and 4 in donkeys in northern China. Equine Vet. J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Song, S.; Park, G.N.; Shin, J.; Kim, K.S.; Cha, R.M.; Hyun, B.H.; Park, B.K.; An, D.J. Detection of subtypes (3a, 4a and 4d) and high prevalence of hepatitis E virus in Korean wild boar. Vet. Microbiol. 2020, 240, 108531. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.J.; Preiss, J.; Schemmerer, M.; Huber, B.; Plentz, A.; Jilg, W. Detection of hepatitis E virus (HEV) from porcine livers in Southeastern Germany and high sequence homology to human HEV isolates. J. Clin. Virol. 2011, 52, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Boadella, M.; Casas, M.; Martin, M.; Vicente, J.; Segales, J.; de la Fuente, J.; Gortazar, C. Increasing contact with hepatitis E virus in red deer, Spain. Emerg. Infect. Dis. 2010, 16, 1994–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schielke, A.; Ibrahim, V.; Czogiel, I.; Faber, M.; Schrader, C.; Dremsek, P.; Ulrich, R.G.; Johne, R. Hepatitis E virus antibody prevalence in hunters from a district in Central Germany, 2013: A cross-sectional study providing evidence for the benefit of protective gloves during disembowelling of wild boars. BMC Infect. Dis. 2015, 15, 440. [Google Scholar] [CrossRef] [Green Version]
- Perez-Gracia, M.T.; Mateos, M.L.; Galiana, C.; Fernandez-Barredo, S.; Garcia, A.; Gomez, M.T.; Moreira, V. Autochthonous hepatitis E infection in a slaughterhouse worker. Am. J. Trop. Med. Hyg. 2007, 77, 893–896. [Google Scholar] [CrossRef] [Green Version]
- Kantala, T.; Kinnunen, P.M.; Oristo, S.; Jokelainen, P.; Vapalahti, O.; Maunula, L. Hepatitis E Virus Antibodies in Finnish Veterinarians. Zoonoses Public Health. 2017, 64, 232–238. [Google Scholar] [CrossRef]
- Feagins, A.R.; Opriessnig, T.; Guenette, D.K.; Halbur, P.G.; Meng, X.J. Detection and characterization of infectious Hepatitis E virus from commercial pig livers sold in local grocery stores in the USA. J. Gen. Virol. 2007, 88, 912–917. [Google Scholar] [CrossRef]
- Khuroo, M.S.; Khuroo, M.S.; Khuroo, N.S. Hepatitis E: Discovery, global impact, control and cure. World J Gastroentero. 2016, 22, 7030–7045. [Google Scholar] [CrossRef]
- Van der Poel, W.H. Food and environmental routes of Hepatitis E virus transmission. Curr. Opin. Virol. 2014, 4, 91–96. [Google Scholar] [CrossRef]
- Huang, F.; Li, Y.; Yu, W.; Jing, S.; Wang, J.; Long, F.; He, Z.; Yang, C.; Bi, Y.; Cao, W.; et al. Excretion of infectious hepatitis E virus into milk in cows imposes high risks of zoonosis. Hepatology 2016, 64, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domanovic, D.; Tedder, R.; Blumel, J.; Zaaijer, H.; Gallian, P.; Niederhauser, C.; Sauleda Oliveras, S.; O´Riordan, J.; Boland, F.; Harritshoj, L.; et al. Hepatitis E and blood donation safety in selected European countries: A shift to screening? Eurosurveillance 2017, 22, 30514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvala, H.; Hewitt, P.E.; Reynolds, C.; Pearson, C.; Haywood, B.; Tettmar, K.I.; Ushiro-Lumb, I.; Braislford, S.R.; Tedder, R.; Ijaz, S. Hepatitis E virus in blood donors in England, 2016 to 2017: From selective to universal screening. Eurosurveillance 2019, 24, 1800386. [Google Scholar] [CrossRef]
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Gottingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, J.; Sheng, Y.; Lu, Q.; Liu, B.; Chen, Y.; Sun, Y.; Zhou, E.M.; Zhao, Q. Prevalence of hepatitis E virus (HEV) infection in various pig farms from Shaanxi Province, China: First detection of HEV RNA in pig semen. Transbound. Emerg. Dis. 2019, 66, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Nishizawa, T.; Nagashima, S.; Jirintai, S.; Kawakami, M.; Sonoda, Y.; Suzuki, T.; Yamamoto, S.; Shigemoto, K.; Ashida, K.; et al. Molecular characterization of a novel hepatitis E virus (HEV) strain obtained from a wild boar in Japan that is highly divergent from the previously recognized HEV strains. Virus Res. 2014, 180, 59–69. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar] [CrossRef]
- Colson, P.; Borentain, P.; Motte, A.; Lagrange, X.; Kaba, M.; Henry, M.; Tamalet, C.; Gerolami, R. First human cases of hepatitis E infection with genotype 3c strains. J. Clin. Virol. 2007, 40, 318–320. [Google Scholar] [CrossRef]
- Kamar, N.; Garrouste, C.; Haagsma, E.B.; Garrigue, V.; Pischke, S.; Chauvet, C.; Dumortier, J.; Cannesson, A.; Cassuto-Viguier, E.; Thervet, E.; et al. Factors associated with chronic hepatitis in patients with hepatitis E virus infection who have received solid organ transplants. Gastroenterology 2011, 140, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Scharn, N.; Ganzenmueller, T.; Wenzel, J.J.; Dengler, R.; Heim, A.; Wegner, F. Guillain-Barre syndrome associated with autochthonous infection by hepatitis E virus subgenotype 3c. Infection 2014, 42, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Deroux, A.; Brion, J.P.; Hyerle, L.; Belbezier, A.; Vaillant, M.; Mosnier, E.; Larrat, S.; Morand, P.; Pavese, P. Association between hepatitis E and neurological disorders: Two case studies and literature review. J. Clin. Virol. 2014, 60, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Weclawiak, H.; Guilbeau-Frugier, C.; Legrand-Abravanel, F.; Cointault, O.; Ribes, D.; Esposito, L.; Cardeau-Desangles, I.; Guitard, J.; Sallusto, F.; et al. Hepatitis E virus and the kidney in solid-organ transplant patients. Transplantation 2012, 93, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [Green Version]
- Ponterio, E.; Di Bartolo, I.; Orru, G.; Liciardi, M.; Ostanello, F.; Ruggeri, F.M. Detection of serum antibodies to hepatitis E virus in domestic pigs in Italy using a recombinant swine HEV capsid protein. BMC Vet. Res. 2014, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, M.; Roche, S.J.; Sammin, D. Seroprevalence of Hepatitis E virus infection in the Irish pig population. Irish. Vet. J. 2015, 68, 8. [Google Scholar] [CrossRef] [Green Version]
- Salines, M.; Andraud, M.; Rose, N. From the epidemiology of hepatitis E virus (HEV) within the swine reservoir to public health risk mitigation strategies: A comprehensive review. Vet. Res. 2017, 48, 31. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Okada, K.; Takahashi, K.; Mishiro, S. Severe hepatitis E virus infection after ingestion of uncooked liver from a wild boar. J. Infect. Dis. 2003, 188, 944. [Google Scholar] [CrossRef] [Green Version]
- Adlhoch, C.; Wolf, A.; Meisel, H.; Kaiser, M.; Ellerbrok, H.; Pauli, G. High HEV presence in four different wild boar populations in East and West Germany. Vet. Microbiol. 2009, 139, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Burri, C.; Vial, F.; Ryser-Degiorgis, M.P.; Schwermer, H.; Darling, K.; Reist, M.; Wu, N.; Beerli, O.; Schoning, J.; Cavassini, M.; et al. Seroprevalence of hepatitis E virus in domestic pigs and wild boars in Switzerland. Zoonoses Public Health. 2014, 61, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, C.; Modesto, P.; Bertolini, S.; Peletto, S.; Acutis, P.L.; Dondo, A.; Robetto, S.; Mignone, W.; Orusa, R.; Ru, G.; et al. Serological and virological survey of hepatitis E virus in wild boar populations in northwestern Italy: Detection of HEV subtypes 3e and 3f. Arch. Virol. 2015, 160, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, J.; Vina-Rodriguez, A.; Fast, C.; Groschup, M.H.; Eiden, M. Chronically infected wild boar can transmit genotype 3 hepatitis E virus to domestic pigs. Vet. Microbiol. 2015, 180, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Khudyakov, Y.; Kamili, S. Serological diagnostics of hepatitis E virus infection. Virus Res. 2011, 161, 84–92. [Google Scholar] [CrossRef]
- Casas, M.; Cortes, R.; Pina, S.; Peralta, B.; Allepuz, A.; Cortey, M.; Casal, J.; Martín, M. Longitudinal study of hepatitis E virus infection in Spanish farrow-to-finish swine herds. Vet. Microbiol. 2011, 148, 27–34. [Google Scholar] [CrossRef]
- Sommerkorn, F.M.; Schauer, B.; Schreiner, T.; Fickenscher, H.; Krumbholz, A. Performance of Hepatitis E Virus (HEV)-antibody tests: A comparative analysis based on samples from individuals with direct contact to domestic pigs or wild boar in Germany. Med. Microbiol. Immun. 2017, 206, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Dahnert, L.; Conraths, F.J.; Reimer, N.; Groschup, M.H.; Eiden, M. Molecular and serological surveillance of Hepatitis E virus in wild and domestic carnivores in Brandenburg, Germany. Transbound. Emerg. Dis. 2018, 65, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Wacheck, S.; Werres, C.; Mohn, U.; Dorn, S.; Soutschek, E.; Fredriksson-Ahomaa, M.; Martlbauer, E. Detection of IgM and IgG against hepatitis E virus in serum and meat juice samples from pigs at slaughter in Bavaria, Germany. Foodborne Pathog Dis. 2012, 9, 655–660. [Google Scholar] [CrossRef]
- Baylis, S.A.; Blumel, J.; Mizusawa, S.; Matsubayashi, K.; Sakata, H.; Okada, Y.; Nubling, C.M.; Hanschmann, K.M. World Health Organization International Standard to harmonize assays for detection of hepatitis E virus RNA. Emerg. Infect. Dis. 2013, 19, 729–735. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods. 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Rutjes, S.A.; Lodder-Verschoor, F.; Lodder, W.J.; van der Giessen, J.; Reesink, H.; Bouwknegt, M.; de Roda Husman, A.M. Seroprevalence and molecular detection of hepatitis E virus in wild boar and red deer in The Netherlands. J. Virol. Methods. 2010, 168, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; Hernandez, M.; Gutierrez-Boada, M.; Valero, A.; Navarro, A.; Munoz-Chimeno, M.; Fernández-Manzano, A.; Escobar, F.M.; Martínez, I.; Bárcena, C.; et al. Occurrence of Hepatitis E virus in pigs, pork cuts and organs at the Time of Slaughter, Spain, 2017. Front. Microbiol. 2019, 10, 2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Muller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Sing, T.; Sander, O.; Beerenwinkel, N.; Lengauer, T. ROCR: Visualizing classifier performance in R. Bioinformatics 2005, 21, 3940–3941. [Google Scholar] [CrossRef] [PubMed]
- Team RDC. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Yonemitsu, K.; Minami, S.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Maeda, K. Detection of anti-viral antibodies from meat juice of wild boars. J. Vet. Med. Sci. 2019, 81, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meemken, D.; Tangemann, A.H.; Meermeier, D.; Gundlach, S.; Mischok, D.; Greiner, M.; Klein, G.; Blaha, T. Establishment of serological herd profiles for zoonoses and production diseases in pigs by “meat juice multi-serology”. Prev. Vet. Med. 2014, 113, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Felin, E.; Nareaho, A.; Fredriksson-Ahomaa, M. Comparison of commercial ELISA tests for the detection of Toxoplasma antibodies in the meat juice of naturally infected pigs. Vet. Parasitol. 2017, 238, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Wallander, C.; Frossling, J.; Vagsholm, I.; Burrells, A.; Lunden, A. “Meat juice” is not a homogeneous serological matrix. Foodborne Pathog. Dis. 2015, 12, 280–288. [Google Scholar] [CrossRef]
- Koppelman, S.J.; Hefle, S.L. Detecting allergens in food; Woodhead Publishing Ltd.: Cambridge, UK, 2006. [Google Scholar]
- Taylor, S.L.; Nordlee, J.A.; Niemann, L.M.; Lambrecht, D.M. Allergen immunoassays—Considerations for use of naturally incurred standards. Anal. Bioanal. Chem. 2009, 395, 83–92. [Google Scholar] [CrossRef]
- Nielsen, B.; Ekeroth, L.; Bager, F.; Lind, P. Use of muscle fluid as a source of antibodies for serologic detection of Salmonella infection in slaughter pig herds. J. Vet. Diagn. Invest. Inc. 1998, 10, 158–163. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Species (n) | Positive Serum Samples, n (%) | Positive Undiluted Liver Transudate Samples, n (%) | Positive Transudate 1:2 (%) | Positive Transudate 1:10 (%) | Positive Transudate 1:20 (%) |
|---|---|---|---|---|---|
| White pig (44) | 34 (77.2) | 37 (84.0) | 38 (86.3) | 34 (77.2) | 28 (63.6) |
| Iberian pig (46) | 29 (63.0) | 20 (43.4) | 29 (63.0) | 17 (36.9) | 11 (23.9) |
| Wild boar (35) | 22 (62.8) | 20 (57.1) | 21 (60.0) | 20 (57.1) | 17 (48.5) |
| All (125) | 85 (68.0) | 77 (61.6) | 88 (70.4) | 71 (56.8) | 56 (44.8) |
| κ | 0.616 | 0.494 | 0.630 | 0.491 |
| Transudate Dilution | Samples n (%) | Specificity, % (95% IC) | Sensitivity, % (95% IC) | PPV, % (95% IC) | NPV % (95% IC) | AUC, % (95% IC) | |
|---|---|---|---|---|---|---|---|
| Undiluted | Positive | 70 (82.3) | 82.5 (70.0–92.5) | 82.3 (74.1–90.6) | 91.1 (85.5–96.0) | 69.1 (59.3–79.6) | 89.7 (83.9–95.4) |
| Negative | 33 (82.5) | ||||||
| 1:2 | Positive | 73 (85.8) | 62.5 (47.5–77.5) | 85.8 (77.6–92.9) | 82.8 (77.1–88.9) | 67.6 (55.3–81.3) | 88.5 (82.6–94.4) |
| Negative | 25 (62.5) | ||||||
| 1:10 | Positive | 67 (78.8) | 90.0 (80.0–97.5) | 78.8 (69.4–87.1) | 94.4 (89.2–98.6) | 66.7 (58.2–77.1) | 88.7 (82.2–95.2) |
| Negative | 36 (90.0) | ||||||
| 1:20 | Positive | 54 (63.5) | 95.0 (87.5–100) | 63.5 (54.1–72.9) | 96.5 (91.1–100) | 55.1 (48.1–62.9) | 89.1 (83.3–94.9) |
| Negative | 38 (95.0) | ||||||
| Species (n) | Serum Samples, n (%) | Liver Transudate Samples, n (%) |
|---|---|---|
| White pig (35) | 8 (22.8) | 0 |
| Iberian pig (5) | 0 | 0 |
| Wild boar (18) | 5 (27.7) | 4 (22.2) |
| All (58) | 13 (22.4) | 4 (6.9) |
| Sample ID | Serum Samples, Ct | Liver Transudate Samples, Ct |
|---|---|---|
| 1 | 33,27 | 27.91 |
| 2 | 41,06 | N/A |
| 3 | 28,04 | N/A |
| 4 | 37,63 | N/A |
| 5 | 36,90 | 24.98 |
| 6 | 41,08 | N/A |
| 7 | 41,96 | N/A |
| 8 | 38,96 | N/A |
| 9 | 40,57 | N/A |
| 10 | 42,21 | N/A |
| 11 | 36,90 | N/A |
| 12 | 42,62 | N/A |
| 13 | 37,28 | 32,78 |
| 14 | N/A | 40 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro, A.; Bárcena, C.; Pozo, P.; Díez-Guerrier, A.; Martínez, I.; Polo, C.; Duque, C.; Rodríguez-Lázaro, D.; Goyache, J.; García, N. Liver Transudate, a Potential Alternative to Detect Anti-Hepatitis E Virus Antibodies in Pigs and Wild Boars (Sus scrofa). Microorganisms 2020, 8, 450. https://doi.org/10.3390/microorganisms8030450
Navarro A, Bárcena C, Pozo P, Díez-Guerrier A, Martínez I, Polo C, Duque C, Rodríguez-Lázaro D, Goyache J, García N. Liver Transudate, a Potential Alternative to Detect Anti-Hepatitis E Virus Antibodies in Pigs and Wild Boars (Sus scrofa). Microorganisms. 2020; 8(3):450. https://doi.org/10.3390/microorganisms8030450
Chicago/Turabian StyleNavarro, Alejandro, Carmen Bárcena, Pilar Pozo, Alberto Díez-Guerrier, Irene Martínez, Coral Polo, Clara Duque, David Rodríguez-Lázaro, Joaquín Goyache, and Nerea García. 2020. "Liver Transudate, a Potential Alternative to Detect Anti-Hepatitis E Virus Antibodies in Pigs and Wild Boars (Sus scrofa)" Microorganisms 8, no. 3: 450. https://doi.org/10.3390/microorganisms8030450