

Application of a Novel Proteomic Microarray Reveals High Exposure to Diarrhoeagenic Escherichia coli among Children in Zambia Participating in a Phase I Clinical Trial
 , , , , and
, , , , and 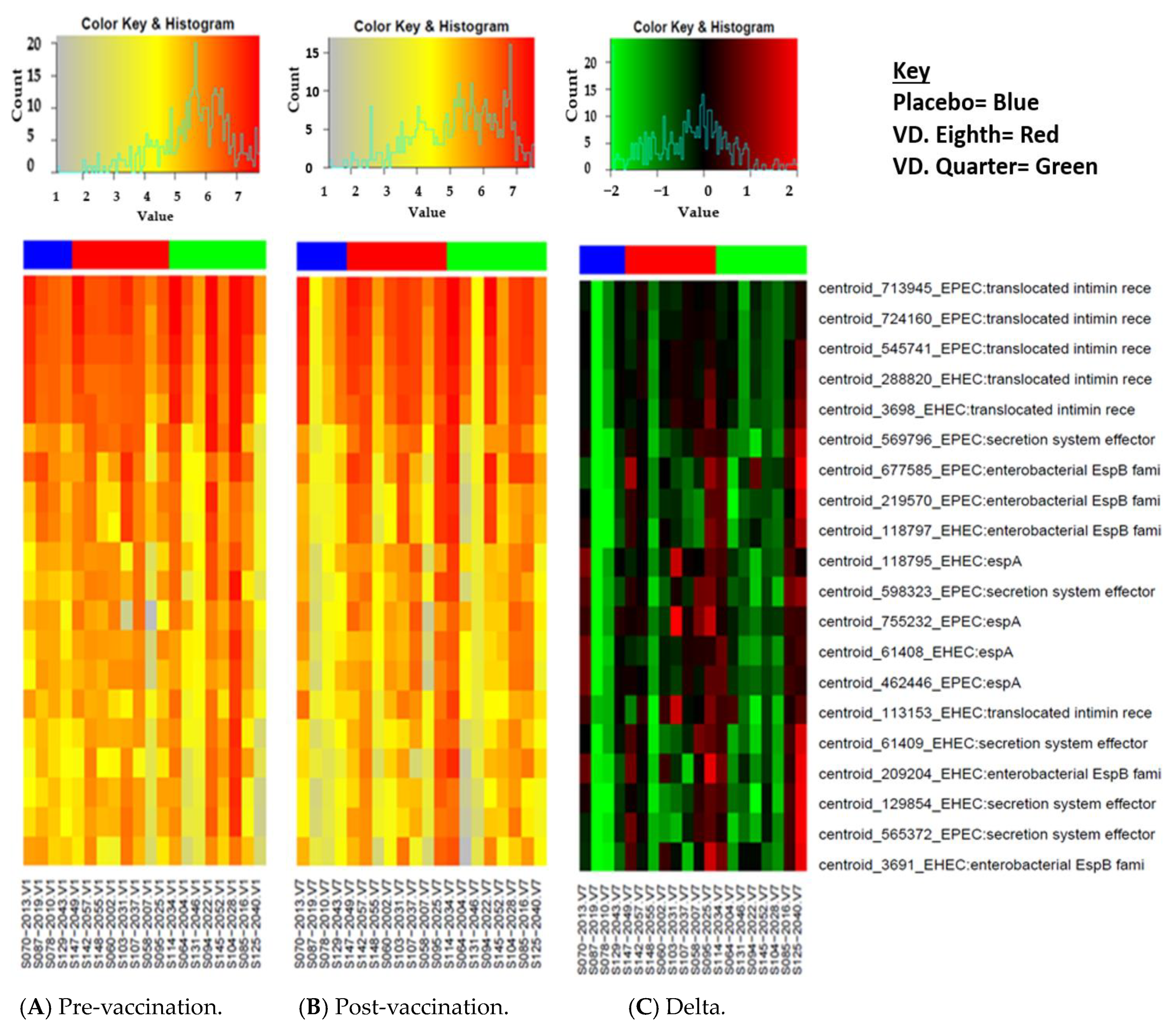{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Summary of the Parent Study and Selection of Samples for the Sub-Analysis
2.2. Ethical Considerations
2.3. Laboratory Analysis
Microarray Creation
2.4. Sample Analysis
2.5. Data Analysis
3. Results
3.1. Selection of Clinical Samples for the Microarray
3.2. Top 20 Reactive Proteins
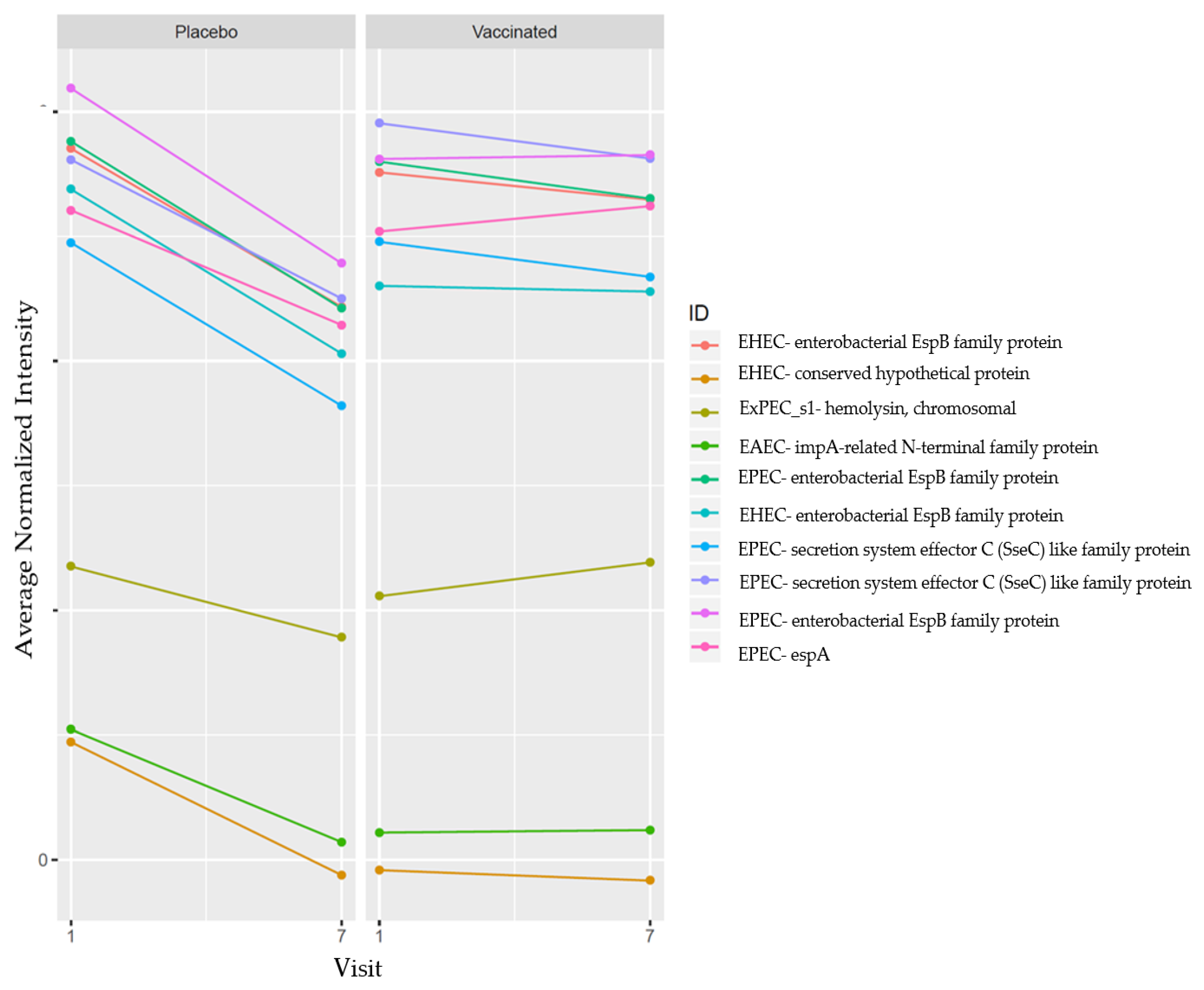
3.3. Top 10 Reactive Proteins by Mean Differences in the Vaccinated Group
3.4. Top 10 Reactive Proteins by Antibody Mean Differences in the Placebo Group
3.5. Delta Changes in the Vaccinated and Placebo Group between V1 and V7
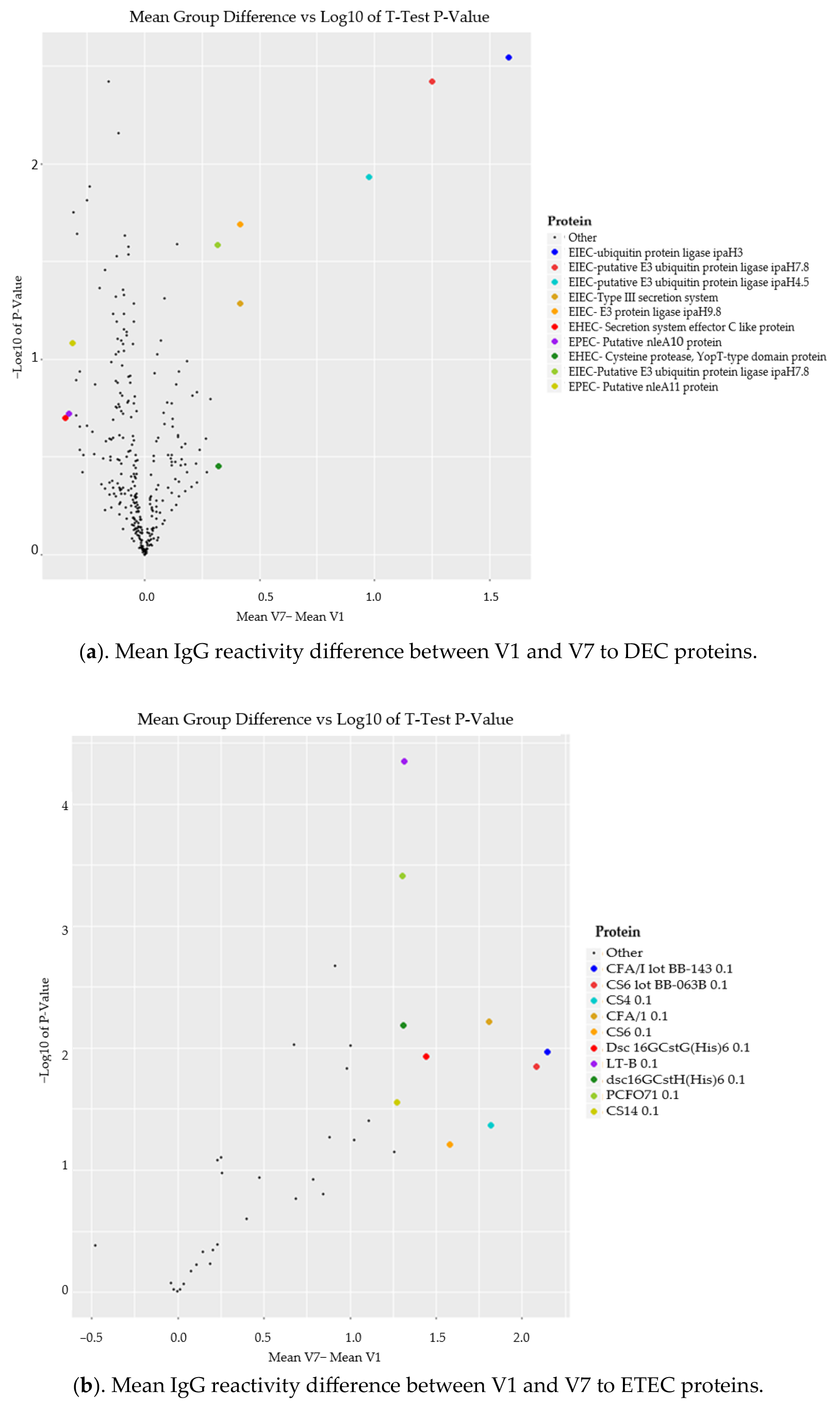
3.6. Comparison of Mean IgG Reactivity Differences (V7-V1) between DEC and ETEC Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, C.L.F.; Rudan, I.; Liu, L.; Nair, H.; Theodoratou, E.; Bhutta, Z.A.; O’Brien, K.L.; Campbell, H.; Black, R.E. Global Burden of Childhood Pneumonia and Diarrhoea. Lancet 2013, 381, 1405–1416. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and Aetiology of Diarrhoeal Disease in Infants and Young Children in Developing Countries (the Global Enteric Multicenter Study, GEMS): A Prospective, Case-Control Study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Pinkerton, R.; Oriá, R.B.; Lima, A.A.M.; Rogawski, E.T.; Oriá, M.O.B.; Patrick, P.D.; Moore, S.R.; Wiseman, B.L.; Niehaus, M.D.; Guerrant, R.L. Early Childhood Diarrhea Predicts Cognitive Delays in Later Childhood Independently of Malnutrition. Am. J. Trop. Med. Hyg. 2016, 95, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Osawa, K.; Raharjo, D.; Wasito, E.B.; Harijono, S.; Shigemura, K.; Osawa, R.; Sudarmo, S.M.; Iijima, Y.; Shirakawa, T. Frequency of Diarrheagenic Escherichia coli among Children in Surabaya, Indonesia. Jpn. J. Infect. Dis. 2013, 66, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Pérez, C.; Gómez-Duarte, O.G.; Arias, M.L. Diarrheagenic Escherichia coli in Children from Costa Rica. Am. J. Trop. Med. Hyg. 2010, 83, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.; Walker, R.; Porter, C.K.; Muhib, F.; Chilengi, R.; Cravioto, A.; Guerrant, R.; Svennerholm, A.-M.; Qadri, F.; Baqar, S.; et al. Enterotoxigenic Escherichia coli (ETEC) Vaccines: Priority Activities to Enable Product Development, Licensure, and Global Access. Vaccine 2021, 39, 4266–4277. [Google Scholar] [CrossRef]
- Geurtsen, J.; de Been, M.; Weerdenburg, E.; Zomer, A.; McNally, A.; Poolman, J. Genomics and Pathotypes of the Many Faces of Escherichia coli. FEMS Microbiol. Rev. 2022, 46, fuac031. [Google Scholar] [CrossRef]
- Mwape, K.; Bosomprah, S.; Chibesa, K.; Silwamba, S.; Luchen, C.C.; Sukwa, N.; Mubanga, C.; Phiri, B.; Chibuye, M.; Liswaniso, F.; et al. Prevalence of Diarrhoeagenic Escherichia coli among Children Aged between 0–36 Months in Peri-Urban Areas of Lusaka. Microorganisms 2023, 11, 2790. [Google Scholar] [CrossRef]
- Gomes, T.A.T.; Elias, W.P.; Scaletsky, I.C.A.; Guth, B.E.C.; Rodrigues, J.F.; Piazza, R.M.F.; Ferreira, L.C.S.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2016, 47 (Suppl. 1), 3–30. [Google Scholar] [CrossRef]
- Yu, F.; Chen, X.; Zheng, S.; Han, D.; Wang, Y.; Wang, R.; Wang, B.; Chen, Y. Prevalence and Genetic Diversity of Human Diarrheagenic Escherichia coli Isolates by Multilocus Sequence Typing. Int. J. Infect. Dis. 2018, 67, 7–13. [Google Scholar] [CrossRef]
- Thakur, N.; Jain, S.; Changotra, H.; Shrivastava, R.; Kumar, Y.; Grover, N.; Vashistt, J. Molecular Characterization of Diarrheagenic Escherichia coli Pathotypes: Association of Virulent Genes, Serogroups, and Antibiotic Resistance among Moderate-to-Severe Diarrhea Patients. J. Clin. Lab. Anal. 2018, 32, e22388. [Google Scholar] [CrossRef] [PubMed]
- Hazen, T.H.; Michalski, J.; Luo, Q.; Shetty, A.C.; Daugherty, S.C.; Fleckenstein, J.M.; Rasko, D.A. Comparative Genomics and Transcriptomics of Escherichia coli Isolates Carrying Virulence Factors of Both Enteropathogenic and Enterotoxigenic E. coli. Sci. Rep. 2017, 7, 3513. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.N.; Bourgeois, A.L.; Ericsson, C.D.; Steffen, R.; Jiang, Z.-D.; Halpern, J.; Haake, R.; Dupont, H.L. A Randomized, Double-Blind, Multicenter Study of Rifaximin Compared with Placebo and with Ciprofloxacin in the Treatment of Travelers’ Diarrhea. Am. J. Trop. Med. Hyg. 2006, 74, 1060–1066. [Google Scholar] [CrossRef]
- Okeke, I.N.; Aboderin, O.A.; Byarugaba, D.K.; Ojo, K.K.; Opintan, J.A. Growing Problem of Multidrug-Resistant Enteric Pathogens in Africa. Emerg. Infect. Dis. 2007, 13, 1640–1646. [Google Scholar] [CrossRef]
- Ingle, D.J.; Levine, M.M.; Kotloff, K.L.; Holt, K.E.; Robins-Browne, R.M. Dynamics of Antimicrobial Resistance in Intestinal Escherichia coli from Children in Community Settings in South Asia and Sub-Saharan Africa. Nat. Microbiol. 2018, 3, 1063–1073. [Google Scholar] [CrossRef]
- Msolo, L.; Iweriebor, B.C.; Okoh, A.I. Antimicrobial Resistance Profiles of Diarrheagenic E. coli (DEC) and Salmonella Species Recovered from Diarrheal Patients in Selected Rural Communities of the Amathole District Municipality, Eastern Cape Province, South Africa. Infect. Drug Resist. 2020, 13, 4615–4626. [Google Scholar] [CrossRef] [PubMed]
- Ohwo, O.; Agusomu, T.D. Assessment of Water, Sanitation and Hygiene Services in Sub-Saharan Africa. Eur. Sci. J. ESJ 2018, 14, 308. [Google Scholar] [CrossRef]
- Qadri, F.; Akhtar, M.; Bhuiyan, T.R.; Chowdhury, M.I.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.A.; Khanam, F.; Lundgren, A.; et al. Safety and Immunogenicity of the Oral, Inactivated, Enterotoxigenic Escherichia coli Vaccine ETVAX in Bangladeshi Children and Infants: A Double-Blind, Randomised, Placebo-Controlled Phase 1/2 Trial. Lancet Infect. Dis. 2020, 20, 208–219. [Google Scholar] [CrossRef]
- Darsley, M.J.; Chakraborty, S.; DeNearing, B.; Sack, D.A.; Feller, A.; Buchwaldt, C.; Bourgeois, A.L.; Walker, R.; Harro, C.D. The Oral, Live Attenuated Enterotoxigenic Escherichia coli Vaccine ACE527 Reduces the Incidence and Severity of Diarrhea in a Human Challenge Model of Diarrheal Disease. Clin. Vaccine Immunol. CVI 2012, 19, 1921–1931. [Google Scholar] [CrossRef]
- Riddle, M.S.; Maciel, M.J.; Porter, C.K.; Poole, S.T.; Gutierrez, R.L.; Gormley, R.; Laird, R.M.; Sebeny, P.J.; Dori, K.E.; Greenleaf, M.E.; et al. A First in Human Clinical Trial Assessing the Safety and Immunogenicity of Transcutaneously Delivered Enterotoxigenic Escherichia coli Fimbrial Tip Adhesin with Heat-Labile Enterotoxin with Mutation R192G. Vaccine 2020, 38, 7040–7048. [Google Scholar] [CrossRef]
- Fleckenstein, J.M. Confronting Challenges to Enterotoxigenic Escherichia coli Vaccine Development. Front. Trop. Dis. 2021, 2, 709907. [Google Scholar] [CrossRef]
- Holmgren, J.; Bourgeois, L.; Carlin, N.; Clements, J.; Gustafsson, B.; Lundgren, A.; Nygren, E.; Tobias, J.; Walker, R.; Svennerholm, A.-M. Development and Preclinical Evaluation of Safety and Immunogenicity of an Oral ETEC Vaccine Containing Inactivated E. coli Bacteria Overexpressing Colonization Factors CFA/I, CS3, CS5 and CS6 Combined with a Hybrid LT/CT B Subunit Antigen, Administered Alone and Together with dmLT Adjuvant. Vaccine 2013, 31, 2457–2464. [Google Scholar] [CrossRef] [PubMed]
- Vojtek, I.; Buchy, P.; Doherty, T.M.; Hoet, B. Would Immunization Be the Same without Cross-Reactivity? Vaccine 2019, 37, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, S.; Khan, R.; Ferreira, D.M.; Mitsi, E.; German, E.; Rørvik, G.H.; Berild, D.; Schenck, K.; Kwon, K.; Petersen, F. Antibodies Reactive to Commensal Streptococcus mitis Show Cross-Reactivity with Virulent Streptococcus pneumoniae Serotypes. Front. Immunol. 2018, 9, 747. [Google Scholar] [CrossRef] [PubMed]
- Ndungo, E.; Randall, A.; Hazen, T.H.; Kania, D.A.; Trappl-Kimmons, K.; Liang, X.; Barry, E.M.; Kotloff, K.L.; Chakraborty, S.; Mani, S.; et al. A Novel Shigella Proteome Microarray Discriminates Targets of Human Antibody Reactivity Following Oral Vaccination and Experimental Challenge. mSphere 2018, 3, e00260-18. [Google Scholar] [CrossRef]
- Mubanga, C.; Simuyandi, M.; Mwape, K.; Chibesa, K.; Chisenga, C.; Chilyabanyama, O.N.; Randall, A.; Liang, X.; Glashoff, R.H.; Chilengi, R. Use of an ETEC Proteome Microarray to Evaluate Cross-Reactivity of ETVAX® Vaccine-Induced IgG Antibodies in Zambian Children. Vaccines 2023, 11, 939. [Google Scholar] [CrossRef]
- Prechl, J.; Papp, K.; Erdei, A. Antigen Microarrays: Descriptive Chemistry or Functional Immunomics? Trends Immunol. 2010, 31, 133–137. [Google Scholar] [CrossRef]
- Sukwa, N.; Mubanga, C.; Hatyoka, L.M.; Chilyabanyama, O.N.; Chibuye, M.; Mundia, S.; Munyinda, M.; Kamuti, E.; Siyambango, M.; Badiozzaman, S.; et al. Safety, Tolerability, and Immunogenicity of an Oral Inactivated ETEC Vaccine (ETVAX®) with dmLT Adjuvant in Healthy Adults and Children in Zambia: An Age Descending Randomised, Placebo-Controlled Trial. Vaccine 2023, 41, 6884–6894. [Google Scholar] [CrossRef]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L.; et al. Human Experimental Challenge with Enterotoxigenic Escherichia coli Elicits Immune Responses to Canonical and Novel Antigens Relevant to Vaccine Development. J. Infect. Dis. 2018, 218, 1436–1446. [Google Scholar] [CrossRef]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L.; et al. Interrogation of a Live-Attenuated Enterotoxigenic Escherichia coli Vaccine Highlights Features Unique to Wild-Type Infection. NPJ Vaccines 2019, 4, 37. [Google Scholar] [CrossRef]
- Mühlen, S.; Zapol’skii, V.A.; Bilitewski, U.; Dersch, P. Identification of Translocation Inhibitors Targeting the Type III Secretion System of Enteropathogenic Escherichia coli. Antimicrob. Agents Chemother. 2021, 65, e0095821. [Google Scholar] [CrossRef] [PubMed]
- Kordbacheh, E.; Nazarian, S.; Hajizadeh, A.; Fasihi-Ramandi, M.; Fathi, J. Recombinant HcpA-EspA-Tir-Stx2B Chimeric Protein Induces Immunity against Attachment and Toxicity of Escherichia coli O157:H7. Microb. Pathog. 2019, 129, 176–182. [Google Scholar] [CrossRef]
- Nyambe, S.; Agestika, L.; Yamauchi, T. The Improved and the Unimproved: Factors Influencing Sanitation and Diarrhoea in a Peri-Urban Settlement of Lusaka, Zambia. PLoS ONE 2020, 15, e0232763. [Google Scholar] [CrossRef]
- Giugliano, L.G.; Ribeiro, S.T.; Vainstein, M.H.; Ulhoa, C.J. Free Secretory Component and Lactoferrin of Human Milk Inhibit the Adhesion of Enterotoxigenic Escherichia coli. J. Med. Microbiol. 1995, 42, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. The Mucosal Immune System and Its Integration with the Mammary Glands. J. Pediatr. 2010, 156, S8–S15. [Google Scholar] [CrossRef]
- Miti, S.; Chilyabanyama, O.N.; Chisenga, C.C.; Chibuye, M.; Bosomprah, S.; Mumba, C.; Chitondo, S.; Siziya, S.; Cohen, D.; Chilengi, R.; et al. Sensitivity and Predictive Value of Dysentery in Diagnosing Shigellosis among under Five Children in Zambia. PLoS ONE 2023, 18, e0279012. [Google Scholar] [CrossRef]
- Calderon Toledo, C.; Arvidsson, I.; Karpman, D. Cross-Reactive Protection against Enterohemorrhagic Escherichia coli Infection by Enteropathogenic E. coli in a Mouse Model. Infect. Immun. 2011, 79, 2224–2233. [Google Scholar] [CrossRef]
- Yang, S.-C.; Hung, C.-F.; Aljuffali, I.A.; Fang, J.-Y. The Roles of the Virulence Factor IpaB in Shigella spp. in the Escape from Immune Cells and Invasion of Epithelial Cells. Microbiol. Res. 2015, 181, 43–51. [Google Scholar] [CrossRef]
- Gaytán, M.O.; Martínez-Santos, V.I.; Soto, E.; González-Pedrajo, B. Type Three Secretion System in Attaching and Effacing Pathogens. Front. Cell. Infect. Microbiol. 2016, 6, 129. [Google Scholar] [CrossRef]
- Thanabalasuriar, A.; Koutsouris, A.; Weflen, A.; Mimee, M.; Hecht, G.; Gruenheid, S. The Bacterial Virulence Factor NleA Is Required for the Disruption of Intestinal Tight Junctions by Enteropathogenic Escherichia coli. Cell. Microbiol. 2010, 12, 31–41. [Google Scholar] [CrossRef]
- Baumann, D.; Salia, H.; Greune, L.; Norkowski, S.; Körner, B.; Uckeley, Z.M.; Frankel, G.; Guenot, M.; Rüter, C.; Schmidt, M.A. Multitalented EspB of Enteropathogenic Escherichia coli (EPEC) Enters Cells Autonomously and Induces Programmed Cell Death in Human Monocytic THP-1 Cells. Int. J. Med. Microbiol. IJMM 2018, 308, 387–404. [Google Scholar] [CrossRef]
- Li, D.; Chen, Z.; Cheng, H.; Zheng, J.-X.; Pan, W.-G.; Yang, W.-Z.; Yu, Z.-J.; Deng, Q.-W. Inhibition of Adhesion of Enteropathogenic Escherichia coli to HEp-2 Cells by Binding of a Novel Peptide to EspB Protein. Curr. Microbiol. 2016, 73, 361–365. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwape, K.; Mubanga, C.; Chilyabanyama, O.N.; Chibesa, K.; Chisenga, C.C.; Silwamba, S.; Randall, A.; Liang, X.; Barnard, T.G.; Simuyandi, M.; et al. Application of a Novel Proteomic Microarray Reveals High Exposure to Diarrhoeagenic Escherichia coli among Children in Zambia Participating in a Phase I Clinical Trial. Microorganisms 2024, 12, 420. https://doi.org/10.3390/microorganisms12030420
Mwape K, Mubanga C, Chilyabanyama ON, Chibesa K, Chisenga CC, Silwamba S, Randall A, Liang X, Barnard TG, Simuyandi M, et al. Application of a Novel Proteomic Microarray Reveals High Exposure to Diarrhoeagenic Escherichia coli among Children in Zambia Participating in a Phase I Clinical Trial. Microorganisms. 2024; 12(3):420. https://doi.org/10.3390/microorganisms12030420
Chicago/Turabian StyleMwape, Kapambwe, Cynthia Mubanga, Obvious Nchimunya Chilyabanyama, Kennedy Chibesa, Caroline Cleopatra Chisenga, Suwilanji Silwamba, Arlo Randall, Xiaowu Liang, Tobias George Barnard, Michelo Simuyandi, and et al. 2024. "Application of a Novel Proteomic Microarray Reveals High Exposure to Diarrhoeagenic Escherichia coli among Children in Zambia Participating in a Phase I Clinical Trial" Microorganisms 12, no. 3: 420. https://doi.org/10.3390/microorganisms12030420