
Differences in Bacterial Communities and Pathogen Indicators of Raw and Lagoon-Stabilized Farm Dairy Effluents


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dairy Effluent Collection and Farm Characteristics
2.2. Characterization of Dairy Effluents
2.3. Quantification of Pathogenic Bacterial Indicators
2.4. Dairy Effluents DNA Extraction and 16S rRNA qPCR and Gene Amplicon Sequencing
2.5. Statistical Analysis
3. Results
3.1. Farm Dairy Effluents Physicochemical Characterization
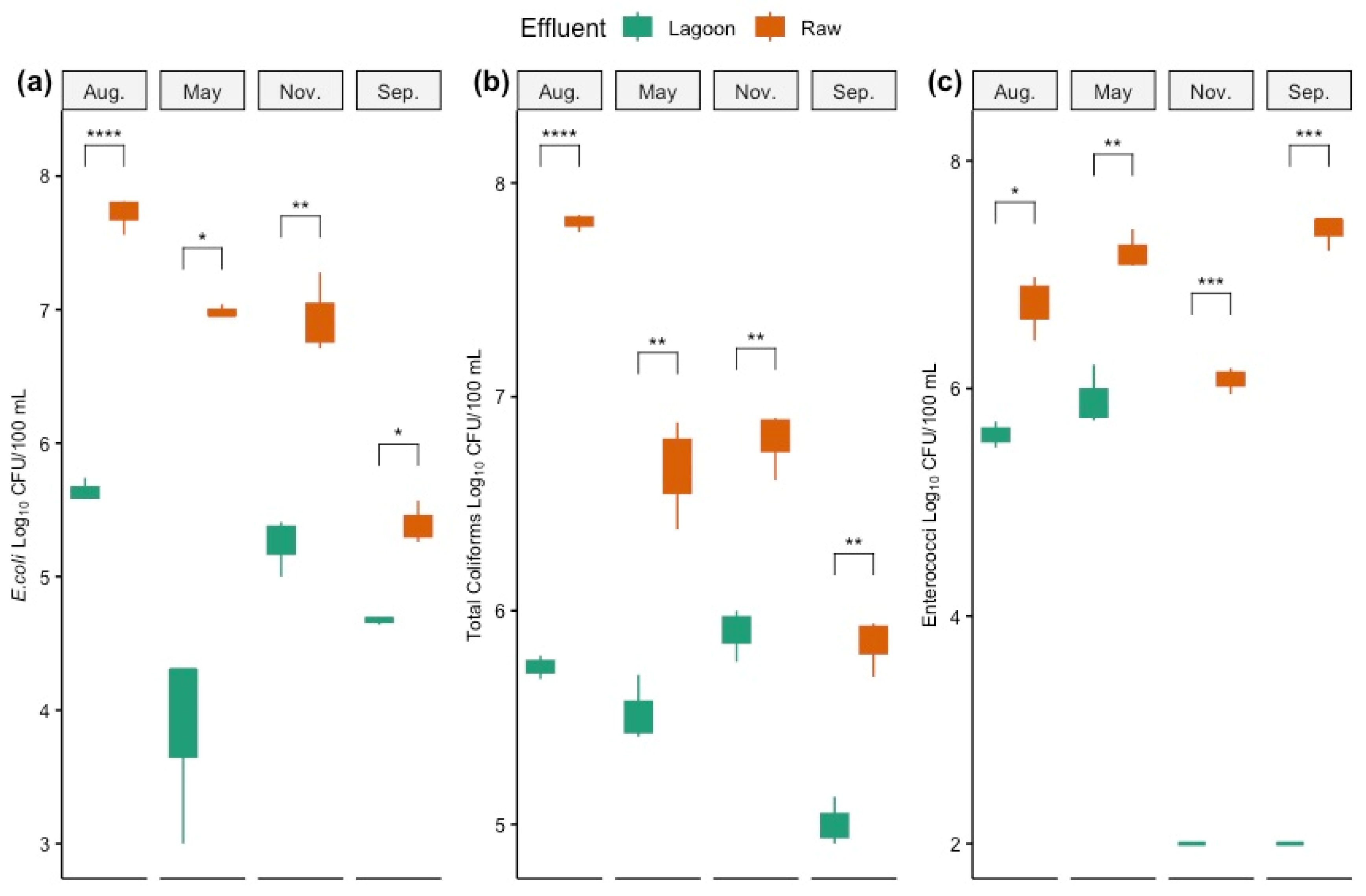
3.2. Quantification of Pathogenic Indicator Bacteria in Farm Dairy Effluents
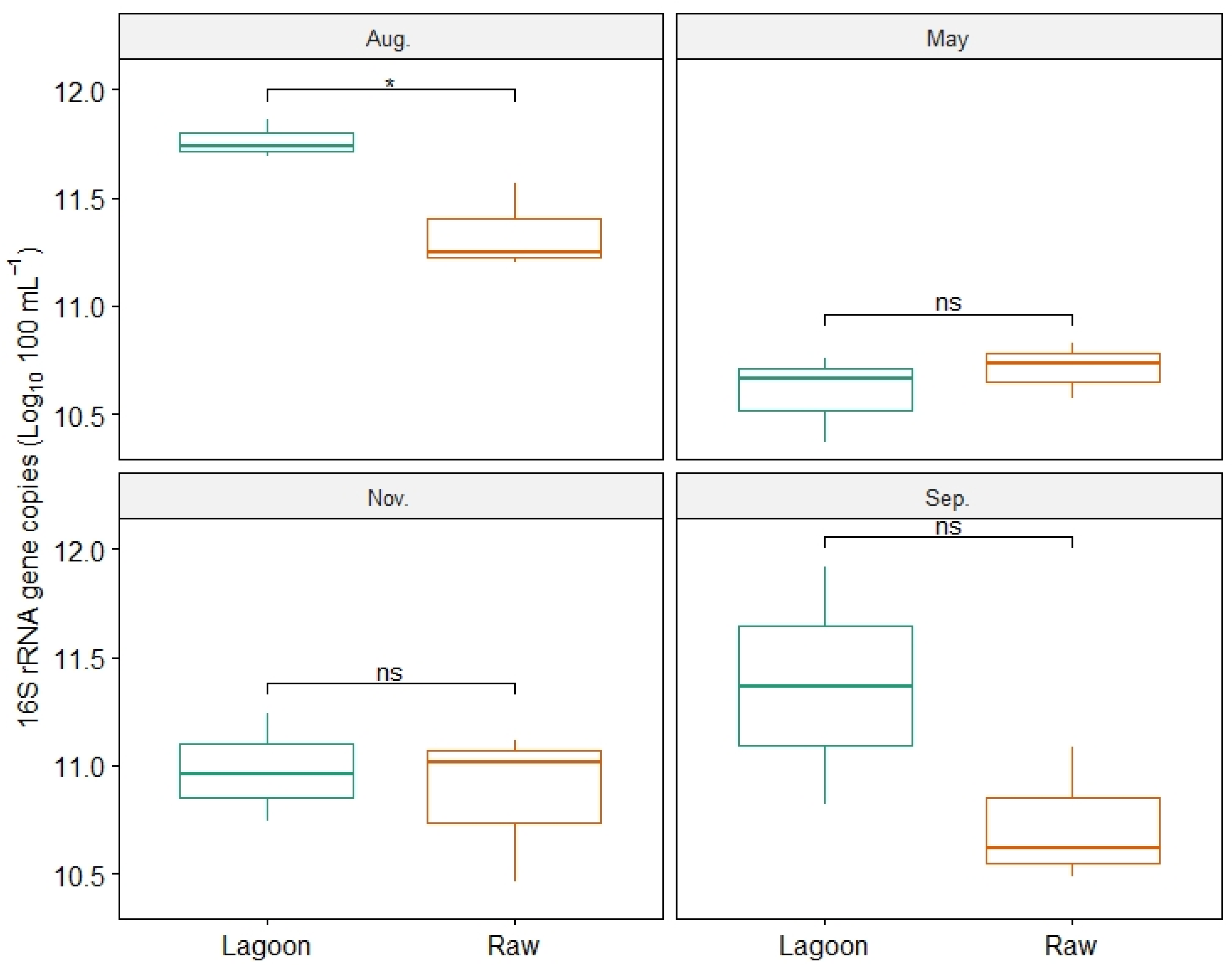
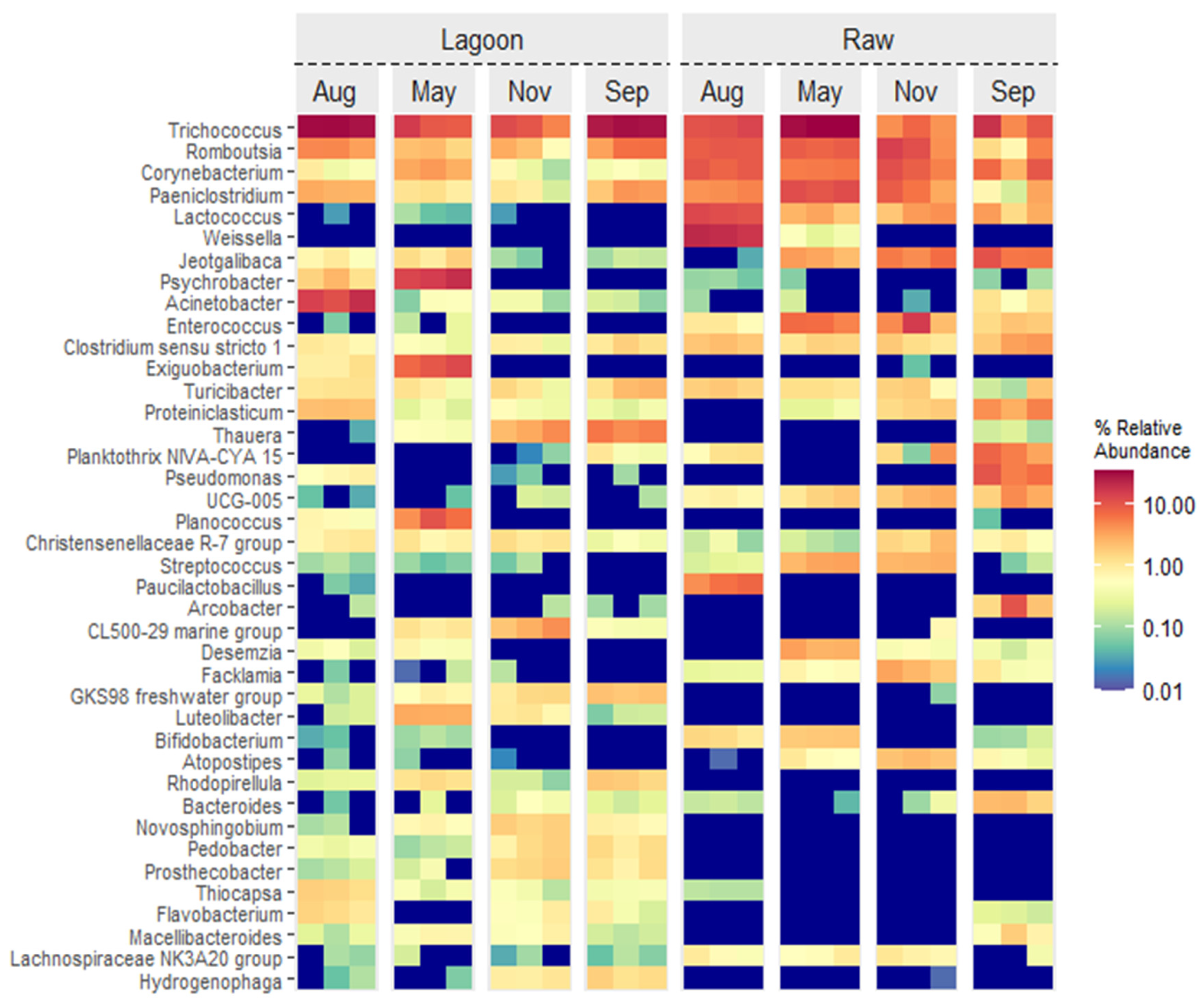
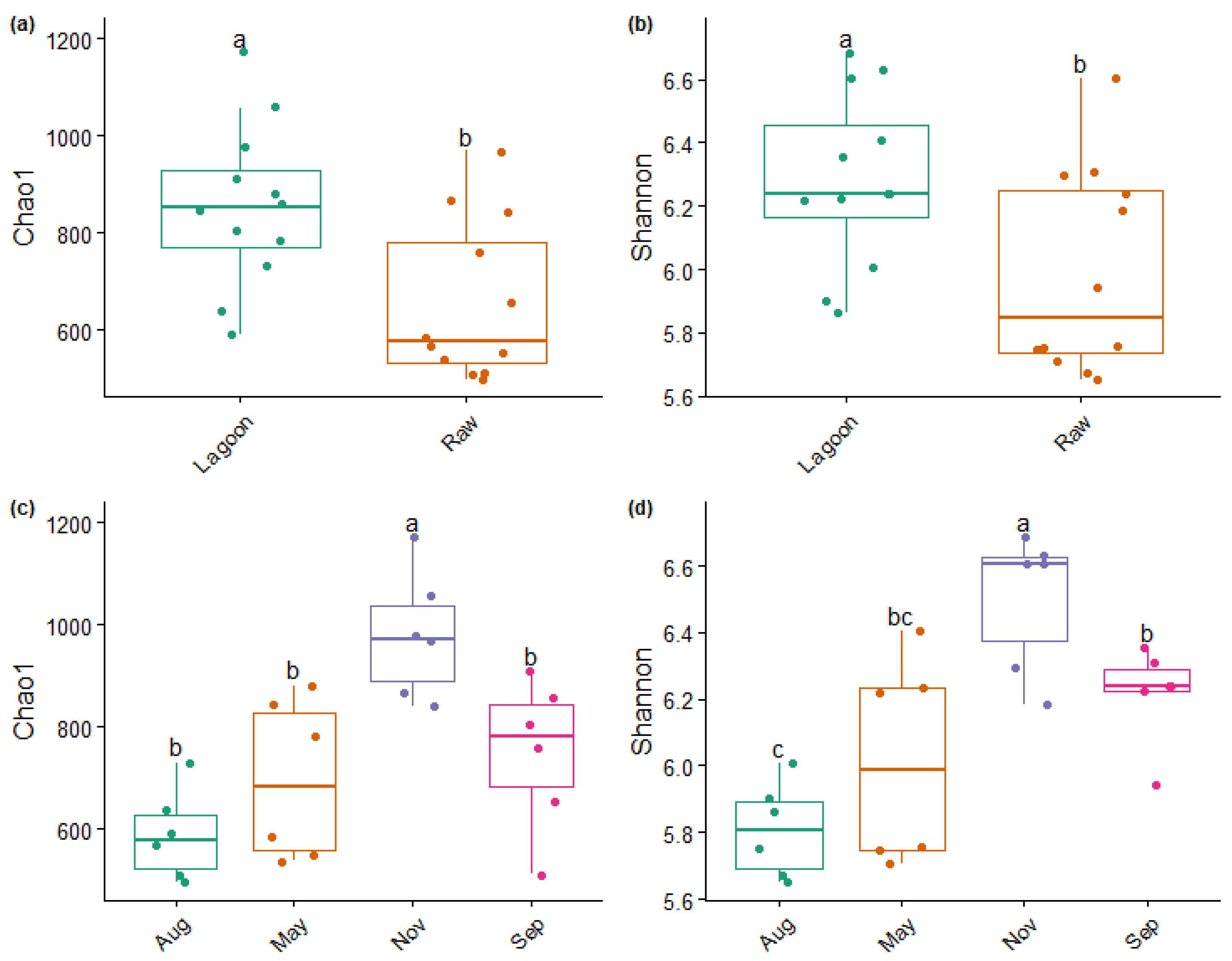
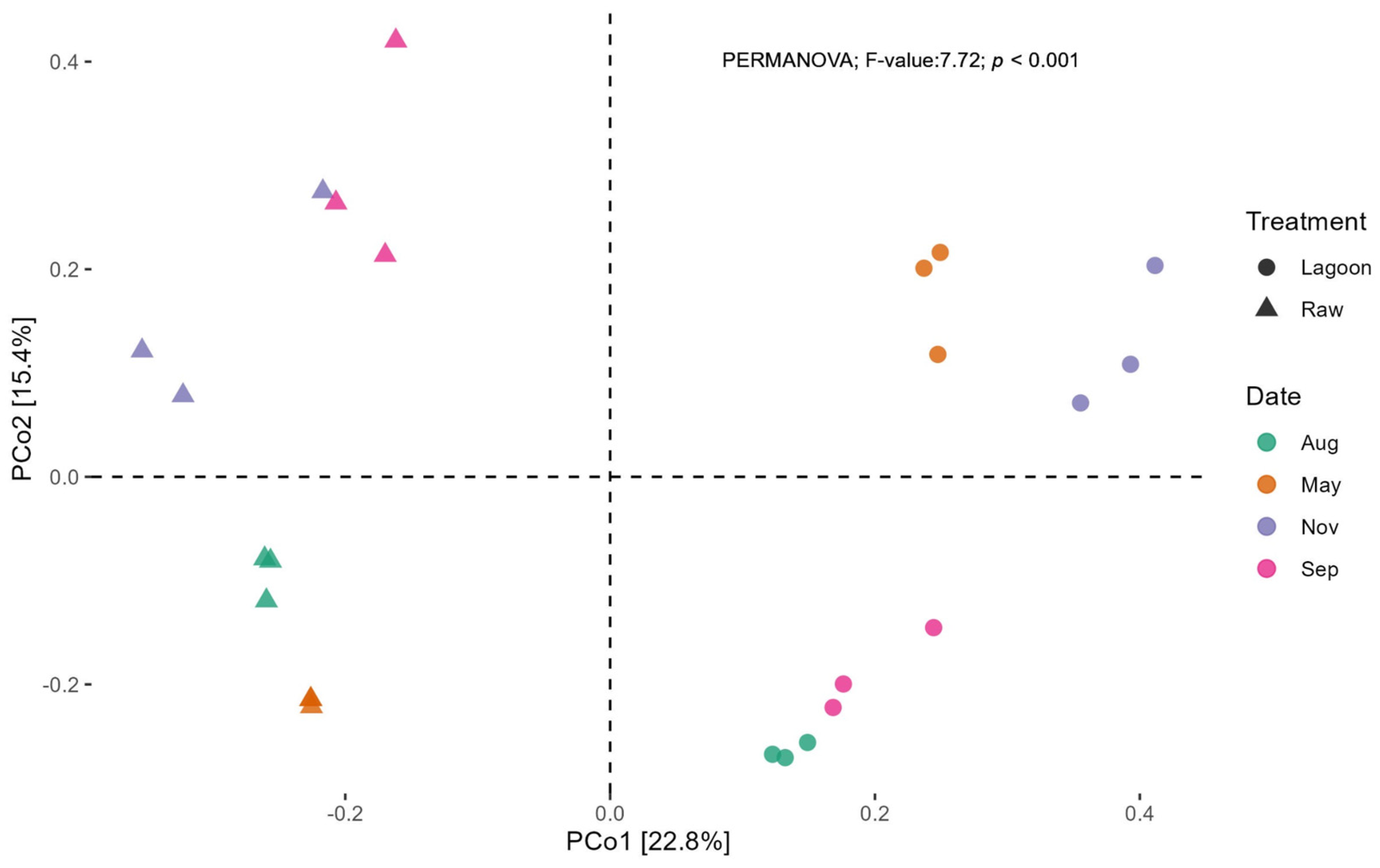
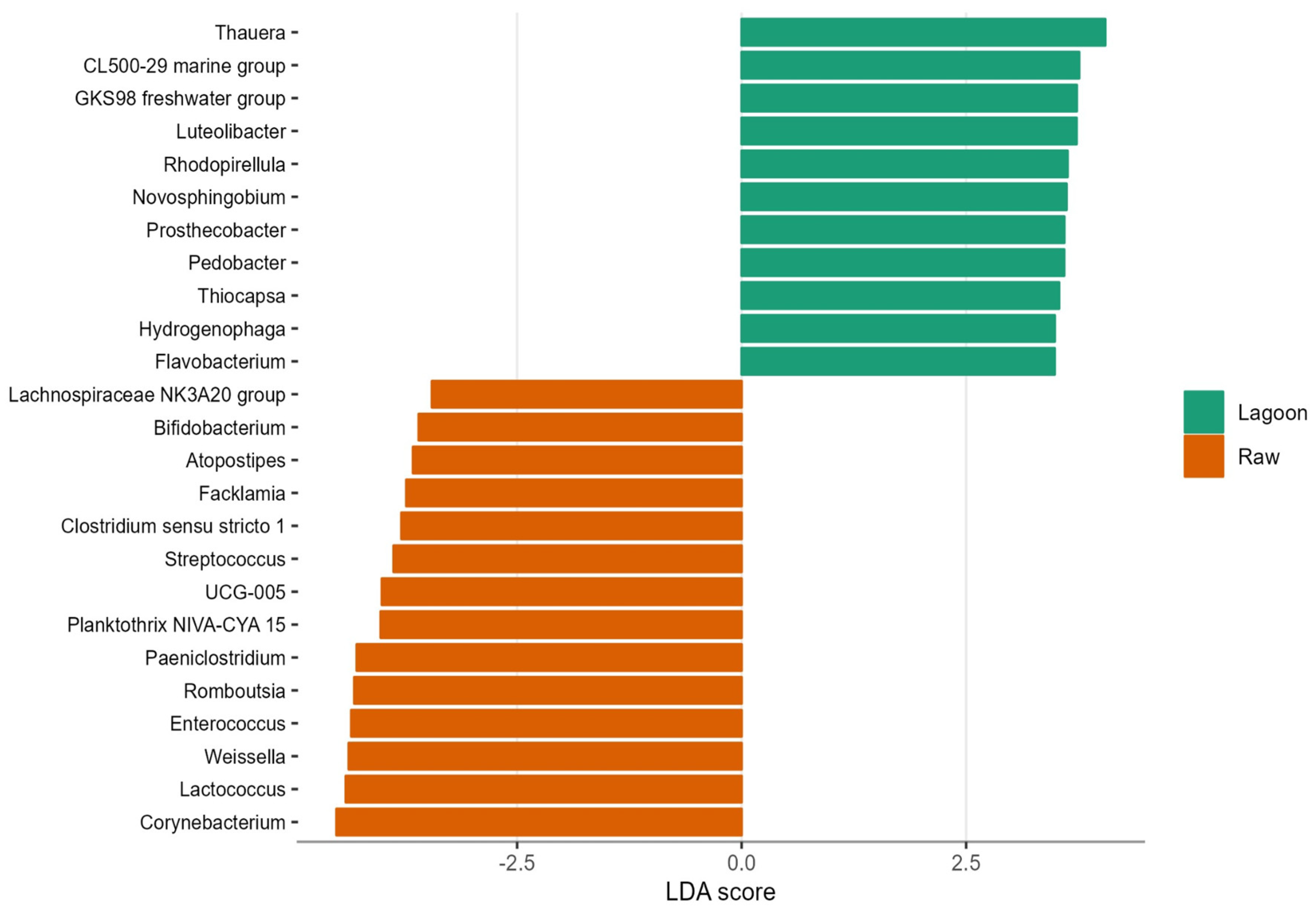
3.3. Bacterial Abundance, Composition, and Diversity of Farm Dairy Effluents
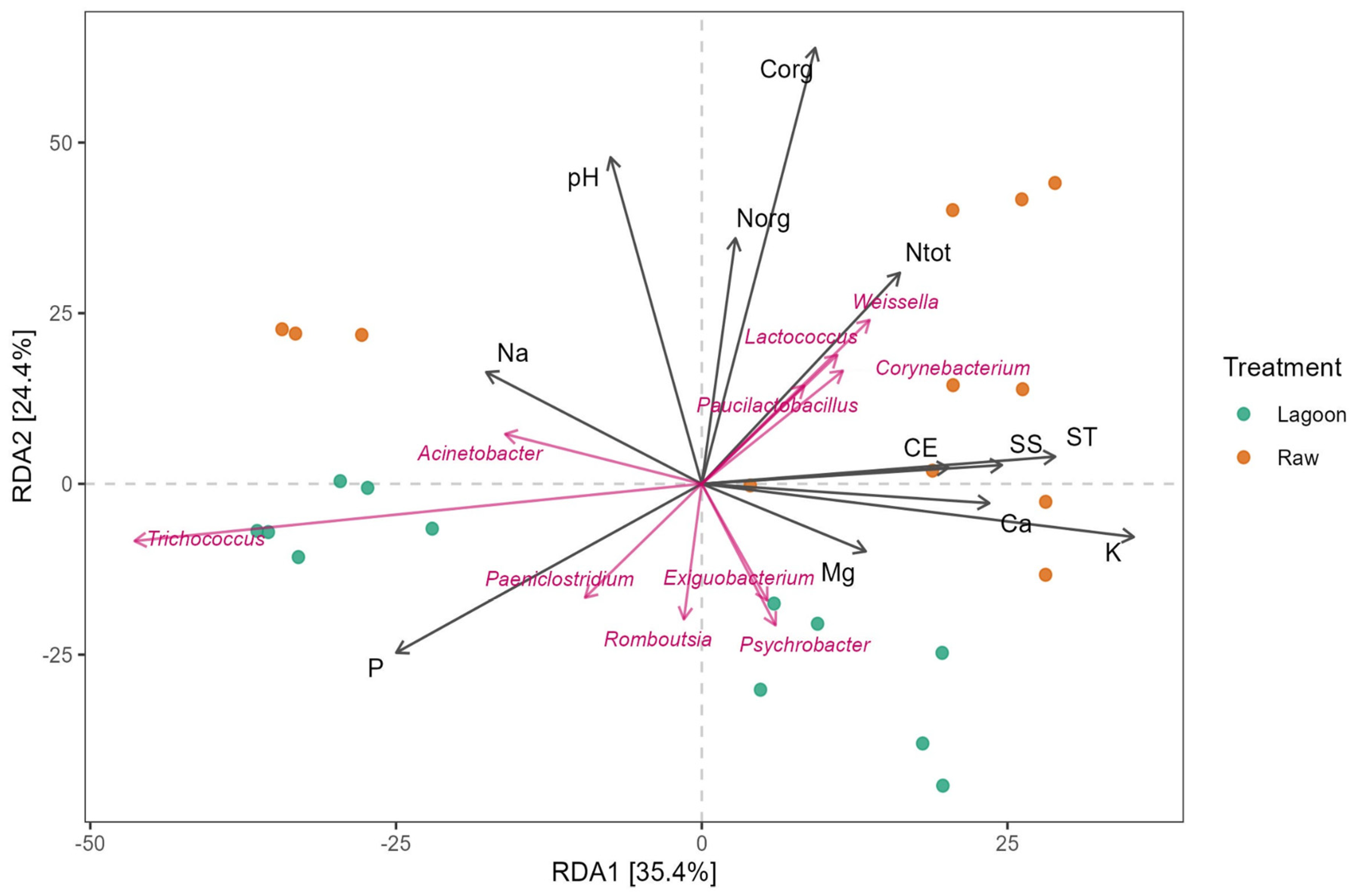
3.4. Linking Bacterial Community with Farm Dairy Effluent Characteristics
4. Discussion
4.1. Bacterial Community Composition Changed with Farm Dairy Effluent Treatment and Season
4.2. Variability in Physicochemical Properties of Farm Dairy Effluents and Their Relation with Bacterial Community Structure
4.3. Risk of Pathogen Loads from Farm Dairy Effluent
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salazar, F.; Herrero, M.A.; Charlón, V.; La Manna, A. Slurry Management in Dairy Grazing Farms in South American Countries. In Proceedings of the 14th International Conference of the FAO ESCORENA Network on the Recycling of Agricultural, Municipal and Industrial Residues in Agriculture, Lisboa, Portugal, 13–15 September 2010; pp. 1–4. [Google Scholar]
- Bhogal, A.; Nicholson, F.A.; Rollett, A.; Taylor, M.; Litterick, A.; Whittingham, M.J.; Williams, J.R. Improvements in the Quality of Agricultural Soils Following Organic Material Additions Depend on Both the Quantity and Quality of the Materials Applied. Front. Sustain. Food Syst. 2018, 2, 9. [Google Scholar] [CrossRef]
- McAllister, T.A.; Topp, E. Role of Livestock in Microbiological Contamination of Water: Commonly the Blame, but Not Always the Source. Anim. Front. 2012, 2, 17–27. [Google Scholar] [CrossRef]
- Alegbeleye, O.O.; Sant’Ana, A.S. Manure-Borne Pathogens as an Important Source of Water Contamination: An Update on the Dynamics of Pathogen Survival/Transport as Well as Practical Risk Mitigation Strategies. Int. J. Hyg. Environ. Health 2020, 227, 113524. [Google Scholar] [CrossRef] [PubMed]
- Von Sperling, M. Wastewater Characteristics, Treatment and Disposal; IWA Publishing: London, UK, 2007; ISBN 1843391619. [Google Scholar]
- Gutiérrez, S.; Cabrera, N. Criterios de Diseño Para Lagunas de Efluentes de Tambos e Condiciones Nacionales. Rev. AIDIS Ing. Cienc. Ambientales. Investig. Desarro. Práctica 2007, 1. [Google Scholar]
- Al-Hazmi, H.E.; Mohammadi, A.; Hejna, A.; Majtacz, J.; Esmaeili, A.; Habibzadeh, S.; Saeb, M.R.; Badawi, M.; Lima, E.C.; Mąkinia, J. Wastewater Reuse in Agriculture: Prospects and Challenges. Environ. Res. 2023, 236, 116711. [Google Scholar] [CrossRef] [PubMed]
- Blaiotta, G.; Di Cerbo, A.; Murru, N.; Coppola, R.; Aponte, M. Persistence of Bacterial Indicators and Zoonotic Pathogens in Contaminated Cattle Wastes. BMC Microbiol. 2016, 16, 87. [Google Scholar] [CrossRef] [PubMed]
- Houlbrooke, D.J.; Horne, D.J.; Hedley, M.J.; Hanly, J.A.; Snow, V.O.; Horne, D.J.; Hedley, M.J.; Hanly, J.A.; Snow, V.O. A Review of Literature on the Land Treatment of Farm—Dairy Effluent in New Zealand and Its Impact on Water Quality. New Zealand J. Agric. Res. 2004, 47, 499–511. [Google Scholar] [CrossRef]
- Hawke, R.M.; Summers, S.A. Effects of Land Application of Farm Dairy Effluent on Soil Properties: A Literature Review. N. Z. J. Agric. Res. 2006, 49, 307–320. [Google Scholar] [CrossRef]
- Hill, D.; Morra, M.J.; Stalder, T.; Jechalke, S.; Top, E.; Pollard, A.T.; Popova, I. Dairy Manure as a Potential Source of Crop Nutrients and Environmental Contaminants. J. Environ. Sci. 2021, 100, 117–130. [Google Scholar] [CrossRef]
- Akari, M.; Uchida, Y. Survival Rates of Microbial Communities from Livestock Waste to Soils: A Comparison between Compost and Digestate. Appl. Environ. Soil. Sci. 2021, 2021, 6645203. [Google Scholar] [CrossRef]
- Coelho, J.J.; Hennessy, A.; Casey, I.; Roberto, C.; Bragança, S.; Woodcock, T.; Kennedy, N. Biofertilisation with Anaerobic Digestates: A Field Study of Effects on Soil Microbial Abundance and Diversity. Appl. Soil Ecol. 2019, 147, 103403. [Google Scholar] [CrossRef]
- Qi, G.; Pan, Z.; Sugawa, Y.; Andriamanohiarisoamanana, F.J.; Yamashiro, T.; Iwasaki, M.; Kawamoto, K.; Ihara, I.; Umetsu, K. Comparative Fertilizer Properties of Digestates from Mesophilic and Thermophilic Anaerobic Digestion of Dairy Manure: Focusing on Plant Growth Promoting Bacteria (PGPB) and Environmental Risk. J. Mater. Cycles Waste Manag. 2018, 20, 1448–1457. [Google Scholar] [CrossRef]
- Irazoqui, J.M.; Eberhardt, M.F.; Adjad, M.M.; Amadio, A.F. Identification of Key Microorganisms in Facultative Stabilization Ponds from Dairy Industries, Using Metagenomics. PeerJ 2022, 10, e12772. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Mebius, L.J. A Rapid Method for the Determination of Organic Carbon in Soil. Anal. Chim. Acta 1960, 22, 120–124. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Total Nitrogen. In Methods of Soil Analysis—Part 2—Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 595–624. [Google Scholar]
- Rhine, E.D.; Mulvaney, R.L.; Pratt, E.J.; Sims, G.K. Improving the Berthelot Reaction for Determining Ammonium in Soil Extracts and Water. Soil Sci. Soc. Am. J. 1998, 62, 473. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A Modified Single Solution Method for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Isaac, R.A.; Kerber, J.D. Atomic Absorption and Flame Photometry: Techniques and Uses in Soil, Plant, and Water Analysis. In Instrumental Methods for Analysis of Soils and Plant Tissue; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1971; pp. 17–37. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R Package for Data Mining in Microbial Community Ecology. FEMS Microbiol. Ecol. 2021, 97, 255. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. Permutation Tests for Univariate or Multivariate Analysis of Variance and Regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Anderson, M. Distance-Based Tests for Homogeneity of Multivariate Dispersions. J. Int. Biom. Soc. 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J. Vegan: Ecological Diversity. R Proj. 2022, 368, 1–11. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Chiu, C.; Miao, M.; Wang, Y.; Settles, M.; Del Rio, N.S.; Castillo, A.; Souza, A.; Pereira, R.; Jeannotte, R. 16S RRNA Analysis of Diversity of Manure Microbial Community in Dairy Farm Environment. PLoS ONE 2018, 13, e0190126. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wang, N.; Wei, D.; Liang, C.; Chen, X.; Liu, L.; Shi, J. Bacterial Community Compositions and Nitrogen Metabolism Function in a Cattle Farm Wastewater Treatment Plant Revealed by Illumina High-Throughput Sequencing. Environ. Sci. Pollut. Res. 2021, 28, 40895–40907. [Google Scholar] [CrossRef] [PubMed]
- McGarvey, J.A.; Miller, W.G.; Zhang, R.; Ma, Y.; Mitloehner, F. Bacterial Population Dynamics in Dairy Waste during Aerobic and Anaerobic Treatment and Subsequent Storage. Appl. Environ. Microbiol. 2007, 73, 193–202. [Google Scholar] [CrossRef]
- Chang, Y.L.; Nagarajan, D.; Chen, J.H.; Chen, C.Y.; Wu, Y.J.; Whang, L.M.; Lee, D.J.; Chang, J.S. Microalgae-Bacteria Consortia for the Treatment of Raw Dairy Manure Wastewater Using a Novel Two-Stage Process: Process Optimization and Bacterial Community Analysis. Chem. Eng. J. 2023, 473, 145388. [Google Scholar] [CrossRef]
- Isobe, K.; Ohte, N. Ecological Perspectives on Microbes Involved in N-Cycling. Microbes Environ. 2014, 29, 4–16. [Google Scholar] [CrossRef]
- Ren, T.; Chi, Y.; Wang, Y.; Shi, X.; Jin, X.; Jin, P. Diversified Metabolism Makes Novel Thauera Strain Highly Competitive in Low Carbon Wastewater Treatment. Water Res. 2021, 206, 117742. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Cho, G.S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M.A.P. The Genus Weissella: Taxonomy, Ecology and Biotechnological Potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef]
- Velázquez-Ordoñez, V.; Valladares-Carranza, B.; Tenorio-Borroto, E.; Talavera-Rojas, M.; Antonio Varela-Guerrero, J.; Acosta-Dibarrat, J.; Puigvert, F.; Grille, L.; González Revello, Á.; Pareja, L. Microbial Contamination in Milk Quality and Health Risk of the Consumers of Raw Milk and Dairy Products. In Nutrition in Health and Disease—Our Challenges Now and Forthcoming Time; Mozsik, G., Figler, M., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Habtewold, J.; Gordon, R.; Sokolov, V.; VanderZaag, A.; Wagner-Riddle, C.; Dunfield, K. Targeting Bacteria and Methanogens to Understand the Role of Residual Slurry as an Inoculant in Stored Liquid Dairy Manure. Appl. Environ. Microbiol. 2018, 84, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Shi, F.; Qian, S.; Ma, J. Variations in the Dairy Wastewater Properties and Bacterial Diversity of Each Unit in Membrane-Enclosed Anaerobic Lagoon Treatment Process in Heilongjiang Province. J. Environ. Manag. 2023, 347, 119035. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct Soil Microbial Diversity under Long-Term Organic and Conventional Farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef]
- Sun, L.; Pope, P.B.; Eijsink, V.G.H.; Schnürer, A. Characterization of Microbial Community Structure during Continuous Anaerobic Digestion of Straw and Cow Manure. Microb. Biotechnol. 2015, 8, 815–827. [Google Scholar] [CrossRef]
- Blasco, L.; Kahala, M.; Ervasti, S.; Tampio, E. Dynamics of Microbial Community in Response to Co-Feedstock Composition in Anaerobic Digestion. Bioresour. Technol. 2022, 364, 128039. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, R.; Zhang, C.; Ding, S.; Ying, M.; Shan, S. In Situ Analysis of Antibiotic Resistance Genes in Anaerobically Digested Dairy Manure and Its Subsequent Disposal Facilities. Bioresour. Technol. 2021, 333, 124988. [Google Scholar] [CrossRef]
- Nelson, M.C.; Morrison, M.; Yu, Z. A Meta-Analysis of the Microbial Diversity Observed in Anaerobic Digesters. Bioresour. Technol. 2011, 102, 3730–3739. [Google Scholar] [CrossRef]
- Villarroel-Schneider, J.; Höglund-Isaksson, L.; Mainali, B.; Martí-Herrero, J.; Cardozo, E.; Malmquist, A.; Martin, A. Energy Self-Sufficiency and Greenhouse Gas Emission Reductions in Latin American Dairy Farms through Massive Implementation of Biogas-Based Solutions. Energy Convers. Manag. 2022, 261, 115670. [Google Scholar] [CrossRef]
- Chen, X.; Lang, X.L.; Xu, A.L.; Song, Z.W.; Yang, J.; Guo, M.Y. Seasonal Variability in the Microbial Community and Pathogens in Wastewater Final Effluents. Water 2019, 11, 2586. [Google Scholar] [CrossRef]
- Szyłak-Szydłowski, M.; Kulig, A.; Miaśkiewicz-Pęska, E. Seasonal Changes in the Concentrations of Airborne Bacteria Emitted from a Large Wastewater Treatment Plant. Int. Biodeterior. Biodegrad. 2016, 115, 11–16. [Google Scholar] [CrossRef]
- Wang, Y.; Sutton, N.B.; Zheng, Y.; Dong, H.; Rijnaarts, H.H.M. Seasonal Variation in Antibiotic Resistance Genes and Bacterial Phenotypes in Swine Wastewater during Three-Chamber Anaerobic Pond Treatment. Environ. Res. 2023, 216, 114495. [Google Scholar] [CrossRef]
- Ducey, T.F.; Rashash, D.M.C.; Szogi, A.A. Differences in Microbial Communities and Pathogen Survival between a Covered and Uncovered Anaerobic Lagoon. Environments 2019, 6, 91. [Google Scholar] [CrossRef]
- Ducey, T.F.; Hunt, P.G. Microbial Community Analysis of Swine Wastewater Anaerobic Lagoons Bynext-Generation DNA Sequencing. Anaerobe 2013, 21, 50–57. [Google Scholar] [CrossRef]
- Illarze, G.; del Pino, A.; Rodríguez-Blanco, A.; Irisarri, P. Application of Dairy Effluents to Pastures Affects Soil Nitrogen Dynamics and Microbial Activity. Agronomy 2023, 13, 470. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, L.; Liu, J.; Guo, D.; Zhu, Z.; Dong, H. Impact of Anaerobic Digestion on Reactive Nitrogen Gas Emissions from Dairy Slurry Storage. J. Environ. Manag. 2022, 316, 115306. [Google Scholar] [CrossRef]
- Thurston-Enriquez, J.A.; Gilley, J.E.; Eghball, B. Microbial Quality of Runoff Following Land Application of Cattle Manure and Swine Slurry. J. Water Health 2005, 3, 157–171. [Google Scholar] [CrossRef]
- Decreto N° 253/979 Artículo 11 Aprobación de Normativa Para Prevenir La Contaminación Ambiental, a Través Del Control de Las Aguas. Available online: https://www.impo.com.uy/bases/decretos/253-1979 (accessed on 15 January 2024).
- Rahmati, E.; Martin, V.; Wong, D.; Sattler, F.; Petterson, J.; Ward, P.; Butler-Wu, S.M.; She, R.C. Facklamia Species as an Underrecognized Pathogen. Open Forum Infect. Dis. 2017, 4, ofw272. [Google Scholar] [CrossRef]
- Dungan, R.S.; Klein, M.; Leytem, A.B. Quantification of Bacterial Indicators and Zoonotic Pathogens in Dairy Wastewater Ponds. Appl. Environ. Microbiol. 2012, 78, 8089–8095. [Google Scholar] [CrossRef]
- Ramos, T.D.M.; Jay-Russell, M.T.; Millner, P.D.; Baron, J.N.; Stover, J.; Pagliari, P.; Hutchinson, M.; Lilley, J.; Rowley, N.; Haghani, V.; et al. Survival and Persistence of Foodborne Pathogens in Manure-Amended Soils and Prevalence on Fresh Produce in Certified Organic Farms: A Multi-Regional Baseline Analysis. Front. Sustain. Food Syst. 2021, 5, 674767. [Google Scholar] [CrossRef]
- Rapp, D.; Ross, C.; Hea, S.Y.; Brightwell, G. Importance of the Farm Environment and Wildlife for Transmission of Campylobacter Jejuni in a Pasture-Based Dairy Herd. Microorganisms 2020, 8, 1877. [Google Scholar] [CrossRef]
- Ross, C.; Donnison, A. Campylobacter and Farm Dairy Effluent Irrigation. N. Z. J. Agric. Res. 2003, 46, 255–262. [Google Scholar] [CrossRef]
- Baker, C.A.; Almeida, G.; Lee, J.A.; Gibson, K.E. Pathogen and Surrogate Survival in Relation to Fecal Indicator Bacteria in Freshwater Mesocosms. Appl. Environ. Microbiol. 2021, 87, 1–14. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, J.; Mao, X. Removal of Pathogens in Onsite Wastewater Treatment Systems: A Review of Design Considerations and Influencing Factors. Water 2021, 13, 1190. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mg L−1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | Effluent | pH | EC (mS m−1) | TS (%) | SS (%) | Corg | TKN | NH4+-N | P | K+ | Na+ | Mg2+ | Ca2+ |
| September | Raw | 8.5 ± 0.0 a | 2.4 ± 0.0 a | 0.8 ± 0.4 a | 0.8 ± 0.4 a | 514 ± 80 a | 800 ± 39 a | 216 ± 23 a | 16 ± 1 b | 722 ± 88 a | 217 ± 17 a | 100 ± 26 a | 197 ± 61 a |
| Lagoon | 7.7 ± 0.0 b | 2.1 ± 0.1 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 359 ± 12 b | 137 ± 14 b | 87 ± 15 b | 38 ± 1 a | 150 ± 18 b | 144 ± 17 b | 31 ± 4 b | 48 ± 5 b | |
| November | Raw | 8.2 ± 0.4 a | 6.9 ± 1.2 a | 0.3 ± 1 a | 0.3 ± 1 a | 643 ± 35 a | 538 ± 11 a | 332 ± 47 a | 84 ± 19 a | 436 ± 107 a | 131 ± 28 a | 46 ± 9 a | 76 ± 15 a |
| Lagoon | 7.7 ± 0.0 a | 2.3 ± 0.1 b | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 414 ± 19 b | 95 ± 7 b | 39 ± 1 b | 36 ± 3 b | 156 ± 13 b | 75 ± 8 b | 26 ± 3 b | 47 ± 5 b | |
| May | Raw | 7.3 ± 0.0 b | 4.8 ± 0.1 a | 0.6 ± 0.1 a | 0.6 ± 0.1 a | 736 ± 145 a | 539 ± 9 a | 75 ± 10 a | 101 ± 5 a | 277 ± 21 a | 226 ± 19 a | 73 ± 5 a | 87 ± 5 a |
| Lagoon | 7.8 ± 0.1 a | 3.8 ± 0.0 b | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 275 ± 18 b | 88 ± 2 b | 37 ± 1 b | 24 ± 4 b | 224 ± 27 a | 157 ± 15 b | 38 ± 6 b | 32 ± 6 b | |
| August | Raw | 6.0 ± 0.0 b | 1.8 ± 0.0 a | 0.1 ± 0.0 a | 0.1 ± 0.0 a | 890 ± 22 a | 281 ± 17 a | 19 ± 1 b | 19 ± 2 b | 69 ± 4 b | 16 ± 1 b | 8.3 ± 1 b | 15 ± 1 b |
| Lagoon | 8.1 ± 0.0 a | 2.3 ± 0.4 a | 0.2 ± 0.0 a | 0.2 ± 0.0 a | 573 ± 42 b | 231 ± 5 b | 53 ± 4 a | 26 ± 2 a | 92 ± 12 a | 52 ± 7 a | 17 ± 2 a | 23 ± 3 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Illarze, G.; del Pino, A.; Irisarri, P. Differences in Bacterial Communities and Pathogen Indicators of Raw and Lagoon-Stabilized Farm Dairy Effluents. Microorganisms 2024, 12, 305. https://doi.org/10.3390/microorganisms12020305
Illarze G, del Pino A, Irisarri P. Differences in Bacterial Communities and Pathogen Indicators of Raw and Lagoon-Stabilized Farm Dairy Effluents. Microorganisms. 2024; 12(2):305. https://doi.org/10.3390/microorganisms12020305
Chicago/Turabian StyleIllarze, Gabriela, Amabelia del Pino, and Pilar Irisarri. 2024. "Differences in Bacterial Communities and Pathogen Indicators of Raw and Lagoon-Stabilized Farm Dairy Effluents" Microorganisms 12, no. 2: 305. https://doi.org/10.3390/microorganisms12020305