Limited Genetic Diversity of blaCMY-2-Containing IncI1-pST12 Plasmids from Enterobacteriaceae of Human and Broiler Chicken Origin in The Netherlands

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Isolates
2.1.1. AmpC E. coli Isolates from i-4-1-Health Dutch-Belgian Cross-Border Project
2.1.2. AmpC E. coli Isolates from Amphia Prevalence Screening
2.1.3. pAmpC-encoding Clinical Isolates from Elisabeth-Tweesteden Hospital
2.2. Whole-Genome Bioinformatics Analysis of Short-Read Sequencing Data
2.3. Long-Read Sequencing and Hybrid Assembly
2.4. Plasmid Analysis
2.5. Classification of Pairwise Comparisons
2.6. Ethical Statement
3. Results
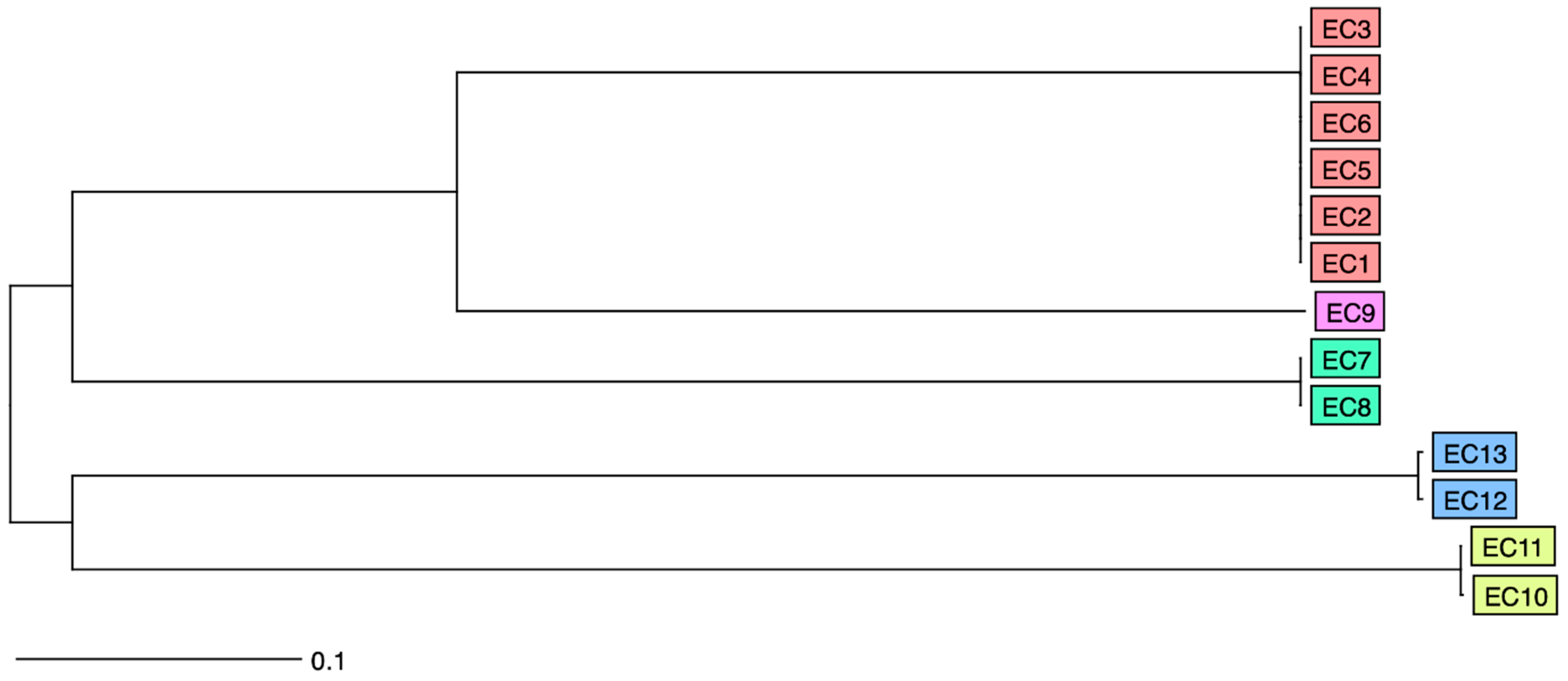
3.1. Isolate Characteristics
3.2. Plasmid Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davies, J. Origins and evolution of antibiotic resistance. Microbiologia 1996, 12, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premanandh, J.; Samara, B.S.; Mazen, A.N. Race Against Antimicrobial Resistance Requires Coordinated Action – An Overview. Front. Microbiol. 2016, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carlet, J. The gut is the epicentre of antibiotic resistance. Antimicrob. Resist. Infect. Control. 2012, 1, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A. Animal reservoirs for extended spectrum β-lactamase producers. Clin. Microbiol. Infect. 2008, 14, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Dierikx, C.; van der Goot, J.; Fabri, T.; van Essen-Zandbergen, A.; Smith, H.; Mevius, D. Extended-spectrum-β-lactamase- and AmpC-β-lactamase-producing Escherichia coli in Dutch broilers and broiler farmers. J. Antimicrob. Chemother. 2013, 68, 60–67. [Google Scholar] [CrossRef]
- Berg, E.S.; Wester, A.L.; Ahrenfeldt, J.; Mo, S.S.; Slettemeås, J.S.; Steinbakk, M.; Samuelsen, Ø.; Grude, N.; Simonsen, G.S.; Løhr, I.H.; et al. Norwegian patients and retail chicken meat share cephalosporin-resistant Escherichia coli and IncK/blaCMY-2 resistance plasmids. Clin. Microbiol. Infect. 2017, 23, 407.e9–407.e15. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. AmpC Β-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [Green Version]
- Accogli, M.; Fortini, D.; Giufrè, M.; Graziani, C.; Dolejska, M.; Carattoli, A.; Cerquetti, M. IncI1 plasmids associated with the spread of CMY-2, CTX-M-1 and SHV-12 in Escherichia coli of animal and human origin. Clin. Microbiol. Infect. 2013, 19, E238–E240. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.H.; Bortolaia, V.; Nielsen, C.A.; Nielsen, J.B.; Schønning, K.; Agersø, Y.; Guardabassia, L. Host-specific patterns of genetic diversity among IncI1-Iγ and IncK plasmids encoding CMY-2 β -lactamase in escherichia coli isolates from humans, poultry meat, poultry, and dogs in Denmark. Appl. Environ. Microbiol. 2016, 82, 4705–4714. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Villa, L.; Fortini, D.; García-Fernández, A. Contemporary IncI1 plasmids involved in the transmission and spread of antimicrobial resistance in Enterobacteriaceae. Plasmid 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Johannesen, T.B.; Stegger, M.; Bortolaia, V.; Leekitcharoenphon, P.; Korsgaard, H.B.; Seyfarth, A.M.; Mossong, J.; et al. ST131 fimH22 Escherichia coli isolate with a blaCMY-2/IncI1/ST12 plasmid obtained from a patient with bloodstream infection: Highly similar to E. coli isolates of broiler origin. J. Antimicrob. Chemother. 2019, 74, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, M.; Irrgang, A.; Roschanski, N.; Brenner Michael, G.; Hamprecht, A.; Rieber, H.; Käsbohrer, A.; Schwarz, S.; Rösler, U.; Kreienbrock, L.; et al. Whole genome analyses of CMY-2-producing Escherichia coli isolates from humans, animals and food in Germany. BMC Genomics 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, L.R.; van der Graaf-Van Bloois, L.; Donado-Godoy, P.; Mevius, D.J.; Wagenaar, J.A.; Hordijk, J.; Zomer, A.L. Phylogenomic investigation of IncI1-I plasmids harboring blaCMY-2 and blaSHV-12 in salmonella enterica and Escherichia coli in multiple countries. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcek, A.; Roer, L.; Overballe-Petersen, S.; Hansen, F.; Bortolaia, V.; Leekitcharoenphon, P.; Korsgaard, H.B.; Seyfarth, A.M.; Hendriksen, R.S.; Hasman, H.; et al. IncI1 ST3 and IncI1 ST7 plasmids from CTX-M-1-producing Escherichia coli obtained from patients with bloodstream infections are closely related to plasmids from E. coli of animal origin. J. Antimicrob. Chemother. 2019, 74, 2171–2175. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, T.; Sekizuka, T.; Kuroda, M.; Suzuki, S.; Ozawa, M.; Abo, H.; Furuya, Y.; Akama, R.; Matsuda, M.; Shimazaki, Y.; et al. Comparative genomic analysis of third-generation- cephalosporin-resistant escherichia coli harboring the blaCMY-2- Positive IncI1 Group, IncB/O/K/Z, and IncC plasmids isolated from healthy broilers in Japan. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Stohr, J.J.J.M.; den Bergh, M.F.Q.K.; Wedema, R.; Friedrich, A.W.; Kluytmans, J.A.J.W.; Rossen, J.W.A. Detection of extended-spectrum beta-lactamase (ESBL) genes and plasmid replicons in Enterobacteriaceae using PlasmidSPAdes assembly of short-read sequence data. bioRxiv 2019, 863316. [Google Scholar] [CrossRef]
- Den Bergh, M.K.; Lammens, C.; Selva, N.P.; Buiting, A.; Leroux-roels, I.; Saegeman, V.; Savelkoul, P. Microbiological methods to detect intestinal carriage of highly-resistant microorganisms (HRMO) in humans and livestock in the i-4-1-Health Dutch- Belgian cross-border project. Preprints 2019, 1–16. [Google Scholar] [CrossRef]
- Den Drijver, E.; Verweij, J.J.; Verhulst, C.; Soer, J.; Veldman, K.; Kooistra-Smid, A.; Kluytmans-van den Bergh, M.F.Q.; Kluytmans, J.W. Detection of AmpC β-lactamases in Escherichia coli using different screening Evert den Drijver. BioRxiv 2019. [Google Scholar] [CrossRef]
- Coolen, J.P.M.; Den Drijver, E.P.M.; Kluytmans, J.A.J.W.; Verweij, J.J.; Lamberts, B.A.; Soer, J.A.C.J.; Verhulst, C.; Wertheim, H.F.L.; Kolwijck, E. Development of an algorithm to discriminate between plasmid- and chromosomal-mediated AmpC β-lactamase production in Escherichia coli by elaborate phenotypic and genotypic characterization. J. Antimicrob. Chemother. 2019, 74, 3481–3488. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Drijver, E.; Verweij, J.J.; Verhulst, C.; Oome, S.; Soer, J.; Willemsen, I.; Schrauwen, E.J.A.; Van Den Bergh, M.F.Q.K.; Kluytmans, J.A.J.W. Decline in AmpC β-lactamase-producing escherichia coli in a Dutch teaching hospital (2013-2016). PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluytmans-Van Den Bergh, M.F.Q.; Rossen, J.W.A.; Bruijning-Verhagen, P.C.J.; Bonten, M.J.M.; Friedrich, A.W.; Vandenbroucke-Grauls, C.M.J.E.; Willems, R.J.L.; Kluytmans, J.A.J.W. Whole-genome multilocus sequence typing of extended-spectrum-beta-lactamase-producing enterobacteriaceae. J. Clin. Microbiol. 2016, 54, 2919–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. Progressivemauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwer, M.S.M.; Tagg, K.A.; Mevius, D.J.; Iredell, J.R.; Bossers, A.; Smith, H.E.; Partridge, S.R. IncI shufflons: Assembly issues in the next-generation sequencing era. Plasmid 2015, 80, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, M.S.M.; Jurburg, S.D.; Harders, F.; Kant, A.; Mevius, D.J.; Roberts, A.P.; Bossers, A. The shufflon of IncI1 plasmids is rearranged constantly during different growth conditions. Plasmid 2019, 102, 51–55. [Google Scholar] [CrossRef]
- Wendorf, K.A.; Kay, M.; Baliga, C.; Weissman, S.J.; Gluck, M.; Verma, P.; D’Angeli, M.; Swoveland, J.; Kang, M.G.; Eckmann, K.; et al. Endoscopic Retrograde Cholangiopancreatography-Associated AmpC Escherichia coli Outbreak. Infect. Control. Hosp. Epidemiol. 2015, 36, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.F.; Chiu, C.H.; Wang, M.H.; Wu, C.Y.; Hsieh, K.S.; Chiou, C.C. Outbreak of dysentery associated with ceftriaxone-resistant Shigella sonnei: First report of plasmid-mediated CMY-2-type AmpC β-lactamase resistance in S. sonnei. J. Clin. Microbiol. 2005, 43, 2608–2612. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, M.; Yabata, J.; Nomura, Y.; Tominaga, K. Detection of CMY-2 AmpC β-lactamase-producing enterohemorrhagic Escherichia coli O157: H7 from outbreak strains in a nursery school in Japan. J. Infect. Chemother. 2015, 21, 544–546. [Google Scholar] [CrossRef]
- Matsumura, Y.; Tanaka, M.; Yamamoto, M.; Nagao, M.; Machida, K.; Ito, Y.; Takakura, S.; Ogawa, K.; Yoshizawa, A.; Fujimoto, Y.; et al. High prevalence of carbapenem resistance among plasmid-mediated AmpC β-lactamase-producing Klebsiella pneumoniae during outbreaks in liver transplantation units. Int. J. Antimicrob. Agents 2015, 45, 33–40. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolate | Species | Multilocus ST a | wgMLST Cluster | Fim | Origin | Sample Location | Flock or Ward | Sample Source | Month and Year of Isolation | Accession No. |
|---|---|---|---|---|---|---|---|---|---|---|
| EC1 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 1 | Fecal swab 1 | Nov 2017 | ERS4591617 |
| EC2 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 1 | Fecal swab 2 | Nov 2017 | ERS4591618 |
| EC3 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 2 | Fecal swab 3 | Nov 2017 | ERS4591619 |
| EC4 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 2 | Fecal swab 4 | Nov 2017 | ERS4591620 |
| EC5 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 3 | Fecal swab 5 | Nov 2017 | ERS4591621 |
| EC6 | E. coli | ST665 | 1 | fimH30 | Broiler | Farm 1 | Flock 3 | Fecal swab 6 | Nov 2017 | ERS4591622 |
| EC7 | E. coli | ST86 | 2 | fimH289 | Broiler | Farm 1 | Flock 3 | Fecal swab 5 | Nov 2017 | ERS4591623 |
| EC8 | E. coli | ST86 | 2 | fimH289 | Broiler | Farm 1 | Flock 3 | Fecal swab 7 | Nov 2017 | ERS4591624 |
| EC9 | E. coli | ST6856 | fimH71 | Broiler | Farm 1 | Flock 3 | Fecal swab 6 | Nov 2017 | ERS4591625 | |
| EC10 | E. coli | ST131 | 3 | fimH22 | Human | Hospital 1 | Ward 1 | Blood 1 | Oct 2013 | ERS4591626 |
| EC11 | E. coli | ST131 | 3 | fimH22 | Human | Hospital 2 | Ward 1 | Blood 2 | Jul 2014 | ERS4591627 |
| EC12 | E. coli | ST973 | 4 | fimH95 | Human | Hospital 3 | Ward 1 | Rectal swab 1 | Dec 2017 | ERS4591628 |
| EC13 | E. coli | ST973 | 4 | fimH95 | Human | Hospital 3 | Ward 2 | Rectal swab 2 | Dec 2017 | ERS4591629 |
| SE1 | Salmonella enteritidis | - | - | Human | Primary care unit | n.a. | Feces | Aug 2018 | ERS4591630 |
| pEC1 | pEC2 | pEC3 | pEC4 | pEC5 | pEC6 | pEC7 | pEC8 | pEC9 | pEC10 | pEC11 | pEC12 | pEC13 | pSE1 | MH472638.1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pEC1 | 0 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 9 | 8 | 6 | 2 |
| pEC2 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| pEC3 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| pEC4 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| pEC5 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| pEC6 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| pEC7 | 3 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 2 | 8 | 7 | 5 | 1 |
| pEC8 | 3 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 2 | 8 | 7 | 5 | 1 |
| pEC9 | 3 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 2 | 8 | 7 | 5 | 1 |
| pEC10 | 3 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 0 | 0 | 8 | 7 | 5 | 1 |
| pEC11 | 3 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 0 | 0 | 8 | 7 | 5 | 1 |
| pEC12 | 9 | 7 | 7 | 7 | 7 | 7 | 8 | 8 | 8 | 8 | 8 | 0 | 1 | 5 | 7 |
| pEC13 | 8 | 6 | 6 | 6 | 6 | 6 | 7 | 7 | 7 | 7 | 7 | 1 | 0 | 4 | 6 |
| pSE1 | 6 | 4 | 4 | 4 | 4 | 4 | 5 | 5 | 5 | 5 | 5 | 5 | 4 | 0 | 4 |
| MH472638.1 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 7 | 6 | 4 | 0 |
| SNP Differences | |||
|---|---|---|---|
| n of Pairwise Comparisons | Median | Range | |
| Same sample | 2 | 1 | 1 |
| Same flock, different sample | 10 | 0.5 | 0–2 |
| Same location, different ward/flock | 25 | 1 | 0–3 |
| Same domain, different location | 9 | 5 | 0–8 |
| Different domain | 45 | 4 | 1–9 |
| Plasmid | Shufflon Segments | |||
|---|---|---|---|---|
| pEC1 | PilV | A | B | rci |
| pEC2 | PilV | A | B | rci |
| pEC3 | PilV | A | B | rci |
| pEC4 | PilV | B | rci | |
| pEC5 | PilV | B | rci | |
| pEC6 | PilV | A | B | rci |
| pEC7 | PilV | B | rci | |
| pEC8 | PilV | A | B | rci |
| pEC9 | PilV | A | B | rci |
| pEC10 | PilV | A | B | rci |
| pEC11 | PilV | B | A | rci |
| pEC12 | PilV | B | rci | |
| pEC13 | PilV | B | rci | |
| pSE1 | PilV | B | rci | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drijver, E.P.M.d.; Stohr, J.J.J.M.; Verweij, J.J.; Verhulst, C.; Velkers, F.C.; Stegeman, A.; Bergh, M.F.Q.K.-v.d.; Kluytmans, J.A.J.W.; Group, i.---H.S. Limited Genetic Diversity of blaCMY-2-Containing IncI1-pST12 Plasmids from Enterobacteriaceae of Human and Broiler Chicken Origin in The Netherlands. Microorganisms 2020, 8, 1755. https://doi.org/10.3390/microorganisms8111755
Drijver EPMd, Stohr JJJM, Verweij JJ, Verhulst C, Velkers FC, Stegeman A, Bergh MFQK-vd, Kluytmans JAJW, Group i---HS. Limited Genetic Diversity of blaCMY-2-Containing IncI1-pST12 Plasmids from Enterobacteriaceae of Human and Broiler Chicken Origin in The Netherlands. Microorganisms. 2020; 8(11):1755. https://doi.org/10.3390/microorganisms8111755
Chicago/Turabian StyleDrijver, Evert P.M. den, Joep J.J.M. Stohr, Jaco J. Verweij, Carlo Verhulst, Francisca C. Velkers, Arjan Stegeman, Marjolein F.Q. Kluytmans-van den Bergh, Jan A.J.W. Kluytmans, and i---Health Study Group. 2020. "Limited Genetic Diversity of blaCMY-2-Containing IncI1-pST12 Plasmids from Enterobacteriaceae of Human and Broiler Chicken Origin in The Netherlands" Microorganisms 8, no. 11: 1755. https://doi.org/10.3390/microorganisms8111755