

The Mutation of the DNA-Binding Domain of Fur Protein Enhances the Pathogenicity of Edwardsiella piscicida via Inducing Overpowering Pyroptosis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Cell Lines, and Fish
2.2. Construction of In-Frame Deletion Mutation of Fur
2.3. Growth Curve Measurement
2.4. Detection of Siderophore Production
2.5. Biofilm Formation Assay
2.6. Swimming Motility Assays
2.7. Resistance to Environmental Stress (Acid, Manganese, and Temperature)
2.8. The Flagella Observation by Transmission Electron Microscopy (TEM)
2.9. Bacterial Invasion of Eukaryotic Cell Lines
2.10. Fluorescence Microscopy
2.11. Pyroptosis Analysis
2.12. Cytotoxicity Assay and IL-1β Detection
2.13. Experimental Challenge of Bacterial Disseminations In Vivo
2.14. Reactive Oxygen Species (ROS) Production
2.15. Statistical Analysis
3. Results
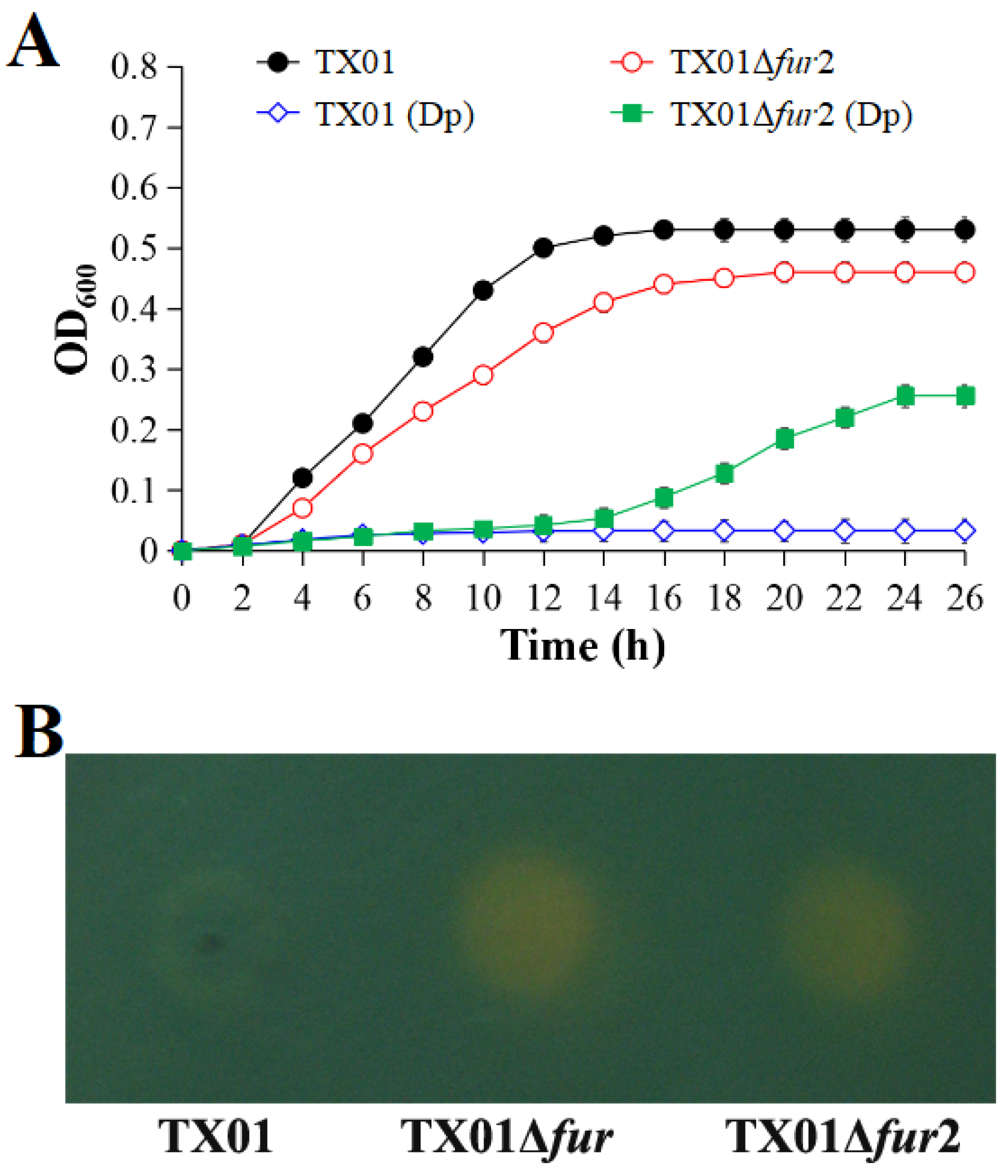
3.1. Effect on Growth and Siderophore Production
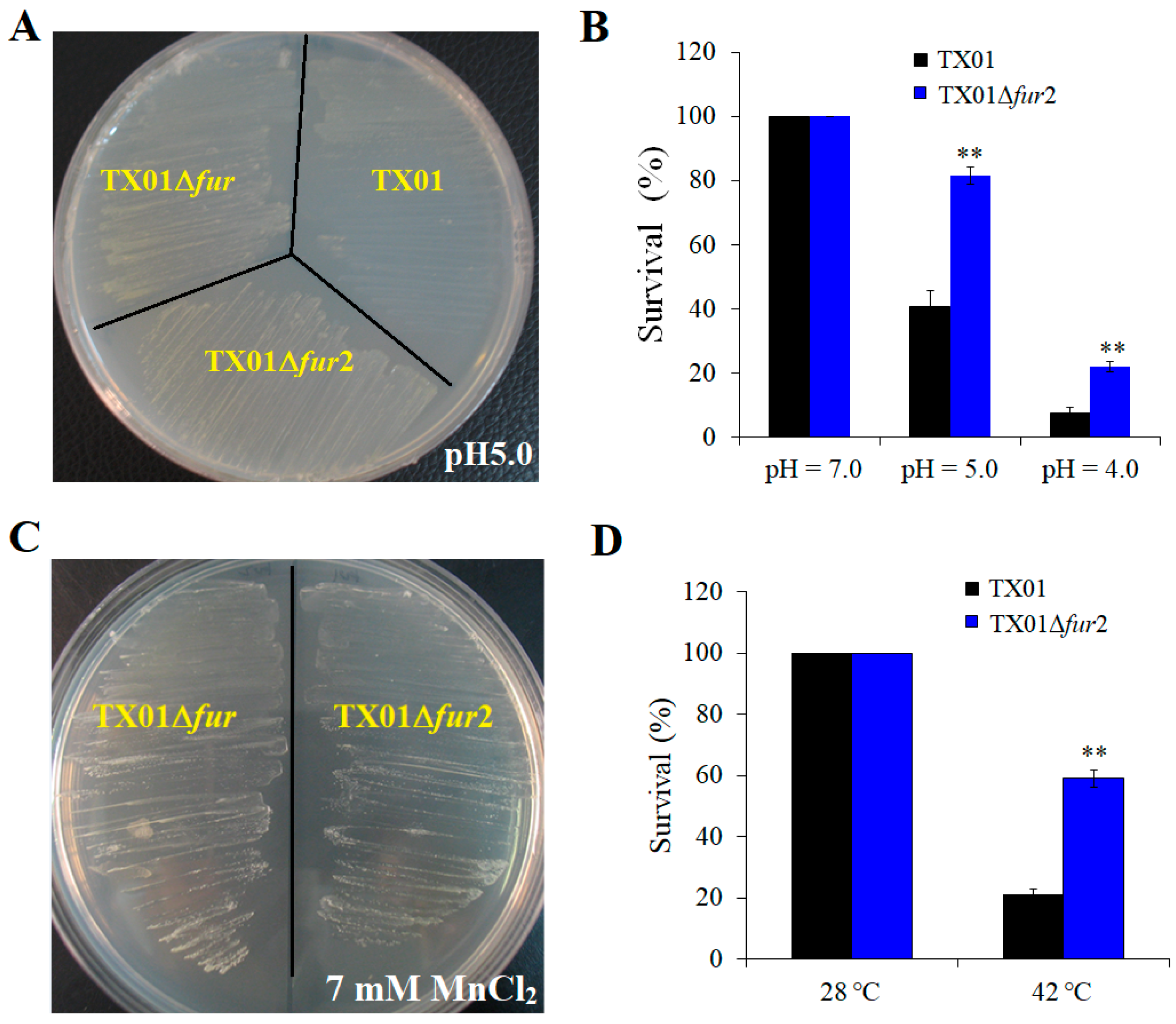
3.2. Effect on Resistance against Environmental Stress
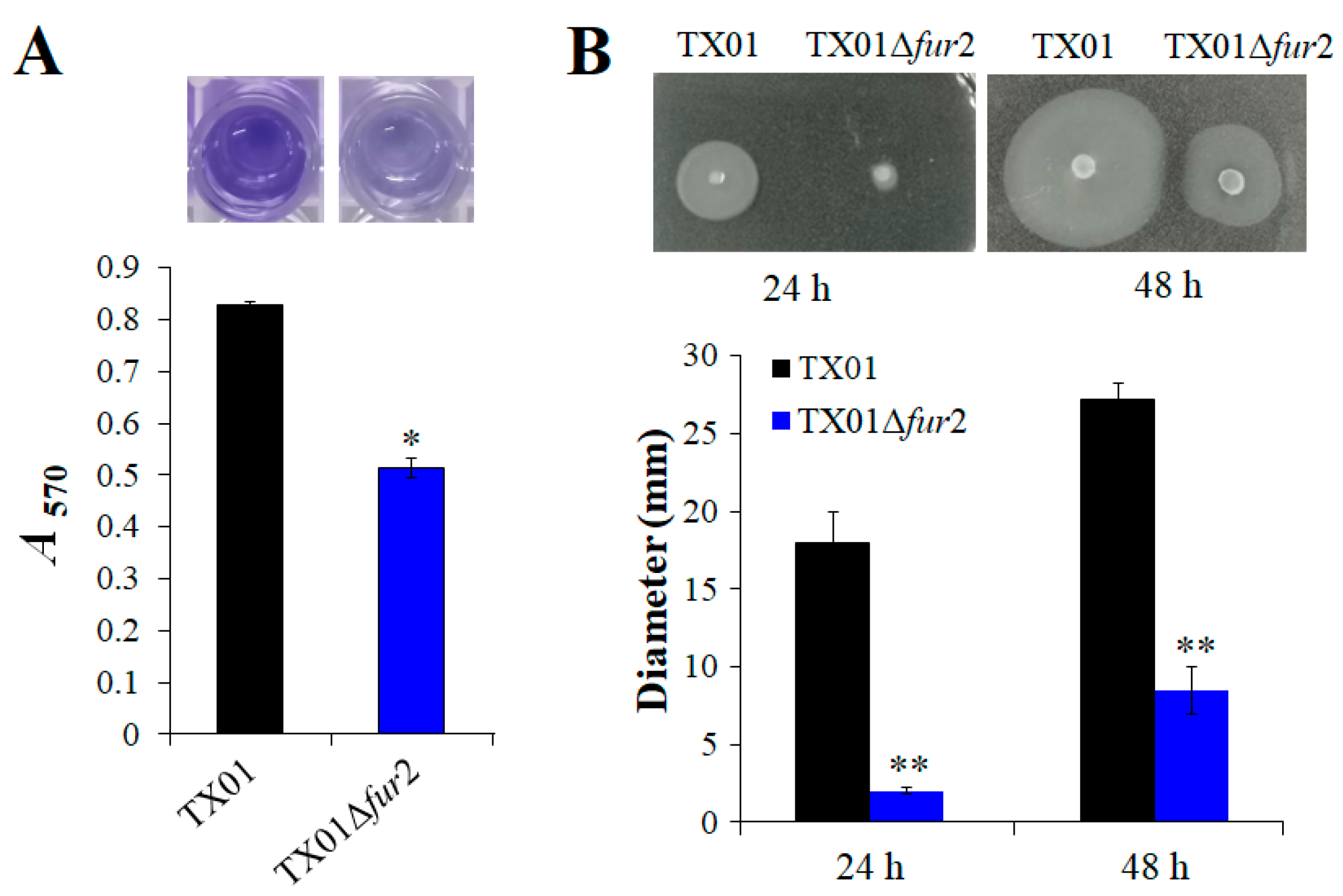
3.3. Effect on the Abilities of Biofilm Formation and Motility
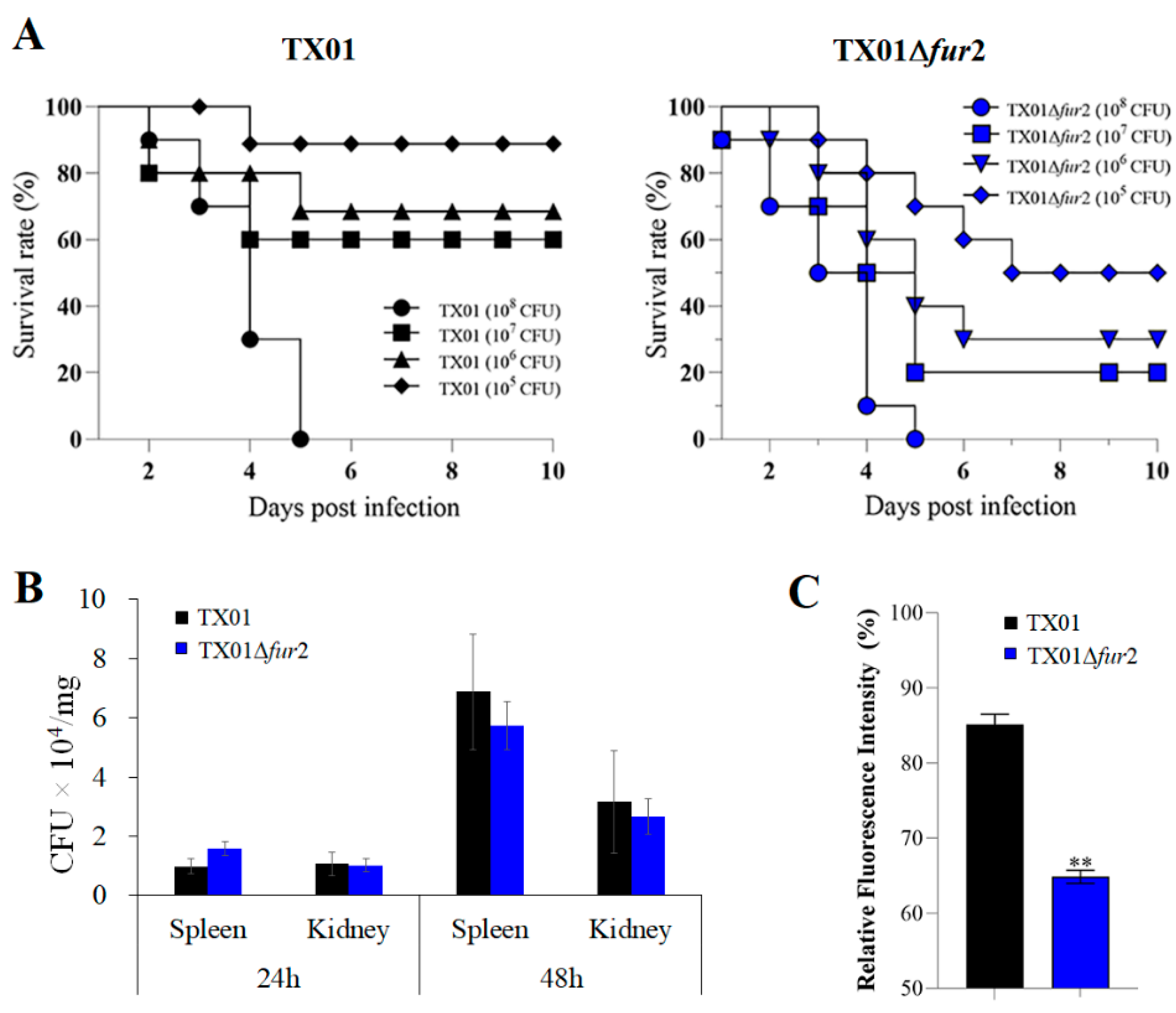
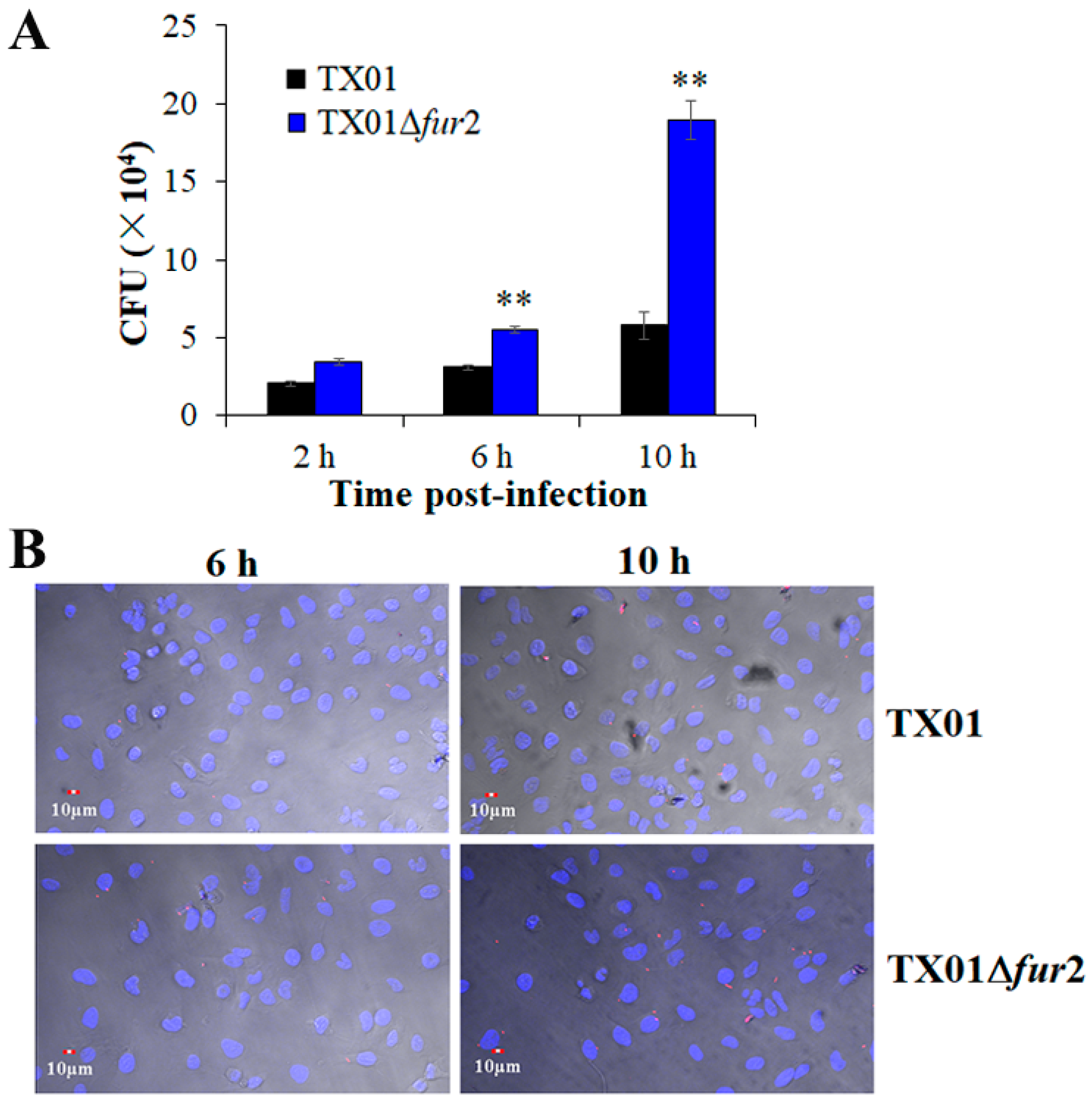
3.4. Effects on Lethality and Infectivity In Vivo
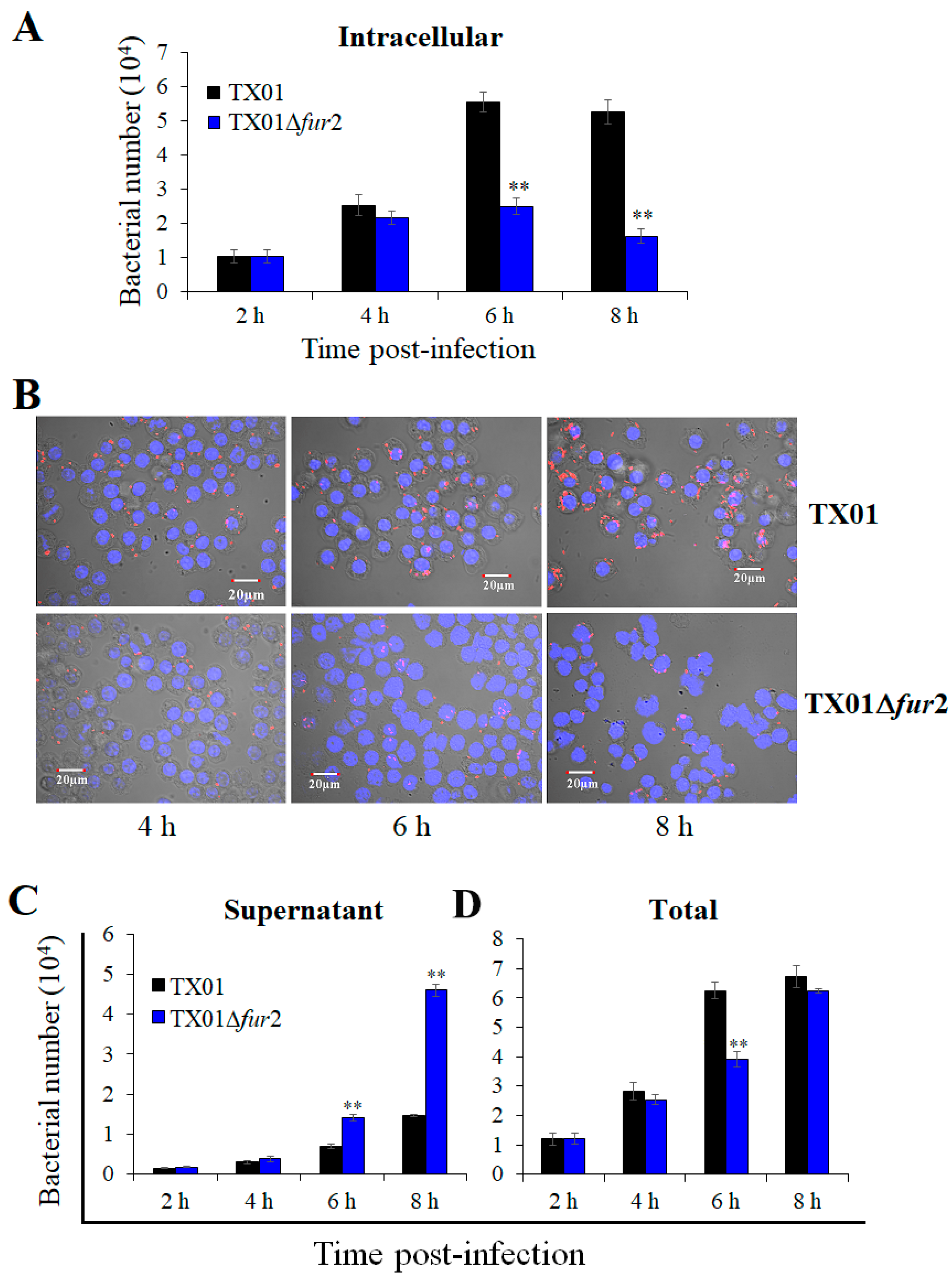
3.5. Effects on Invasion and Replication In Vitro
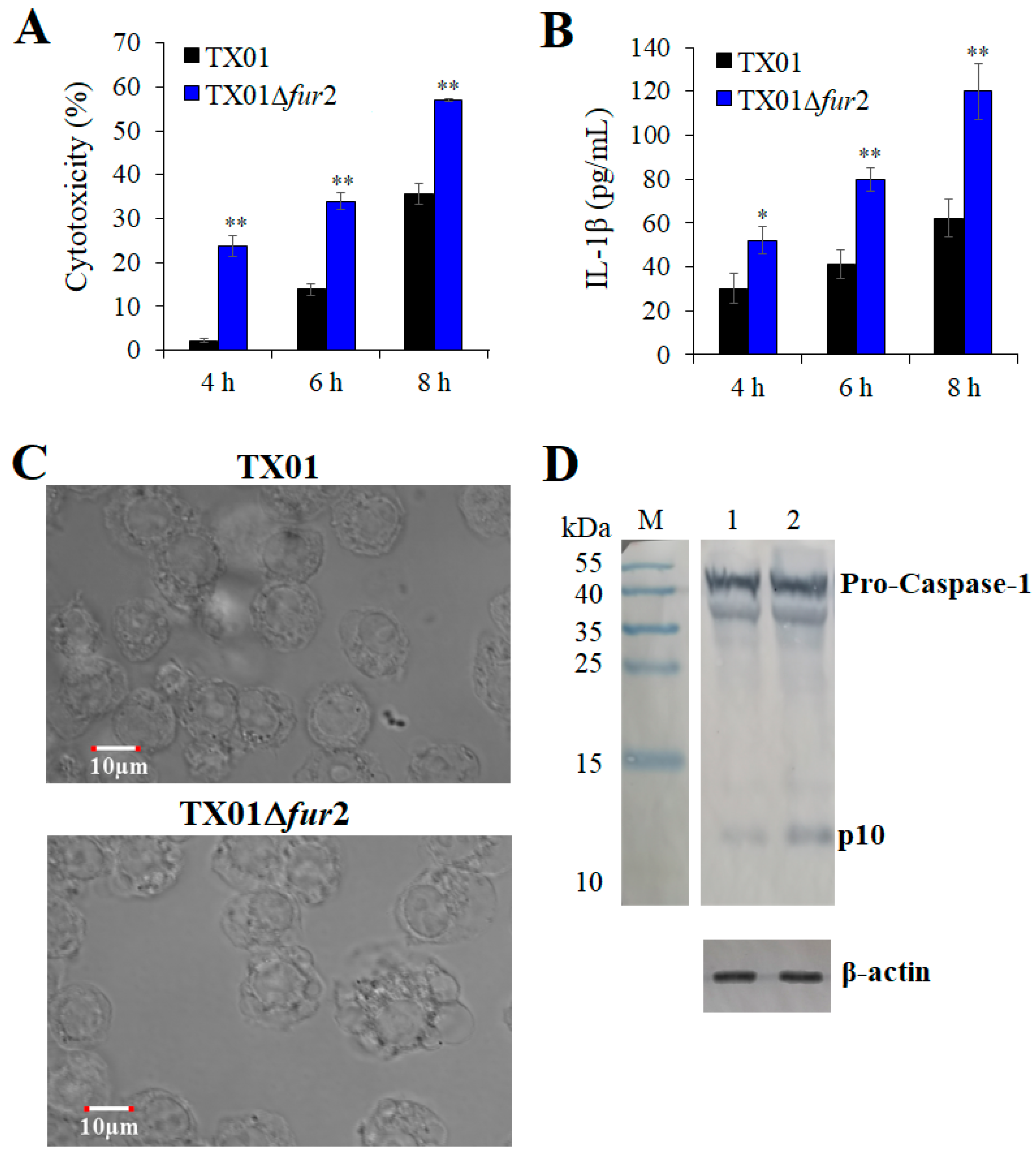
3.6. Effect on the Inflammatory Responses of Macrophage Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nairz, M.; Weiss, G. Iron in Infection and Immunity. Mol. Aspects. Med. 2020, 75, 100864. [Google Scholar] [CrossRef] [PubMed]
- Zughaier, S.M.; Cornelis, P. Editorial: Role of Iron in Bacterial Pathogenesis. Front. Cell. Infect. Microbiol. 2018, 8, 344. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, W.; Wu, S.; Gao, H. Recent Advances in the Siderophore Biology of Shewanella. Front. Microbiol. 2022, 13, 823758. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, J.R.; Laakso, H.A.; Heinrichs, D.E. Iron Acquisition Strategies of Bacterial Pathogens. Microbiol. Spectr. 2016, 4, 43–85. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; di Masi, A.; Leboffe, L.; Frangipani, E.; Nardini, M.; Verde, C.; Visca, P. Structural Biology of Bacterial Haemophores. Adv. Microb. Physiol. 2015, 67, 127–176. [Google Scholar] [CrossRef] [PubMed]
- Cescau, S.; Cwerman, H.; Létoffé, S.; Delepelaire, P.; Wandersman, C.; Biville, F. Heme Acquisition by Hemophores. Biometals 2007, 20, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Wandersman, C.; Delepelaire, P. Bacterial iron sources: From siderophores to hemophores. Annu. Rev. Microbiol. 2004, 58, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Cassat, J.E.; Skaar, E.P. Iron in Infection and Immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- van Vliet, A.H.; Stoof, J.; Vlasblom, R.; Wainwright, S.A.; Hughes, N.J.; Kelly, D.J.; Bereswill, S.; Bijlsma, J.J.; Hoogenboezem, T.; Vandenbroucke-Grauls, C.M.; et al. The role of the Ferric Uptake Regulator (Fur) in regulation of Helicobacter pylori iron uptake. Helicobacter 2002, 7, 237–244. [Google Scholar] [CrossRef]
- D’Aquino, J.A.; Lattimer, J.R.; Denninger, A.; D’Aquino, K.E.; Ringe, D. Role of the N-Terminal Helix in the Metal Ion-Induced Activation of the Diphtheria Toxin Repressor DtxR. Biochemistry 2007, 46, 11761–11770. [Google Scholar] [CrossRef] [PubMed]
- Hantke, K. Iron and metal regulation in bacteria. Curr. Opin. Microbiol. 2001, 4, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Stojiljkovic, I.; Hantke, K. Functional domains of the Escherichia coli ferric uptake regulator protein (Fur). Mol. Gen. Genet. 1995, 247, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.W.; Bogard, R.W.; Mekalanos, J.J. Mapping the regulon of Vibrio cholerae ferric uptake regulator expands its known network of gene regulation. Proc. Natl. Acad. Sci. USA 2011, 108, 12467–12472. [Google Scholar] [CrossRef]
- Pasqua, M.; Visaggio, D.; Lo Sciuto, A.; Genah, S.; Banin, E.; Visca, P.; Imperi, F. Ferric Uptake Regulator Fur Is Conditionally Essential in Pseudomonas aeruginosa. J. Bacteriol. 2017, 199, e00472-17. [Google Scholar] [CrossRef] [PubMed]
- Pennella, M.A.; Giedroc, D.P. Structural Determinants of Metal Selectivity in Prokaryotic Metal-responsive Transcriptional Regulators. Biometals 2005, 18, 413–428. [Google Scholar] [CrossRef] [PubMed]
- Pym, A.S.; Domenech, P.; Honoré, N.; Song, J.; Deretic, V.; Cole, S.T. Regulation of catalase-peroxidase (KatG) expression, isoniazid sensitivity and virulence by furA of Mycobacterium tuberculosis. Mol. Microbiol. 2001, 40, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.Y.; Wang, Q.; Yang, Z.; Siame, B.A. Edwardsiella piscicida: A versatile emerging pathogen of fish. Virulence 2019, 10, 555–567. [Google Scholar] [CrossRef]
- Buján, N.; Toranzo, A.E.; Magariños, B. Edwardsiella piscicida: A significant bacterial pathogen of cultured fish. Dis. Aquat. Organ. 2018, 131, 59–71. [Google Scholar] [CrossRef]
- Wang, F.; Cheng, S.; Sun, K.; Sun, L. Molecular analysis of the fur (ferric uptake regulator) gene of a pathogenic Edwardsiella tarda strain. J. Microbiol. 2008, 46, 350–355. [Google Scholar] [CrossRef]
- Coy, M.; Doyle, C.; Besser, J.; Neilands, J.B. Site-directed mutagenesis of the ferric uptake regulation gene of Escherichia coli. Biometals 1994, 7, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.H.; Sun, L. The global regulatory effect of Edwardsiella tarda Fur on iron acquisition, stress resistance, and host infection: A proteomics-based interpretation. J. Proteom. 2016, 140, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Sivaraman, J.; Leung, K.Y.; Mok, Y.K. Two-component PhoB-PhoR Regulatory System and Ferric Uptake Regulator Sense Phosphate and Iron to Control Virulence Genes in Type III and VI Secretion Systems of Edwardsiella tarda. J. Biol. Chem. 2011, 286, 39417–39430. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.Y.; Huang, H.Q.; Li, S.; Tang, P.; Dai, H.F.; Xian, J.A.; Sun, D.M.; Hu, Y.H. Thioredoxin H (TrxH) contributes to adversity adaptation and pathogenicity of Edwardsiella piscicida. Vet. Res. 2019, 50, 26. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Wang, H.L.; Zhang, M.; Xiao, Z.Z.; Sun, L. Genetic mechanisms of multi-antimicrobial resistance in a pathogenic Edwardsiella tarda strain. Aquaculture 2009, 289, 134–139. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Hu, Y.H.; Chen, L.; Sun, L. CXCL8 of Scophthalmus maximus: Expression, biological activity and immunoregulatory effect. Dev. Comp. Immunol. 2011, 35, 1032–1039. [Google Scholar] [CrossRef]
- Li, H.; Sun, B.; Ning, X.; Jiang, S.; Sun, L. A Comparative Analysis of Edwardsiella tarda-Induced Transcriptome Profiles in RAW264.7 Cells Reveals New Insights into the Strategy of Bacterial Immune Evasion. Int. J. Mol. Sci. 2019, 20, 5724. [Google Scholar] [CrossRef]
- Troxell, B.; Hassan, H.M. Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria. Front. Cell. Infect. Microbiol. 2013, 3, 59. [Google Scholar] [CrossRef]
- Santander, J.; Golden, G.; Wanda, S.Y.; Curtiss, R. Fur-regulated iron uptake system of Edwardsiella ictaluri and its influence on pathogenesis and immunogenicity in the catfish host. Infect. Immun. 2012, 80, 2689–2703. [Google Scholar] [CrossRef]
- Pohl, E.; Haller, J.C.; Mijovilovich, A.; Meyer-Klaucke, W.; Garman, E.; Vasil, M.L. Architecture of a protein central to iron homeostasis: Crystal structure and spectroscopic analysis of the ferric uptake regulator. Mol. Microbiol. 2003, 47, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Ebanks, R.O.; Goguen, M.; Knickle, L.; Dacanay, A.; Leslie, A.; Ross, N.W.; Pinto, D.M. Analysis of a ferric uptake regulator (Fur) knockout mutant in Aeromonas salmonicida subsp. Salmonicida. Vet. Microbiol. 2013, 162, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, M.J.; Ingham, E.; Foster, S.J. In Staphylococcus aureus, Fur Is an Interactive Regulator with PerR, Contributes to Virulence, and Is Necessary for Oxidative Stress Resistance through Positive Regulation of Catalase and Iron Homeostasis. J. Bacteriol. 2001, 183, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Vasileva, D.; Janssen, H.; Hönicke, D.; Ehrenreich, A.; Bahl, H. Effect of iron limitation and fur gene inactivation on the transcriptional profile of the strict anaerobe Clostridium acetobutylicum. Microbiology 2012, 158, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Prince, R.W.; Cox, C.D.; Vasil, M.L. Coordinate regulation of siderophore and exotoxin A production: Molecular cloning and sequencing of the Pseudomonas aeruginosa fur gene. J. Bacteriol. 1993, 175, 2589–2598. [Google Scholar] [CrossRef] [PubMed]
- Neilands, J.B. Siderophores. Arch. Biochem. Biophys. 1993, 302, 1–3. [Google Scholar] [CrossRef]
- Zhou, Z.J.; Zhang, L.; Sun, L. Pseudomonas fluorescens: Fur is required for multiple biological properties associated with pathogenesis. Vet. Microbiol. 2015, 175, 145–149. [Google Scholar] [CrossRef]
- Allard, K.A.; Viswanathan, V.K.; Cianciotto, N.P. lbtA and lbtB Are Required for Production of the Legionella pneumophila Siderophore Legiobactin. J. Bacteriol. 2006, 188, 1351–1363. [Google Scholar] [CrossRef]
- Baichoo, N.; Wang, T.; Ye, R.; Helmann, J.D. Global analysis of the Bacillus subtilis Fur regulon and the iron starvation stimulon. Mol. Microbiol. 2002, 45, 1613–1629. [Google Scholar] [CrossRef]
- Thompson, D.K.; Beliaev, A.S.; Giometti, C.S.; Tollaksen, S.L.; Khare, T.; Lies, D.P.; Nealson, K.H.; Lim, H.; Yates, J., 3rd; Brandt, C.C.; et al. Transcriptional and Proteomic Analysis of a Ferric Uptake Regulator (Fur) Mutant of Shewanella oneidensis: Possible Involvement of Fur in Energy Metabolism, Transcriptional Regulation, and Oxidative Stress. Appl. Environ. Microbiol. 2002, 68, 881–892. [Google Scholar] [CrossRef]
- Platero, R.; Peixoto, L.; O’Brian, M.R.; Fabiano, E. Fur Is Involved in Manganese-Dependent Regulation of mntA (sitA) Expression in Sinorhizobium meliloti. Appl. Environ. Microbiol. 2004, 70, 4349–4355. [Google Scholar] [CrossRef] [PubMed]
- Becerra, G.; Merchán, F.; Blasco, R.; Igeño, M.I. Characterization of a ferric uptake regulator (Fur)-mutant of the cyanotrophic bacterium Pseudomonas pseudoalcaligenes CECT5344. J. Biotechnol. 2014, 190, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Smid, E.J.; Boeren, S.; Notebaart, R.A.; Abee, T. Bacterial Microcompartment-Dependent 1,2-Propanediol Utilization Stimulates Anaerobic Growth of Listeria monocytogenes EGDe. Front. Microbiol. 2019, 10, 2660. [Google Scholar] [CrossRef] [PubMed]
- Cortes, B.W.; Naditz, A.L.; Anast, J.M.; Schmitz-Esser, S. Transcriptome Sequencing of Listeria monocytogenes Reveals Major Gene Expression Changes in Response to Lactic Acid Stress Exposure but a Less Pronounced Response to Oxidative Stress. Front. Microbiol. 2020, 10, 3110. [Google Scholar] [CrossRef]
- Bijlsma, J.J.; Waidner, B.; Vliet, A.H.; Hughes, N.J.; Häg, S.; Bereswill, S.; Kelly, D.J.; Vandenbroucke-Grauls, C.M.; Kist, M.; Kusters, J.G. The Helicobacter pylori Homologue of the Ferric Uptake Regulator Is Involved in Acid Resistance. Infect. Immun. 2002, 70, 606–611. [Google Scholar] [CrossRef]
- Perry, R.D.; Craig, S.K.; Abney, J.; Bobrov, A.G.; Kirillina, O.; Mier, I.; Truszczynska, H.; Fetherston, J.D. Manganese transporters Yfe and MntH are Fur-regulated and important for the virulence of Yersinia pestis. Microbiology 2012, 158, 804–815. [Google Scholar] [CrossRef]
- Liu, J.; Tian, Y.; Zhao, Y.; Zeng, R.; Chen, B.; Hu, B.; Walcott, R.R. Ferric Uptake Regulator (FurA) is Required for Acidovorax citrulli Virulence on Watermelon. Phytopathology 2019, 109, 1997–2008. [Google Scholar] [CrossRef]
- Srinivasa Rao, P.S.; Lim, T.M.; Leung, K.Y. Opsonized Virulent Edwardsiella tarda Strains Are Able to Adhere to and Survive and Replicate within Fish Phagocytes but Fail to Stimulate Reactive Oxygen Intermediates. Infect. Immun. 2001, 69, 5689–5697. [Google Scholar] [CrossRef]
- Zhang, L.; Ni, C.; Xu, W.; Dai, T.; Yang, D.; Wang, Q.; Zhang, Y.; Liu, Q. Intramacrophage Infection Reinforces the Virulence of Edwardsiella tarda. J. Bacteriol. 2016, 198, 1534–1542. [Google Scholar] [CrossRef]
- Chen, H.; Yang, D.; Han, F.; Tan, J.; Zhang, L.; Xiao, J.; Zhang, Y.; Liu, Q. The Bacterial T6SS Effector EvpP Prevents NLRP3 Inflammasome Activation by Inhibiting the Ca2+-Dependent MAPK-Jnk Pathway. Cell Host Microbe 2017, 21, 47–58. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′→3′) # |
|---|---|
| fur2-up-F | AGCGGGATCCGACGCCTGGGTCAAGCAGC |
| fur2-up-R | GTATACCGCGGTGTTGTTGTCAGTCA |
| fur2-down-F | ACAACAACACCGCGGTATACCGCGTGCTCAACCAG |
| fur2-down-R | AGCGGGATCCTTAGGCCTTTTCGTCGTGCA |
| fur2-test-F | AGGACAGAATCCGAATGAC |
| fur2-test-R | TTCACGCTGACGCTTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, M.; Sui, Z.; Jiang, G.; Wang, L.; Yao, X.; Hu, Y. The Mutation of the DNA-Binding Domain of Fur Protein Enhances the Pathogenicity of Edwardsiella piscicida via Inducing Overpowering Pyroptosis. Microorganisms 2024, 12, 11. https://doi.org/10.3390/microorganisms12010011
Niu M, Sui Z, Jiang G, Wang L, Yao X, Hu Y. The Mutation of the DNA-Binding Domain of Fur Protein Enhances the Pathogenicity of Edwardsiella piscicida via Inducing Overpowering Pyroptosis. Microorganisms. 2024; 12(1):11. https://doi.org/10.3390/microorganisms12010011
Chicago/Turabian StyleNiu, Mimi, Zhihai Sui, Guoquan Jiang, Ling Wang, Xuemei Yao, and Yonghua Hu. 2024. "The Mutation of the DNA-Binding Domain of Fur Protein Enhances the Pathogenicity of Edwardsiella piscicida via Inducing Overpowering Pyroptosis" Microorganisms 12, no. 1: 11. https://doi.org/10.3390/microorganisms12010011