
Human Milk-Derived Enterococcus faecalis HM20: A Potential Alternative Agent of Antimicrobial Effect against Methicillin-Resistant Staphylococcus aureus (MRSA)
 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Bacteria
2.2. Bacterial Growth Conditions and Metabolite Preparation
2.3. Agar-Well Diffusion Assay
2.4. Broth Microdilution Assay
2.5. Crystal Violet Assay for Biofilm Formation
2.6. Characterization of Antimicrobial Substances
2.7. Sodium Dodecyl Sulfate (SDS)–Polyacrylamide Gel Electrophoresis (PAGE) Antibacterial Assay
2.8. Scanning Electron Microscope (SEM)
2.9. Statistical Analysis
3. Results
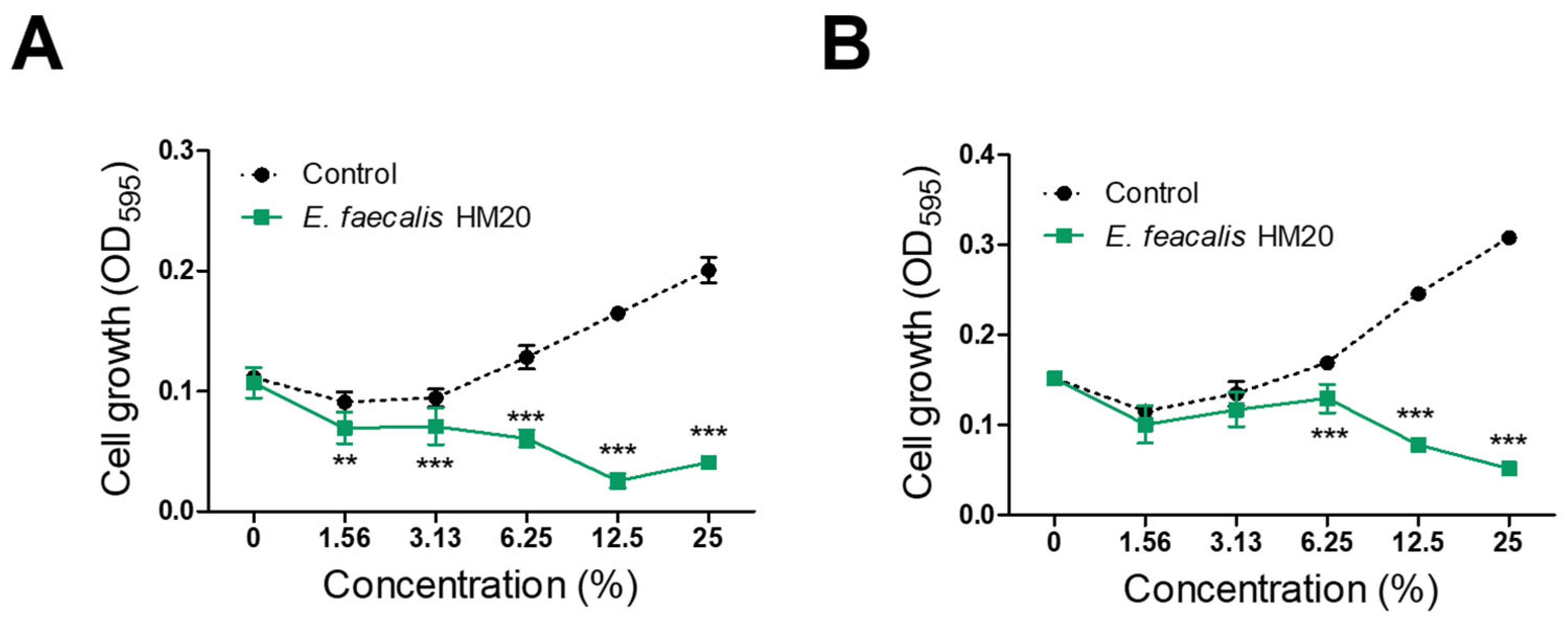
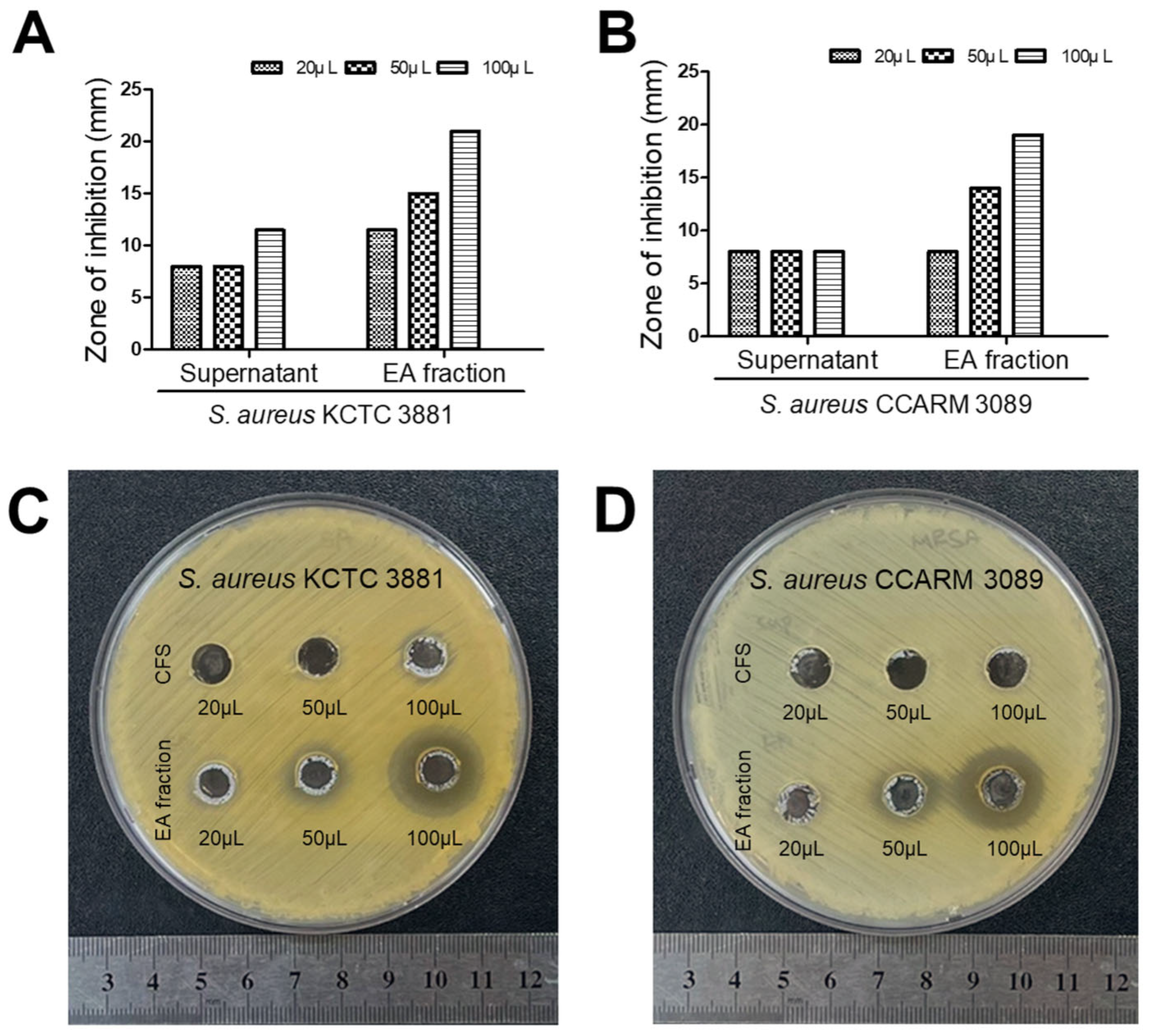
3.1. Antibacterial Effect of Isolated LAB
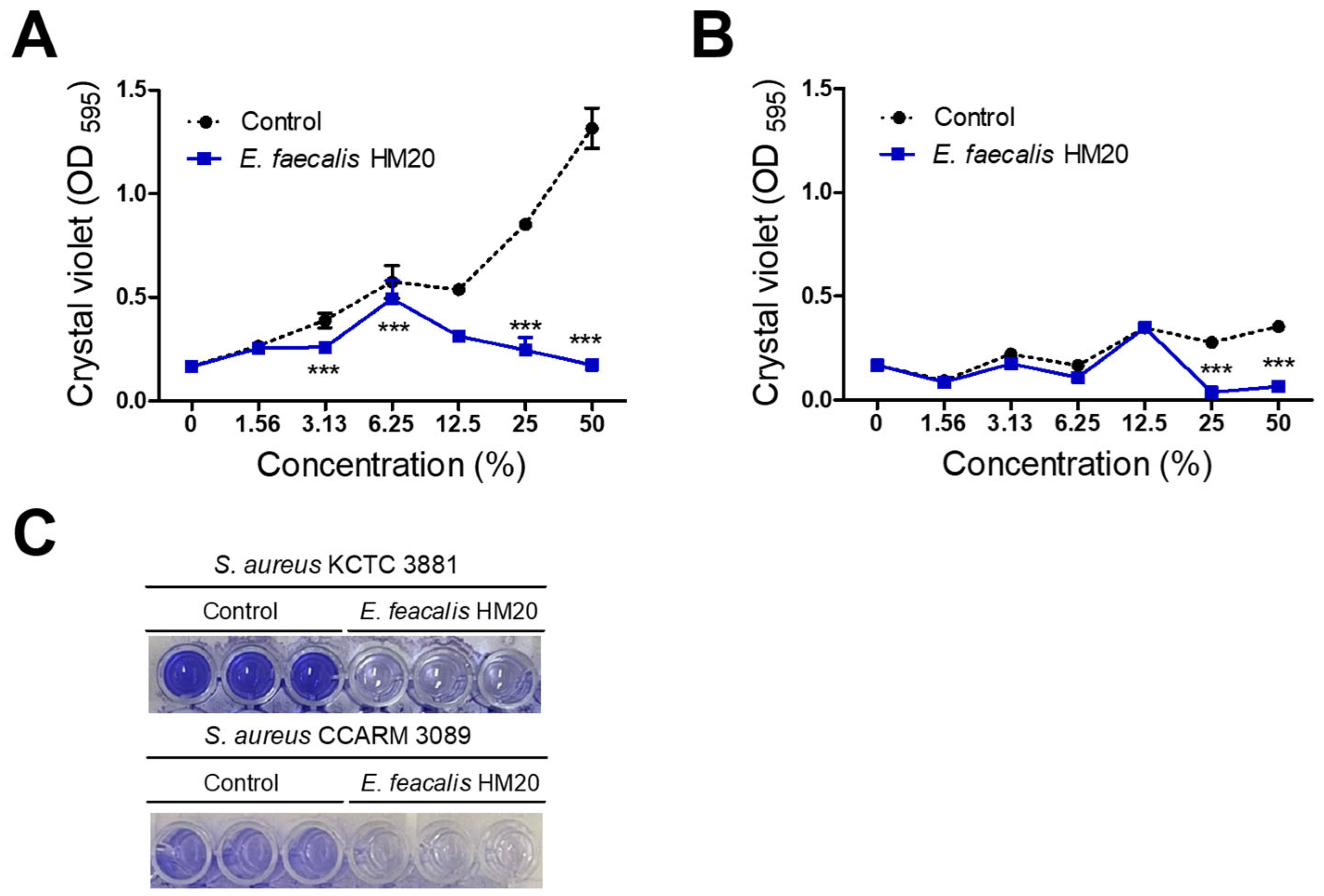
3.2. Inhibition of Biofilm Formation by E. faecalis HM20
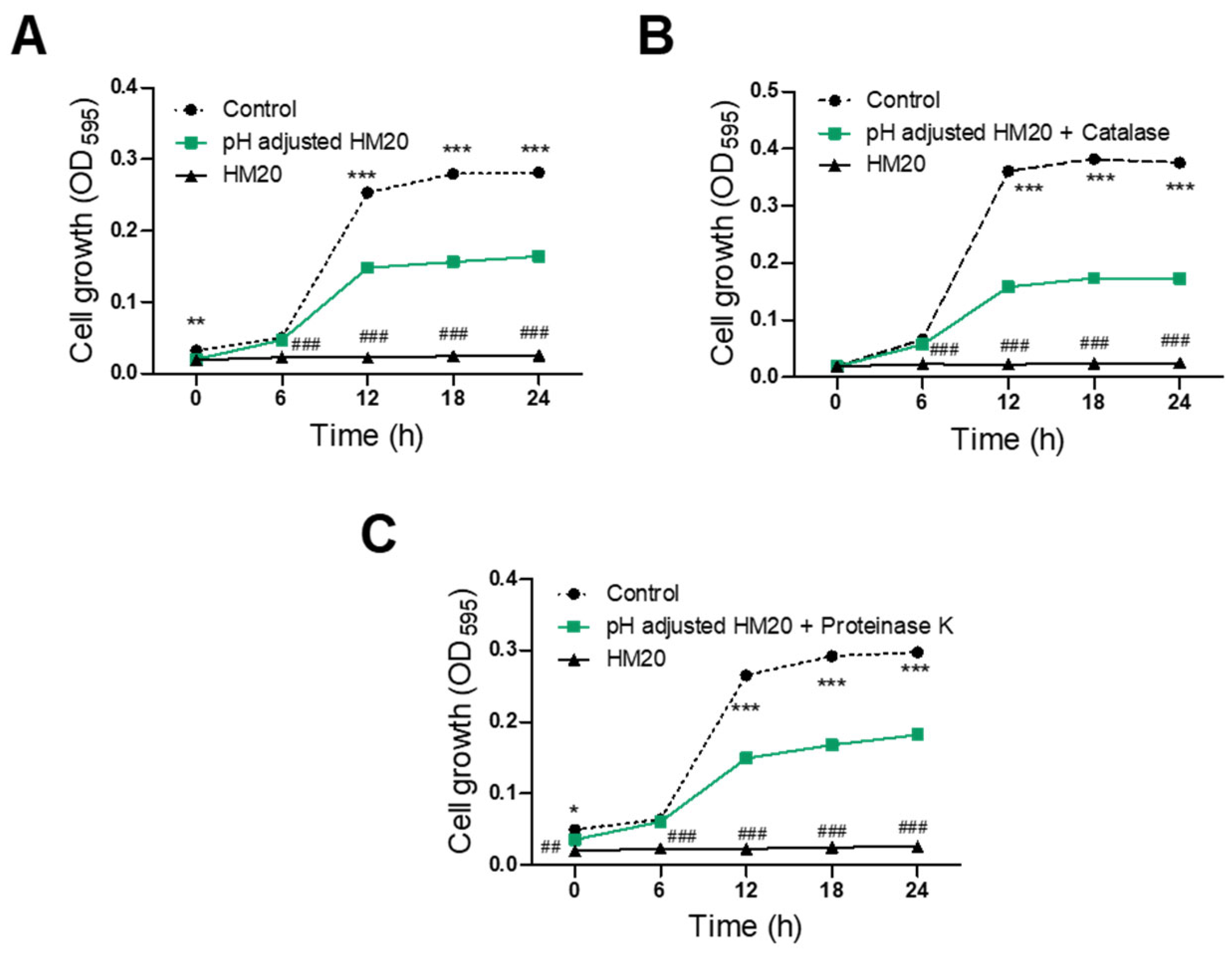
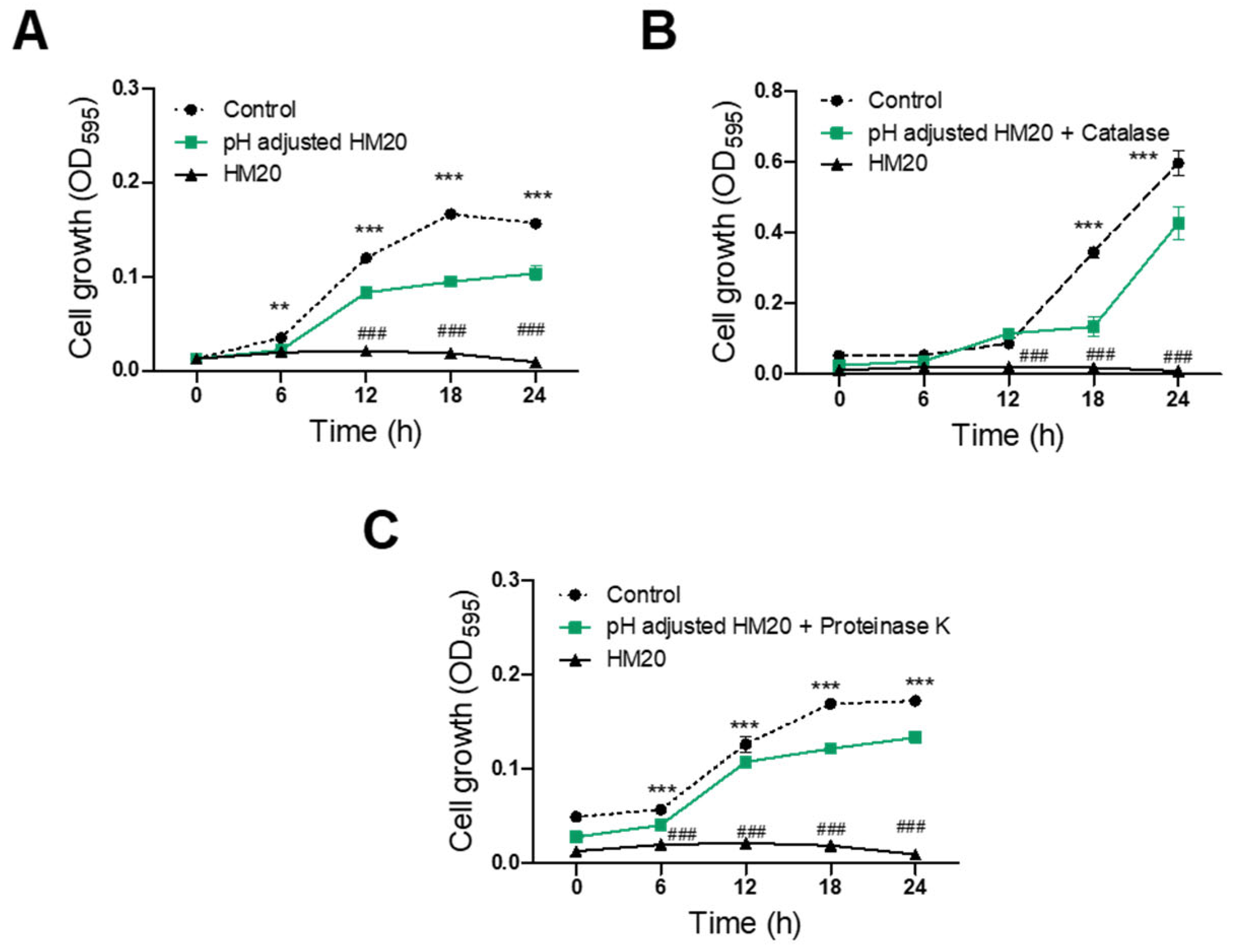
3.3. Characterization of Antimicrobial Substances of HM20
3.4. HM20 Antibacterial Effect against MRSA on Agar-Well Diffusion after Purification
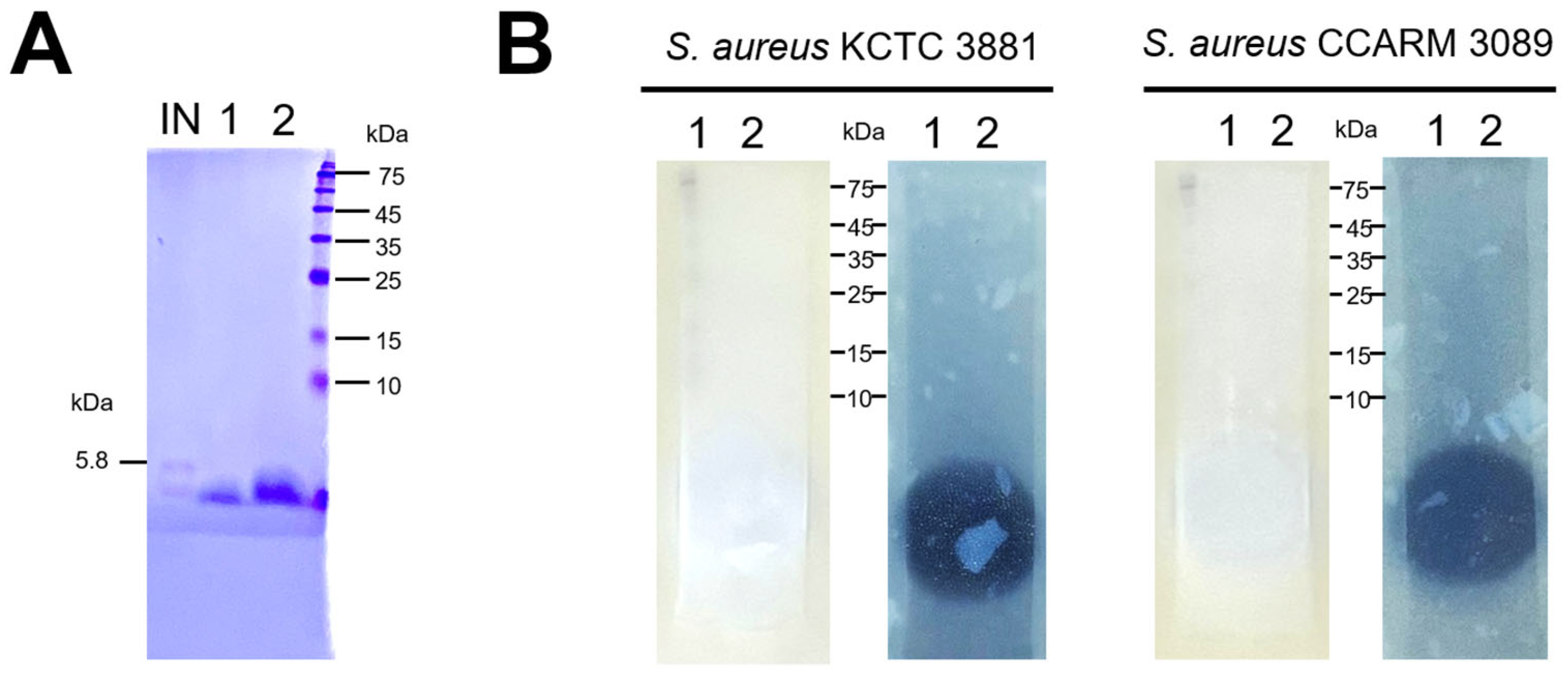
3.5. Analysis of Active Bacteriocins in E. faecalis HM20 Using SDS-PAGE and Overlaid Agar
3.6. SEM Result of Pathogens with E. faecalis HM20 Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ratnaraja, N.V.; Hawkey, P.M. Current Challenges in Treating MRSA: What Are the Options? Expert. Rev. Anti-Infect. Ther. 2008, 6, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.; Torres, V.J. Staphylococcus aureus Secreted Toxins and Extracellular Enzymes. Microbiol. Spectr. 2019, 7, 1–59. [Google Scholar] [CrossRef]
- De Jong, N.W.M.; Van Kessel, K.P.M.; Van Strijp, J.A.G. Immune Evasion by Staphylococcus aureus. Microbiol. Spectr. 2019, 7, 1–27. [Google Scholar] [CrossRef]
- Peate, I. The Skin: Largest Organ of the Body. Br. J. Healthc. Assist. 2021, 15, 446–451. [Google Scholar] [CrossRef]
- Eells, S.J.; David, M.Z.; Taylor, A.; Ortiz, N.; Kumar, N.; Sieth, J.; Boyle-Vavra, S.; Daum, R.S.; Miller, L.G. Persistent Environmental Contamination with USA300 Methicillin-Resistant Staphylococcus aureus and Other Pathogenic Strain Types in Households with S. aureus Skin Infections. Infect. Control Hosp. Epidemiol. 2014, 35, 1373–1382. [Google Scholar] [CrossRef]
- Azarian, T.; Daum, R.S.; Petty, L.A.; Steinbeck, J.L.; Yin, Z.; Nolan, D.; Boyle-Vavra, S.; Hanage, W.P.; Salemi, M.; David, M.Z. Intrahost Evolution of Methicillin-Resistant Staphylococcus aureus USA300 Among Individuals with Reoccurring Skin and Soft-Tissue Infections. J. Infect. Dis. 2016, 214, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; DeLeo, F.R. Mobile Genetic Elements of Staphylococcus aureus. Cell. Mol. Life Sci. 2010, 67, 3057–3071. [Google Scholar] [CrossRef]
- Deurenberg, R.H.; Vink, C.; Kalenic, S.; Friedrich, A.W.; Bruggeman, C.A.; Stobberingh, E.E. The Molecular Evolution of Methicillin-Resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2007, 13, 222–235. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Lodise, T.P.; Graves, J.; Evans, A.; Graffunder, E.; Helmecke, M.; Lomaestro, B.M.; Stellrecht, K. Relationship between Vancomycin MIC and Failure among Patients with Methicillin-Resistant Staphylococcus aureus Bacteremia Treated with Vancomycin. Antimicrob. Agents Chemother. 2008, 52, 3315–3320. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
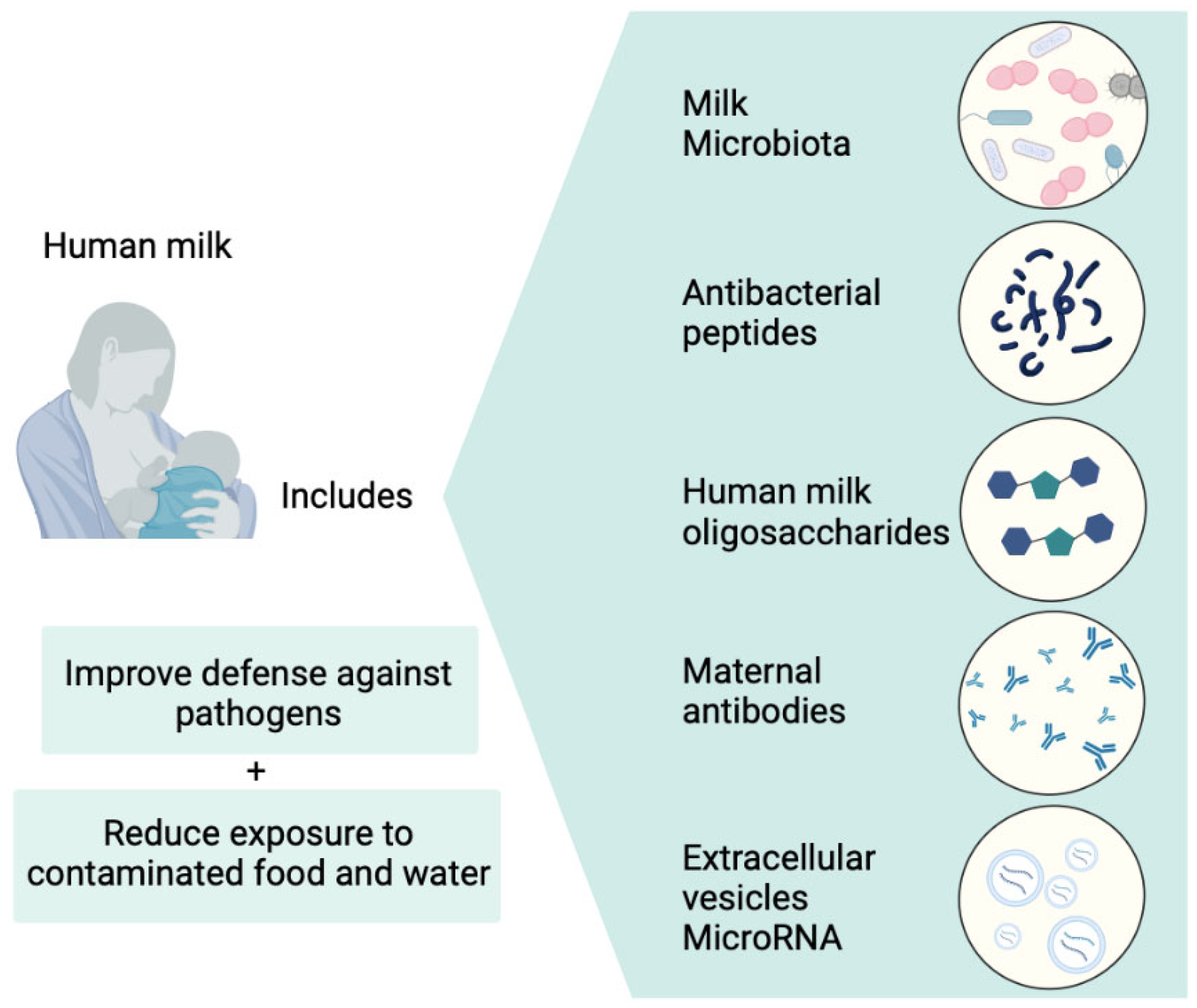
- Donnet-Hughes, A.; Perez, P.F.; Doré, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J. Potential Role of the Intestinal Microbiota of the Mother in Neonatal Immune Education. Proc. Nutr. Soc. 2010, 69, 407–415. [Google Scholar] [CrossRef]
- Fernández, L.; Langa, S.; Martín, V.; Maldonado, A.; Jiménez, E.; Martín, R.; Rodríguez, J.M. The Human Milk Microbiota: Origin and Potential Roles in Health and Disease. Pharmacol. Res. 2013, 69, 1–10. [Google Scholar] [CrossRef]
- LaTuga, M.; Stuebe, A.; Seed, P. A Review of the Source and Function of Microbiota in Breast Milk. Semin. Reprod. Med. 2014, 32, 068–073. [Google Scholar] [CrossRef]
- Lyons, K.E.; Ryan, C.A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Breast Milk, a Source of Beneficial Microbes and Associated Benefits for Infant Health. Nutrients 2020, 12, 1039. [Google Scholar] [CrossRef]
- Sakwinska, O.; Moine, D.; Delley, M.; Combremont, S.; Rezzonico, E.; Descombes, P.; Vinyes-Pares, G.; Zhang, Y.; Wang, P.; Thakkar, S.K. Microbiota in Breast Milk of Chinese Lactating Mothers. PLoS ONE 2016, 11, e0160856. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, A.; Asadi, A.; Mahdizade Ari, M.; Ohadi, E.; Talebi, M.; Halaj Zadeh, M.; Darb Emamie, A.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and Potential Use as Antimicrobials. Clin. Lab. Anal. 2022, 36, e24093. [Google Scholar] [CrossRef]
- García-Cano, I.; Serrano-Maldonado, C.E.; Olvera-García, M.; Delgado-Arciniega, E.; Peña-Montes, C.; Mendoza-Hernández, G.; Quirasco, M. Antibacterial Activity Produced by Enterococcus Spp. Isolated from an Artisanal Mexican Dairy Product, Cotija Cheese. LWT-Food Sci. Technol. 2014, 59, 26–34. [Google Scholar] [CrossRef]
- Promrug, D.; Wittayacom, K.; Nathapanan, N.; Dong, H.T.; Thongyoo, P.; Unajak, S.; Reamtong, O.; Boonyuen, U.; Aroonnual, A.; Shioda, T.; et al. Cocultures of Enterococcus faecium and Aeromonas veronii Induce the Secretion of Bacteriocin-like Substances against Aeromonas. J. Agric. Food Chem. 2023, 71, 16194–16203. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, Antimicrobial Peptides from Bacterial Origin: Overview of Their Biology and Their Impact against Multidrug-Resistant Bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Tagg, J.R.; McGiven, A.R. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef]
- Ruiz, M.J.; García, M.D.; Canalejo, L.M.M.; Krüger, A.; Padola, N.L.; Etcheverría, A.I. Antimicrobial Activity of Lactiplantibacillus plantarum against Shiga Toxin-Producing Escherichia coli. J. Appl. Microbiol. 2023, 134, lxad202. [Google Scholar] [CrossRef]
- Krause, R.G.E.; Goldring, J.P.D. Crystal Violet Stains Proteins in SDS-PAGE Gels and Zymograms. Anal. Biochem. 2019, 566, 107–115. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Togawa, Y.; Shimosaka, M.; Okazaki, M. Purification and Characterization of a Novel Bacteriocin Produced by Enterococcus faecalis Strain RJ-11. Appl. Environ. Microbiol. 2003, 69, 5746–5753. [Google Scholar] [CrossRef]
- Chinemerem Nwobodo, D.; Ugwu, M.C.; Oliseloke Anie, C.; Al-Ouqaili, M.T.S.; Chinedu Ikem, J.; Victor Chigozie, U.; Saki, M. Antibiotic Resistance: The Challenges and Some Emerging Strategies for Tackling a Global Menace. Clin. Lab. Anal. 2022, 36, e24655. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.-H.; Lee, J.-H.; Chang, S.-J.; Chae, Y.; Lee, M.-H.; Kim, S.-H.; Han, K.-I.; Kim, T.-J. Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice. Int. J. Mol. Sci. 2022, 23, 5413. [Google Scholar] [CrossRef]
- Choi, E.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Tang, Y.; Hwang, Y.; Chae, J.; Han, W.; Shin, Y.-S.; Kim, E.-K. Heat-Killed Enterococcus faecalis EF-2001 Ameliorates Atopic Dermatitis in a Murine Model. Nutrients 2016, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-S.; Chang, S.-J.; Chae, Y.; Lee, M.-H.; Kim, W.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Kim, T.-J. Anti-Inflammatory Effect of Heat-Killed Enterococcus faecalis, EF-2001. J. Life Sci. 2018, 28, 1361–1368. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Khan, J.; Tarar, S.M.; Gul, I.; Nawaz, U.; Arshad, M. Challenges of Antibiotic Resistance Biofilms and Potential Combating Strategies: A Review. 3 Biotech. 2021, 11, 169. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Bubonja-Šonje, M.; Knežević, S.; Abram, M. Challenges to Antimicrobial Susceptibility Testing of Plant-Derived Polyphenolic Compounds. Arch. Ind. Hyg. Toxicol. 2020, 71, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhen, Z.; Wang, X.; Guo, N. Synergy of a Combination of Nisin and Citric Acid against Staphylococcus aureus and Listeria monocytogenes. Food Addit. Contam. Part A 2017, 34, 2058–2068. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; Maris, P. Synergism between Hydrogen Peroxide and Seventeen Acids against Six Bacterial Strains. J. Appl. Microbiol. 2012, 113, 578–590. [Google Scholar] [CrossRef]
- Javed, I.; Ahmed, S. Production and Characterization of Enterocin from Enterococcus faecium IJ-06 and IJ-21 Isolated from Indigenous Dairy Products. In Science and Technology Against Microbial Pathogens: Research, Development and Evaluation; World Scientific: Valladolid, Spain, 2011; pp. 411–415. [Google Scholar]
- Tateda, K.; Ishii, Y.; Matsumoto, T.; Kobayashi, T.; Miyazaki, S.; Yamaguchi, K. Potential of Macrolide Antibiotics to Inhibit Protein Synthesis of Pseudomonas aeruginosa: Suppression of Virulence Factors and Stress Response. J. Infect. Chemother. 2000, 6, 1–7. [Google Scholar] [CrossRef]
- Wu, Y.; Pang, X.; Wu, Y.; Liu, X.; Zhang, X. Enterocins: Classification, Synthesis, Antibacterial Mechanisms and Food Applications. Molecules 2022, 27, 2258. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, E.-J.; Nguyen, T.T.M.; Jin, X.; Bellere, A.D.; Kim, M.-J.; Yi, T.-H. Human Milk-Derived Enterococcus faecalis HM20: A Potential Alternative Agent of Antimicrobial Effect against Methicillin-Resistant Staphylococcus aureus (MRSA). Microorganisms 2024, 12, 306. https://doi.org/10.3390/microorganisms12020306
Yi E-J, Nguyen TTM, Jin X, Bellere AD, Kim M-J, Yi T-H. Human Milk-Derived Enterococcus faecalis HM20: A Potential Alternative Agent of Antimicrobial Effect against Methicillin-Resistant Staphylococcus aureus (MRSA). Microorganisms. 2024; 12(2):306. https://doi.org/10.3390/microorganisms12020306
Chicago/Turabian StyleYi, Eun-Ji, Trang Thi Minh Nguyen, Xiangji Jin, Arce Defeo Bellere, Mi-Ju Kim, and Tae-Hoo Yi. 2024. "Human Milk-Derived Enterococcus faecalis HM20: A Potential Alternative Agent of Antimicrobial Effect against Methicillin-Resistant Staphylococcus aureus (MRSA)" Microorganisms 12, no. 2: 306. https://doi.org/10.3390/microorganisms12020306