Molecular Epidemiology of Multi-Drug Resistant Pseudomonas aeruginosa Isolates from Hospitalized Patients in Greece


 and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Settings and Bacterial Isolates
2.2. Collection and Analysis of Clinical and Epidemiological Data
2.3. Ethical Considerations/Approval
2.4. Isolation of Genomic DNA
2.5. Antibiotic Susceptibility Testing (AST)
2.6. Screening for Acquired Resistant Genes
2.7. Molecular Typing
2.7.1. Double-Locus Sequence Typing and oprD-Typing
2.7.2. DLST and oprD Data Analysis
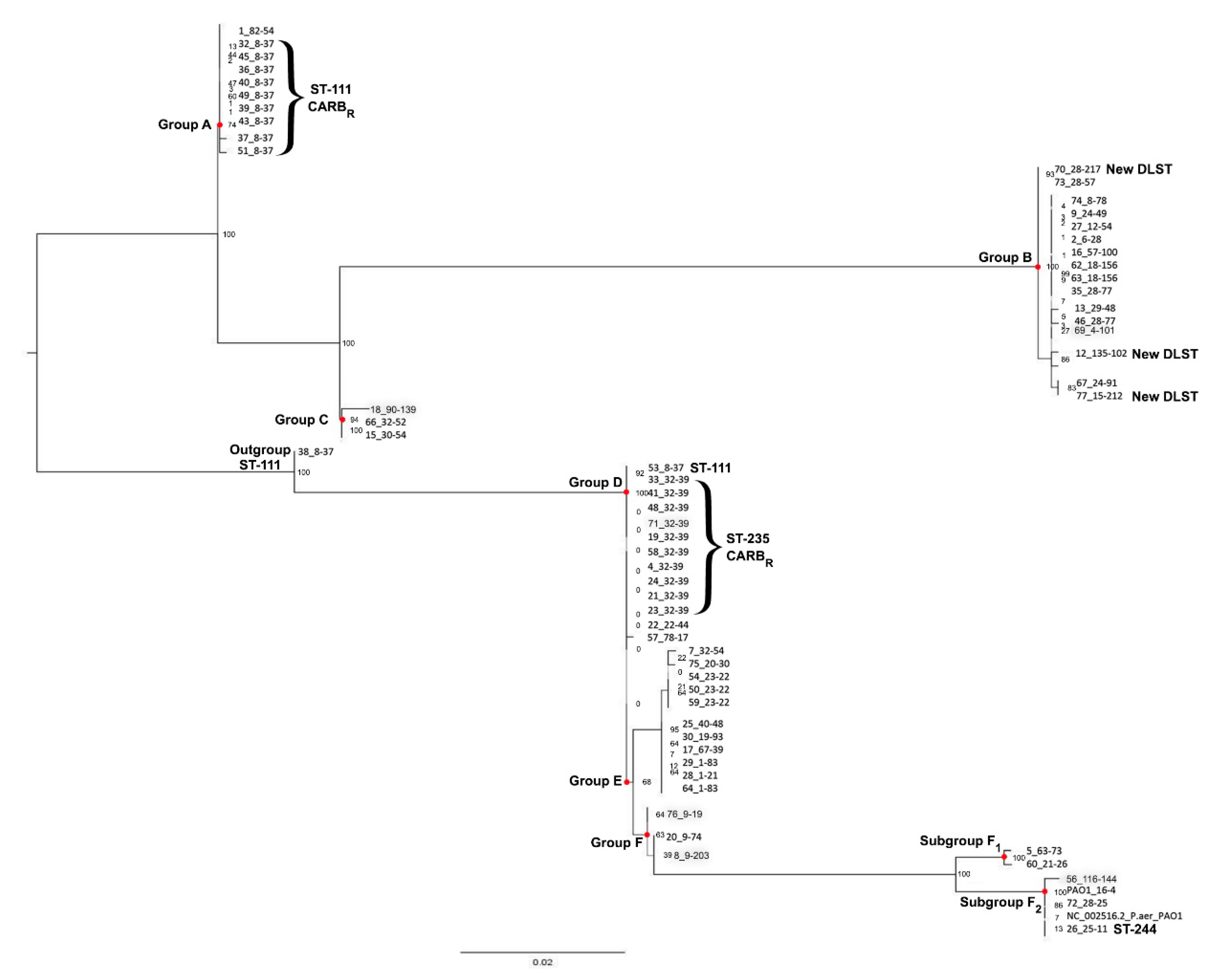
3. Results
3.1. Study Population and Characteristics
3.2. Antimicrobial Susceptibility Profiles and Detection of Resistant Genes
3.3. Molecular Typing
3.3.1. Double-Locus Sequence Typing
3.3.2. oprD Typing
3.4. Spatial and Temporal Mapping of the Main DLST Types in the two Hospital Settings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Surveillance of antimicrobial resistance in Europe. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net, ECDC). 2018. Available online: https://www.ecdc.europa.eu/en/antimicrobial-resistance/surveillance-and-disease-data/report (accessed on 18 November 2019).
- Papadimitriou-Olivgeris, M.; Fligou, F.; Spiliopoulou, A.; Koutsileou, K.; Kolonitsiou, F.; Spyropoulou, A.; Zotou, A.; Marangos, M.; Anastassiou, E.D.; Christofidou, M.; et al. Risk factors and predictors of carbapenem-resistant Pseudomonas aeruginosa and Acinetobacter baumannii mortality in critically ill bacteraemic patients over a 6-year period (2010-15): Antibiotics do matter. J. Med. Microbiol. 2017, 66, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.; Almeida, M.; Friedman, N.D.; Aragão, I.; Costa-Pereira, A.; Sarmento, A.E.; Azevedo, L. Classification of healthcare-associated infection: A systematic review 10 years after the first proposal. BMC Med. 2014, 12, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappa, O.; Vantarakis, A.; Galanis, A.; Vantarakis, G.; Mavridou, A. Antibiotic resistance profiles of Pseudomonas aeruginosa isolated from various Greek aquatic environments. FEMS Microbiol. Ecol. 2016, 92, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bălăşoiu, M.; Bălăşoiu, A.T.; Mănescu, R.; Avramescu, C.; Ionete, O. Pseudomonas aeruginosa resistance phenotypes and phenotypic highlighting methods. Curr. Health Sci. J. 2014, 40, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Meletis, G.; Bagkeri, M. Pseudomonas aeruginosa: Multi-drug-resistance development and treatment options. Infect. Control. 2013. [Google Scholar] [CrossRef] [Green Version]
- Igbinosa, E.O.; Odjadjare, E.E.; Igbinosa, I.H.; Orhue, P.O.; Omoigberale, M.N.; Amhanre, N.I. Antibiotic synergy interaction against multidrug-resistant Pseudomonas aeruginosa isolated from an abattoir effluent environment. Sci. World J. 2012, 308034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turton, J.F.; Turton, S.E.; Yearwood, L.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Evaluation of a nine-locus variable-number tandem-repeat scheme for typing of Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2010, 16, 1111–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappa, O.; Mandilara, G.; Vatopoulos, A.; Mavridou, A. Typing of Pseudomonas aeruginosa strains isolated from Greek water samples by three typing methods: Serotyping, Random Amplified Polymorphic DNA (RAPD) and Pulsed Field Gel Electrophoresis (PFGE). Water Sci. Technol. 2013, 67, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Roldán, L.; Bellés, A.; Bueno, J.; Azcona-Gutiérrez, J.M.; Rojo-Bezares, B.; Torres, C.; Castillo, F.J.; Sáenz, Y.; Seral, C. Pseudomonas aeruginosa isolates from Spanish children: Occurrence in faecal samples, antimicrobial resistance, virulence, and molecular typing. BioMed Res. Int. 2018, 2018, 8060178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, D.S.; Gomes Magalhaes, B.; Abdelbary, M.; Prod’hom, G.; Greub, G.; Wasserfallen, J.B.; Genoud, P.; Zanetti, G.; Senn, L. Hand soap contamination by Pseudomonas aeruginosa in a tertiary care hospital: No evidence of impact on patients. J. Hosp. Infect. 2016, 93, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Pappa, O.; Beloukas, A.; Vantarakis, A.; Mavridou, A.; Kefala, A.M.; Galanis, A. Molecular characterization and phylogenetic analysis of Pseudomonas aeruginosa isolates recovered from Greek aquatic habitats implementing the Double-Locus Sequence Typing Scheme. Microb. Ecol. 2017, 74, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Telling, K.; Laht, M.; Brauer, A.; Remm, M.; Kisand, V.; Maimets, M.; Tenson, T.; Lutsar, I. Multidrug resistant Pseudomonas aeruginosa in Estonian hospitals. BMC Infect Dis. 2018, 18, 513. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi-Akiyama, T.; Tada, T.; Ohmagari, N.; Viet Hung, N.; Tharavichitkul, P.; Pokhrel, B.M.; Gniadkowski, M.; Shimojima, M.; Kirikae, T. Emergence and Spread of Epidemic Multidrug-Resistant Pseudomonas aeruginosa. Genome Biol. Evol. 2017, 9, 3238–3245. [Google Scholar] [CrossRef] [PubMed]
- Basset, P.; Blanc, D.S. Fast and simple epidemiological typing of Pseudomonas aeruginosa using the double-locus sequence typing (DLST) method. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Maatallah, M.; Cheriaa, J.; Backhrouf, A.; Iversen, A.; Grundmann, H.; Do, T.; Lanotte, P.; Mastouri, M.; Elghmati, M.S.; Rojo, F.; et al. Population structure of Pseudomonas aeruginosa from five Mediterranean Countries: Evidence for frequent recombination and epidemic occurrence of CC235. PLoS ONE. 2011, 6, e25617. [Google Scholar] [CrossRef]
- Pirnay, J.P.; Bilocq, F.; Pot, B.; Cornelis, P.; Zizi, M.; Van Eldere, J.; Deschaght, P.; Vaneechoutte, M.; Jennes, S.; Pitt, T.; et al. Pseudomonas aeruginosa population structure revisited. PLoS ONE 2009, 4, e7740. [Google Scholar] [CrossRef] [Green Version]
- Estepa, V.; Rojo-Bezares, B.; Torres, C.; Sáenz, Y. Faecal carriage of Pseudomonas aeruginosa in healthy humans: Antimicrobial susceptibility and global genetic lineages. FEMS Microbiol. Ecol. 2014, 89, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Pan, Y.; Fang, Y. Role of the outer membrane protein OprD2 in carbapenem-resistance mechanisms of Pseudomonas aeruginosa. PLoS ONE. 2015, 10, e0139995. [Google Scholar] [CrossRef]
- Yan, Y.; Yao, X.; Li, H.; Zhou, Z.; Huang, W.; Stratton, C.W.; Lu, C.D.; Tang, Y.W. A novel Pseudomonas aeruginosa strain with an oprD mutation in relation to a nosocomial respiratory infection outbreak in an intensive care unit. J. Clin. Microbiol. 2014, 52, 4388–4390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cholley, P.; Stojanov, M.; Hocquet, D.; Thouverez, M.; Bertrand, X.; Blanc, D.S. Comparison of double-locus sequence typing (DLST) and multilocus sequence typing (MLST) for the investigation of Pseudomonas aeruginosa populations. Diagn. Microbiol. Infect. Dis. 2015, 82, 274–277. [Google Scholar] [CrossRef]
- Fothergill, J.L.; Upton, A.L.; Pitt, T.L.; Hart, C.A.; Winstanley, C. Diagnostic multiplex PCR assay for the identification of the Liverpool, Midlands 1 and Manchester CF epidemic strains of Pseudomonas aeruginosa. J. Cystic Fibr. 2008, 7, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EUCAST. Breakpoint Tables for Interpretation of Minimum Inhibitory Concentration (MICs) and Zone Diameters. Version 7.1. 2017. Available online: http://www.eucast:ast_of_bacteria/ (accessed on 1 December 2017).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giakkoupi, P.; Vourli, S.; Polemis, M.; Kalapothaki, V.; Tzouvelekis, L.S.; Vatopoulos, A.C. Supplementation of growth media with Zn2+ facilitates detection of VIM-2-producing Pseudomonas aeruginosa. J. Clin. Microbiol. 2008, 46, 1568–1569. [Google Scholar] [CrossRef] [Green Version]
- Aubert, D.; Poirel, L.; Chevalier, J.; Leotard, S.; Pages, J.M.; Nordmann, P. Oxacillinase-mediated resistance to cefepime and susceptibility to ceftazidime in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2001, 45, 1615–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña, C.; Suarez, C.; Tubau, F.; Juan, C.; Moya, B.; Dominguez, M.A.; Oliver, A.; Pujol, M.; Ariza, J. Nosocomial outbreak of a non-cefepime-susceptible ceftazidime-susceptible Pseudomonas aeruginosa strain overexpressing MexXY-OprM and producing an integron-borne PSE-1 betta-lactamase. J. Clin. Microbiol. 2009, 47, 2381–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquisabel, C.A.B.; Rodríguez, M.C.; Campo-Sosa, A.O.; Rodríguez, C.; Martínez-Martínez, L. Mechanisms of resistance in clinical isolates of Pseudomonas aeruginosa less susceptible to cefepime than to ceftazidime. Clin. Microbiol. Infect. 2011, 17, 1817–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EuScape. Capacity Building Workshop ‘Train the Trainer’. NSPH/CPHL. KEELPNO. 2013. Available online: http://www2.keelpno.gr/blog/?p=4650&lang=en (accessed on 20 March 2016).
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro. Surveill. 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carriço, J.A. Global optimal eBURST analysis of multilocus typing data using a graphic matroid approach. BMC Bioinform. 2009, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D.J. Modeltest 2: More models, new heuristics and parallel computing. Nat. Methods. 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaukat, A.; Almaslamani, M.; Al Soub, H.; Alkhal, A.; Abukhattab, M. Pseudomonas Aeruginosa Bacteremia in ICU: Clinical Presentation and Outcome, Emergence of MDR Strains. 2018. Available online: http://www.alliedacademies.org/journal-infectious-diseases-medical-microbiology/ (accessed on 19 October 2020).
- Hirsch, E.B.; Tam, V.H. Impact of multidrug-resistant Pseudomonas aeruginosa infection on patient outcomes. Expert Rev. Pharmacoecon Outcomes Res. 2010, 10, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strateva, T.; Yordanov, D. Pseudomonas aeruginosa—A phenomenon of bacterial resistance. J. Med. Microbiol. 2009, 58, 1133–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraki, S.; Mantadakis, E.; Nioti, E.; Samonis, G. Susceptibility of 2,252 Pseudomonas aeruginosa Clinical Isolates Over 4 Years to 9 Antimicrobials in a Tertiary Greek Hospital. Chemother. 2005, 60, 334–341. [Google Scholar] [CrossRef]
- Potter, R.F.; D’Souza, A.W.; Dantas, G. The rapid spread of carbapenem-resistant Enterobacteriaceae. Drug. Resist. Updat. 2016, 29, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Morrill, H.J.; Pogue, J.M.; Kaye, K.S.; LaPlante, K.L. Treatment Options for Carbapenem-Resistant Enterobacteriaceae Infections. OFID 2015, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, A.; Mavroidi, A.; Katsifas, E.A. Carbapenemase-producing Pseudomonas aeruginosa from central Greece: Molecular epidemiology and genetic analysis of class I integrons. BMC Infect. Dis. 2013, 13, 505. [Google Scholar] [CrossRef] [Green Version]
- Samonis, G.; Maraki, S.; Karageorgopoulos, D.E.; Vouloumanou, E.K.; Falagas, M.E. Synergy of fosfomycin with carbapenems, colistin, netilmicin, and tigecycline against multidrug-resistant Klebsiella pneumoniae, Escherichia coli, and Pseudomonas aeruginosa clinical isolates. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 695–701. [Google Scholar] [CrossRef]
- Stefani, S.; Campana, S.; Cariani, L.; Carnovale, V.; Colombo, C.; Lleo, M.M.; Iula, V.D.; Minicucci, L.; Morelli, P.; Pizzamiglio, G.; et al. Relevance of multidrug-resistant Pseudomonas aeruginosa infections in cystic fibrosis. Int. J. Med. Microbiol. 2017, 307, 353–362. [Google Scholar] [CrossRef]
- Guzek, A.; Korzeniewski, K.; Tomaszewski, D.; Rybicki, Z.; Zwolińska, E. Bacteriological Assessment of Pneumonia Caused by Gram-Negative Bacteria in Patients Hospitalized in Intensive Care Unit. Biology 2017, 955, 39–46. [Google Scholar] [CrossRef]
- Horianopoulou, M.; Legakis, N.J.; Kanellopoulou, M.; Lambropoulos, S.; Tsakris, A.; Falagas, M.E. Frequency and predictors of colonization of the respiratory tract by VIM-2-producing Pseudomonas aeruginosa in patients of a newly established intensive care unit. J. Med. Microbiol. 2006, 10, 1435–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.A.; Amyes, S.G. OXA β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P. Gram-negative bacteriae with resistance to carbapenems. Med. Sci. 2010, 26, 950–959. [Google Scholar] [CrossRef]
- Guzvinec, M.; Izdebski, R.; Butic, I.; Jelic, M.; Abram, M.; Koscak, I.; Baraniak, A.; Hryniewicz, W.; Gniadkowski, M.; Andrasevic, A.T. Sequence Types 235, 111, and 132 Predominate among Multidrug-Resistant Pseudomonas aeruginosa Clinical Isolates in Croatia. Antimicrob. Agents Chemother. 2014, 58, 6277–6283. [Google Scholar] [CrossRef] [Green Version]
- Cholley, P.; Roughyatou Ka, R.; Guyeux, C.; Thouverez, M.; Guessennd, N.; Ghebremedhin, B.; Frank, T.; Bertrand, X.; Hocquet, D. Population Structure of Clinical Pseudomonas aeruginosa from West and Central African Countries. PLoS ONE 2014, 9, e107008. [Google Scholar] [CrossRef] [Green Version]
- Cholley, P.; Thouverez, M.; Hocquet, H.; Van der Mee-Marquet, N.; Talon, D.; Bertrand, X. Most Multidrug-Resistant Pseudomonas aeruginosa Isolates from Hospitals in Eastern France Belong to a Few Clonal Types. J. Clin. Microbiol. 2011, 49, 2578–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treepong, P.; Kos, V.N.; Guyeux, C.; Blanc, D.S.; Bertrand, X.; Valot, B.; Hocquet, D. Global emergence of the widespread Pseudomonas aeruginosa ST235 clone. Clin. Microbiol. Infect. 2018, 24, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Moremi, N.; Claus, H.; Vogel, U.; Mshana, S.E. Surveillance of surgical site infections by Pseudomonas aeruginosa and strain characterization in Tanzanian hospitals does not provide proof for a role of hospital water plumbing systems in transmission. Antimicrob. Resist. Infect. Control. 2017, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyo, S.; Haldorsen, B.; Aboud, S.; Blomberg, B.; Maselle, S.Y.; Sundsfjord, A.; Langeland, N.; Samuelsen, O. Identification of VIM-2-Producing Pseudomonas aeruginosa from Tanzania is Associated with Sequence Types 244 and 640 and the Location of blaVIM-2 in a TniC Integron. Antimicrob. Agents Chemother. 2015, 59, 682–685. [Google Scholar] [CrossRef] [Green Version]
- Viedma, E.; Estepa, V.; Juan, C.; Castillo-Vera, J.; Rojo-Bezares, B.; Seral, C.; Castillo, F.J.; Sáenz, Y.; Torres, C.; Chaves, F.; et al. Comparison of local features from two Spanish hospitals reveals common and specific traits at multiple levels of the molecular epidemiology of metallo-β-lactamase-producing Pseudomonas spp. Antimicrob. Agents Chemother. 2014, 58, 2454–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, A.F.; Shahin, M.; Shokoohizadeh, L.; Halaji, M.; Shahcheraghi, F.; Ghanbari, F. Molecular epidemiology of colistin-resistant Pseudomonas aeruginosa producing NDM-1 from hospitalized patients in Iran. Iran. J. Basic Med. Sci. 2019, 22, 38–42. [Google Scholar] [CrossRef]
- Al-Bayssari, C.; Diene, S.M.; Loucif, L.; Gupta, S.K.; Dabboussi, F.; Mallat, H.; Hamze, M.; Rolain, J.M. Emergence of VIM-2 and IMP-15 carbapenemases and inactivation of oprD gene in carbapenem-resistant Pseudomonas aeruginosa clinical isolates from Lebanon. Antimicrob. Agents Chemother. 2014, 58, 4966–4970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estepa, V.; Rojo-Bezares, B.; Azcona-Gutiérrez, J.M.; Olarte, I.; Torres, C.; Sáenz, Y. Characterisation of carbapenem-resistance mechanisms in clinical Pseudomonas aeruginosa isolates recovered in a Spanish hospital. Enferm. Infecc. Microbiol. Clin. 2017, 35, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Chen, S.S.; Hung, K.H.; Wu, H.M.; Hsueh, P.R.; Yan, J.J.; Wu, J.J. Overproduction of active efflux pump and variations of OprD dominate in imipenem-resistant Pseudomonas aeruginosa isolated from patients with bloodstream infections in Taiwan. BMC Microbiol. 2016, 16, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Kong, W.; Yang, W.; Chen, G.; Liang, H.; Zhang, Y. Multilocus sequence typing and variations in the oprD gene of Pseudomonas aeruginosa isolated from a hospital in China. Infect. Drug Resist. 2018, 11, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocampo-Sosa, A.A.; Cabot, G.; Rodríguez, C.; Roman, E.; Tubau, F.; Macia, M.D.; Moya, B.; Zamorano, L.; Suárez, C.; Peña, C.; et al. Alterations of OprD in carbapenem-intermediate and-susceptible strains of Pseudomonas aeruginosa isolated from patients with bacteremia in a Spanish multicenter study. Antimicrob. Agents Chemother. 2012, 56, 1703–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappa, O.; Kefala, A.M.; Tryfinopoulou, K.; Dimitriou, M.; Kostoulas, K.; Dioli, C.; Moraitou, E.; Panopoulou, M.; Vogiatzakis, E.; Mavridou, A.; et al. Molecular Epidemiology of Multi-Drug Resistant Pseudomonas aeruginosa Isolates from Hospitalized Patients in Greece. Microorganisms 2020, 8, 1652. https://doi.org/10.3390/microorganisms8111652
Pappa O, Kefala AM, Tryfinopoulou K, Dimitriou M, Kostoulas K, Dioli C, Moraitou E, Panopoulou M, Vogiatzakis E, Mavridou A, et al. Molecular Epidemiology of Multi-Drug Resistant Pseudomonas aeruginosa Isolates from Hospitalized Patients in Greece. Microorganisms. 2020; 8(11):1652. https://doi.org/10.3390/microorganisms8111652
Chicago/Turabian StylePappa, Olga, Anastasia Maria Kefala, Kyriaki Tryfinopoulou, Marios Dimitriou, Kostas Kostoulas, Chrysa Dioli, Eleni Moraitou, Maria Panopoulou, Evaggelos Vogiatzakis, Athena Mavridou, and et al. 2020. "Molecular Epidemiology of Multi-Drug Resistant Pseudomonas aeruginosa Isolates from Hospitalized Patients in Greece" Microorganisms 8, no. 11: 1652. https://doi.org/10.3390/microorganisms8111652