
Discovery of Antimicrobial Peptides That Can Accelerate Culture Diagnostics of Slow-Growing Mycobacteria Including Mycobacterium tuberculosis

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Peptides
2.2. Mycobacterial Culture
2.3. Peptide Screening
2.4. Quantitative PCR
3. Results
3.1. Initial Screen against Mycobacterium avium subsp. paratuberculosis (MAP)
3.2. Test of Growth-Enhancing Activity in Other Mycobacterial Species and Inoculum Effect
3.3. Peptides Stabilized against Protease Cleavage
3.4. Peptides Tested on Mycobacterium Tuberculosis
3.5. Testing Naturally Occurring Antimicrobial Peptides against M. tuberculosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cox, R.A. Quantitative relationships for specific growth rates and macromolecular compositions of Mycobacterium tuberculosis, Streptomyces coelicolor A3(2) and Escherichia coli B/r: An integrative theoretical approach. Microbiology 2004, 150, 1413–1426. [Google Scholar] [CrossRef]
- CDC. Data and Statistics on TB. Available online: https://www.cdc.gov/tb/statistics/default.htm (accessed on 1 November 2018).
- WHO. Tuberculosis—Key Facts. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 1 November 2018).
- Stevenson, K.; Alvarez, J.; Bakker, D.; Biet, F.; de Juan, L.; Denham, S.; Dimareli, Z.; Dohmann, K.; Gerlach, G.F.; Heron, I.; et al. Occurrence of Mycobacterium avium subspecies paratuberculosis across host species and European countries with evidence for transmission between wildlife and domestic ruminants. BMC Microbiol. 2009, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Patrzykat, A. Clinical development of cationic antimicrobial peptides: From natural to novel antibiotics. Curr. Drug Targets Infect. Disord. 2002, 2, 79–83. [Google Scholar] [CrossRef]
- Hancock, R.E. Cationic peptides: Effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 2001, 1, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Mania, D.; Hilpert, K.; Ruden, S.; Fischer, R.; Takeshita, N. Screening for antifungal peptides and their modes of action in Aspergillus nidulans. Appl. Environ. Microbiol. 2010, 76, 7102–7108. [Google Scholar] [CrossRef]
- Cole, A.M. Minidefensins and other antimicrobial peptides: Candidate anti-HIV microbicides. Expert Opin. Ther. Targets 2003, 7, 329–341. [Google Scholar] [CrossRef]
- Silva, J.P.; Appelberg, R.; Gama, F.M. Antimicrobial peptides as novel anti-tuberculosis therapeutics. Biotechnol. Adv. 2016, 34, 924–940. [Google Scholar] [CrossRef]
- Ramón-García, S.; Mikut, R.; Ng, C.; Ruden, S.; Volkmer, R.; Reischl, M.; Hilpert, K.; Thompson, C.J. Targeting mycobacterium tuberculosis and other microbial pathogens using improved synthetic antibacterial peptides. Antimicrob. Agents Chemother. 2013, 57, 2295–2303. [Google Scholar] [CrossRef]
- Nuti, R.; Goud, N.S.; Saraswati, A.P.; Alvala, R.; Alvala, M. Antimicrobial Peptides: A Promising Therapeutic Strategy in Tackling Antimicrobial Resistance. Curr. Med. Chem. 2017, 24, 4303–4314. [Google Scholar] [CrossRef]
- Territo, M.C.; Ganz, T.; Selsted, M.E.; Lehrer, R. Monocyte-chemotactic activity of defensins from human neutrophils. J. Clin. Investig. 1989, 84, 2017–2020. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Iwabuchi, K.; Matsuda, H.; Ogawa, H.; Nagaoka, I. Epithelial cell-derived human beta-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase C-dependent pathway. Int. Immunol. 2002, 14, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Elssner, A.; Duncan, M.; Gavrilin, M.; Wewers, M.D. A novel P2X7 receptor activator, the human cathelicidin-derived peptide LL37, induces IL-1 beta processing and release. J. Immunol. 2004, 172, 4987–4994. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, A.; Braff, M.H.; Taylor, K.R.; Na, C.; Granstein, R.D.; McInturff, J.E.; Krutzik, S.; Modlin, R.L.; Gallo, R.L. Cathelicidin antimicrobial peptides block dendritic cell TLR4 activation and allergic contact sensitization. J. Immunol. 2007, 178, 1829–1834. [Google Scholar] [CrossRef]
- Heilborn, J.D.; Nilsson, M.F.; Sørensen, O.; Ståhle-Bäckdahl, M.; Kratz, G.; Weber, G.; Borregaard, N. The Cathelicidin Anti-Microbial Peptide LL-37 is Involved in Re-Epithelialization of Human Skin Wounds and is Lacking in Chronic Ulcer Epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Carretero, M.; Escámez, M.J.; García, M.; Duarte, B.; Holguín, A.; Retamosa, L.; Jorcano, J.L.; del Río, M.; Larcher, F. In vitro and In vivo Wound Healing-Promoting Activities of Human Cathelicidin LL-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef]
- Hilpert, K.; McLeod, B.; Yu, J.; Elliott, M.R.; Rautenbach, M.; Ruden, S.; Bürck, J.; Muhle-Goll, C.; Ulrich, A.S.; Keller, S.; et al. Short cationic antimicrobial peptides interact with ATP. Antimicrob. Agents Chemother. 2010, 54, 4480–4483. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Cho, J.H.; Sung, B.H.; Kim, S.C. Buforins: Histone H2A-derived antimicrobial peptides from toad stomach. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1564–1569. [Google Scholar] [CrossRef]
- Kobayashi, S.; Takeshima, K.; Park, C.B.; Kim, S.C.; Matsuzaki, K. Interactions of the novel anfimicrobial peptide buforin 2 with lipid bilayers: Proline as a translocation promoting factor. Biochemistry 2000, 39, 8648–8654. [Google Scholar] [CrossRef]
- Marchand, C.; Krajewski, K.; Lee, H.-F.; Antony, S.; Johnson, A.A.; Amin, R.; Roller, P.; Kvaratskhelia, M.; Pommier, Y. Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res. 2006, 34, 5157–5165. [Google Scholar] [CrossRef]
- Carlsson, A.; Engström, P.; Palva, E.T.; Bennich, H. Attacin, an antibacterial protein from Hyalophora cecropia, inhibits synthesis of outer membrane proteins in Escherichia coli by interfering with omp gene transcription. Infect. Immun. 1991, 59, 3040–3045. [Google Scholar] [CrossRef]
- Mardirossian, M.; Sola, R.; Beckert, B.; Collis, D.W.P.; Di Stasi, A.; Armas, F.; Hilpert, K.; Wilson, D.N.; Scocchi, M. Proline-rich peptides with improved antimicrobial activity against E. coli, K. pneumoniae and A. baumannii. ChemMedChem 2019, 14, 2025–2033. [Google Scholar] [CrossRef]
- Mardirossian, M.; Sola, R.; Beckert, B.; Valencic, E.; Collis, D.W.P.; Borišek, J.; Armas, F.; Di Stasi, A.; Buchmann, J.; Syroegin, E.A.; et al. Peptide Inhibitors of Bacterial Protein Synthesis with Broad Spectrum and SbmA-Independent Bactericidal Activity against Clinical Pathogens. J. Med. Chem. 2020, 63, 9590–9602. [Google Scholar] [CrossRef]
- Mardirossian, M.; Pérébaskine, N.; Benincasa, M.; Gambato, S.; Hofmann, S.; Huter, P.; Müller, C.; Hilpert, K.; Innis, C.A.; Tossi, A.; et al. The Dolphin Proline-Rich Antimicrobial Peptide Tur1A Inhibits Protein Synthesis by Targeting the Bacterial Ribosome. Cell Chem. Biol. 2018, 25, 530.e7–539.e7. [Google Scholar] [CrossRef]
- Iavicoli, I.; Fontana, L.; Agathokleous, E.; Santocono, C.; Russo, F.; Vetrani, I.; Fedele, M.; Calabrese, E.J. Hormetic dose responses induced by antibiotics in bacteria: A phantom menace to be thoroughly evaluated to address the environmental risk and tackle the antibiotic resistance phenomenon. Sci. Total Environ. 2021, 798, 149255. [Google Scholar] [CrossRef]
- Gorr, S.U.; Brigman, H.V.; Anderson, J.C.; Hirsch, E.B. The antimicrobial peptide DGL13K is active against drug-resistant gram-negative bacteria and sub-inhibitory concentrations stimulate bacterial growth without causing resistance. PLoS ONE 2022, 17, e0273504. [Google Scholar] [CrossRef] [PubMed]
- Nema, V. Tuberculosis diagnostics: Challenges and opportunities. Lung India 2012, 29, 259. [Google Scholar] [CrossRef] [PubMed]
- Luelmo, F. What is the role of sputum microscopy in patients attending health facilities. In Toman’s Tuberculosis: Case Detection, Treatment, and Monitoring—Questions and Answers; Frieden, T., Ed.; World Health Organization: Geneva, Switzerland, 2004; pp. 7–10. [Google Scholar]
- Suleiman, K. An Activist’s Guide to Tuberculosis Diagnostic Tools; Lessem, E., Ed.; Treatment Action Group (TAG): New York, NY, USA, 2017. [Google Scholar]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E.W. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Jenssen, H.; Hilpert, K.; Cheung, W.A.; Panté, N.; Hancock, R.E.W.; Cherkasov, A. Identification of novel antibacterial peptides by chemoinformatics and machine learning. J. Med. Chem. 2009, 52, 2006–2015. [Google Scholar] [CrossRef] [PubMed]
- Knappe, D.; Ruden, S.; Langanke, S.; Tikkoo, T.; Ritzer, J.; Mikut, R.; Martin, L.L.L.; Hoffmann, R.; Hilpert, K. Optimization of oncocin for antibacterial activity using a SPOT synthesis approach: Extending the pathogen spectrum to Staphylococcus aureus. Amino Acids 2016, 48, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Knappe, D.; Kabankov, N.; Hoffmann, R. Bactericidal oncocin derivatives with superior serum stabilities. Int. J. Antimicrob. Agents 2011, 37, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, F.C.; Budnick, G.E.; Elliott, T.; Gillim-Ross, L.; Hildred, M.V.; Mahlmeister, P.; Parrish, N.; Pentella, M.; Vanneste, J.; Wang, Y.F.; et al. Probability of negative Mycobacterium tuberculosis complex cultures based on time to detection of positive cultures: A multicenter evaluation of commercial-broth-based culture systems. J. Clin. Microbiol. 2012, 50, 3275–3282. [Google Scholar] [CrossRef]
- Barlow, P.G.; Findlay, E.G.; Currie, S.M.; Davidson, D.J. Antiviral potential of cathelicidins. Future Microbiol. 2014, 9, 55–73. [Google Scholar] [CrossRef]
- Kokryakov, V.N.; Harwig, S.S.; Panyutich, E.A.; Shevchenko, A.A.; Aleshina, G.M.; Shamova, O.V.; Korneva, H.A.; Lehrer, R.I. Protegrins: Leukocyte antimicrobial peptides that combine features of corticostatic defensins and tachyplesins. FEBS Lett. 1993, 327, 231–236. [Google Scholar] [CrossRef]
- Čeřovský, V.; Buděšínský, M.; Hovorka, O.; Cvačka, J.; Voburka, Z.; Slaninová, J.; Borovičková, L.; Fučík, V.; Bednárová, L.; Votruba, I.; et al. Lasioglossins: Three Novel Antimicrobial Peptides from the Venom of the Eusocial Bee Lasioglossum laticeps (Hymenoptera: Halictidae). ChemBioChem 2009, 10, 2089–2099. [Google Scholar] [CrossRef]
- Battista, F.; Oliva, R.; del Vecchio, P.; Winter, R.; Petraccone, L. Insights into the Action Mechanism of the Antimicrobial Peptide Lasioglossin III. Int. J. Mol. Sci. 2021, 22, 2857. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Mignogna, G.; Andreu, D.; Shai, Y.; Barra, D.; Simmaco, M. Ranacyclins, a New Family of Short Cyclic Antimicrobial Peptides: Biological Function, Mode of Action, and Parameters Involved in Target Specificity. Biochemistry 2003, 42, 14023–14035. [Google Scholar] [CrossRef]
- He, M.; Zhang, H.; Li, Y.; Wang, G.; Tang, B.; Zhao, J.; Huang, Y.; Zheng, J. Cathelicidin-Derived Antimicrobial Peptides Inhibit Zika Virus Through Direct Inactivation and Interferon Pathway. Front. Immunol. 2018, 9, 722. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Han, H.; Miller, D.W.; Wang, G. Solution Structures of Human LL-37 Fragments and NMR-Based Identification of a Minimal Membrane-Targeting Antimicrobial and Anticancer Region. J. Am. Chem. Soc. 2006, 128, 5776–5785. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Bacterial sensing of antimicrobial peptides. Contrib. Microbiol. 2009, 16, 136–149. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
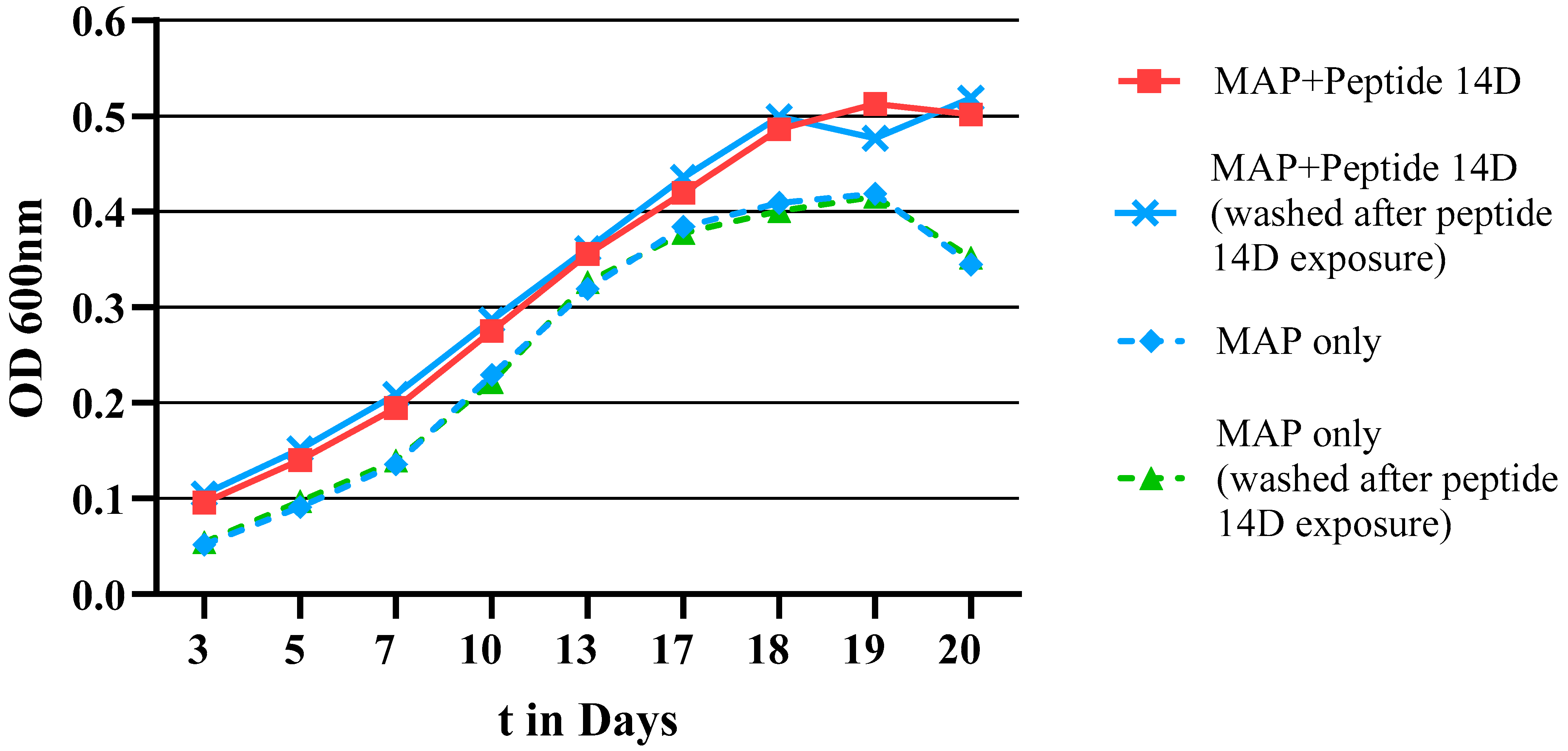{kind=link}
{kind=link}
{kind=link}
{kind=link}
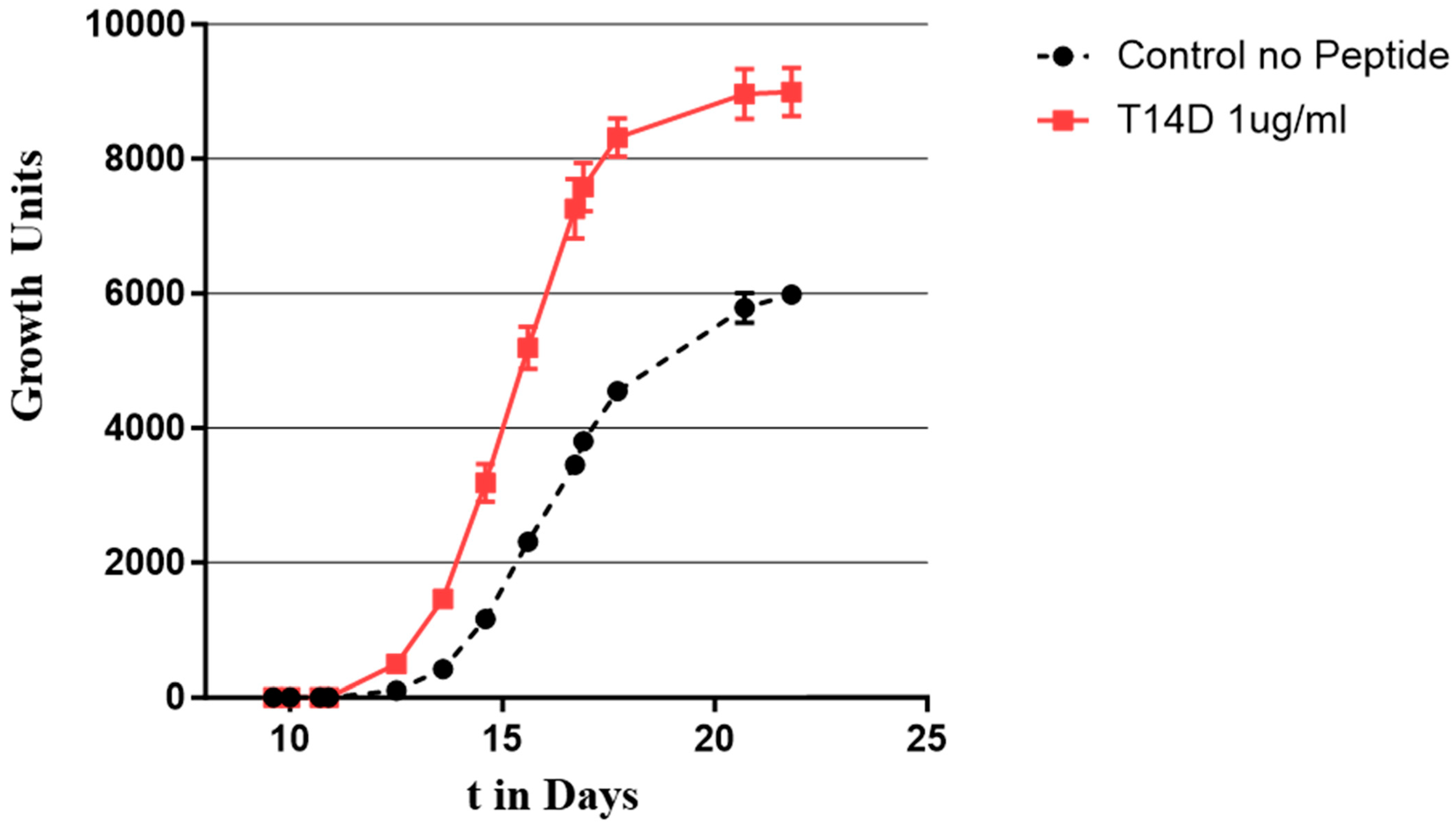{kind=link}
{kind=link}
| Peptide Code | Sequence | Time to Positivity (Days.Hours) Experiment 1a | Time to Positivity (Days.Hours) Experiment 1b | Time to Positivity (Days.Hours) Experiment 2a | Time to Positivity (Days.Hours) Experiment 2b | Average % Faster than Control |
|---|---|---|---|---|---|---|
| TB control (no peptide) | 7.02 | 7.01 | 15.01 | 15.20 | ||
| 59 | ILKWKRKWWKWRR | 5.17 | 5.22 | 13.19 | 12.22 | 14.5 |
| 69 | RRWRIVVIRVRR | 6.02 | 6.02 | 13.12 | 13.02 | 12.8 |
| 61 | ILKWKTKWWKWFR | 6.20 | 5.23 | 13.11 | 14.21 | 10.4 |
| 12 | RRRIKIRWY | 6.14 | 5.20 | 14.08 | 14.22 | 10.2 |
| 71 | RLWRIVVIRVKR | 6.04 | 6.06 | 14.01 | 14.02 | 9.4 |
| 14 | WKIVFWWRR | 6.01 | 6.12 | 13.23 | 14.01 | 9.1 |
| 5 | FFIYVWRRR | 5.21 | 6.08 | 14.06 | 14.16 | 8.8 |
| 16 | RLKRWWKFL | 6.01 | 5.22 | 14.17 | 14.20 | 8.6 |
| 20 | KWKWWWRKI | 6.11 | 6.00 | 14.01 | 14.16 | 8.3 |
| 60 | ILKWKIFKWKWFR | 6.03 | 6.02 | 14.19 | 14.16 | 7.9 |
| 49 | VRLRIRVRVIRK | 5.22 | 6.06 | 15.12 | 15.02 | 6.2 |
| 9 | KRRWRIWLV | 6.07 | 6.04 | 15.08 | 14.21 | 5.8 |
| 78 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | 6.08 | 6.09 | 14.15 | 15.00 | 5.8 |
| 77 | VRLRIRVAVIRA | 6.14 | 6.04 | 14.22 | 15.03 | 5.0 |
| 29 | RWRWWWRVY | 6.13 | 6.00 | 15.12 | 15.01 | 4.9 |
| 22 | IRMRIRVLL | 6.00 | 6.07 | 15.19 | 15.10 | 4.7 |
| 70 | RLARIVVIRVRR | 6.03 | 6.19 | 14.22 | 15.06 | 4.2 |
| 3 | WKWRVRVTI | 6.08 | 6.17 | 14.22 | 15.02 | 4.0 |
| 68 | RLRRIVVIRVAR | 6.09 | 6.17 | 15.00 | 15.23 | 2.3 |
| 65 | RGARIVVIRVAR | 6.16 | 6.03 | 16.11 | 16.01 | 0.8 |
| 72 | RLARIVVIRWAR | 6.22 | 7.01 | 15.01 | 15.13 | −0.2 |
| 67 | RLWRIVVIRVAR | 6.19 | 6.11 | 16.00 | 16.05 | −0.4 |
| 64 | RLARIVVIRVAR | 6.19 | 6.16 | 15.21 | 16.13 | −1.4 |
| 76 | VRWRIRVAVIRA | 6.13 | 6.16 | 16.08 | 16.14 | −1.4 |
| 62 | ILKWKMFKWKWFR | 6.07 | 7.02 | 16.20 | 17.16 | −3.4 |
| 66 | RRARIVVIRVAR | 6.05 | 6.16 | 17.06 | 17.16 | −3.5 |
| 63 | ILPWKWPWWPWRR | 6.22 | 6.22 | 17.01 | 17.10 | −5.5 |
| Peptide 14D Concentration (µg/mL) | Time to Positivity (Days.Hours) Experiment 1 | Time to Positivity (Days.Hours) Experiment 2 | Average % Faster than Control |
|---|---|---|---|
| 0 | 6.19 | 6.12 | |
| 0.1 | 5.12 | 5.19 | 15.0 |
| 0.5 | 5.17 | 5.17 | 14.1 |
| 1 | 5.05 | 5.08 | 21.6 |
| 10 | 7.03 | 7.08 | −7.3 |
| 25 | 9.21 | 9.17 | −48.7 |
| 50 | 19.15 | 21.5 | −207.6 |
| Name/Source | Sequence | Time to Positivity (Day.Hours) Experiment 1 | Time to Positivity (Day.Hours) Experiment 2 | Average % Faster than Control |
|---|---|---|---|---|
| Control (TB only) | 11.10 | 9.10 | ||
| Protegrin 1/Pig * | RGGRLCYCRRRFCVCV | 10.23 | 6.22 | 15.3 |
| Lasioglossin LL-III/Bee | VNWKKILGKIIKVVK | 9.18 | 8.08 | 13.1 |
| Ranacyclin E/Frog | SAPRGCWTKSYPPKPCK | 10.07 | 8.01 | 12.2 |
| GF-17/Human * | GFKRIVQRIKDFLRNLV | 10.02 | 8.07 | 11.8 |
| Arenicin 1/Annelid worm * | RWCVYAYVRVRGVLVR | 10.15 | 8.02 | 10.5 |
| PP13/Wasp * | IRLHRLYTWKATIYTR | 10.17 | 8.02 | 10.2 |
| Peptide B9/Frog | FLPLIAGLIGKLF | 10.19 | 8.01 | 10.0 |
| CM15/Bee venom | KWKLFKKIGAVLKVL | 10.19 | 8.01 | 10.0 |
| Macropin 1/Wasp | GFGMALKLLKKVL | 10.12 | 8.07 | 10.0 |
| Salusin-beta/Scorpion * | FIFIRWLLKLGHHGRA | 10.18 | 8.03 | 9.8 |
| GLK-19/Designed * | KKLLGKLLKKLGKLLL | 10.17 | 8.04 | 9.7 |
| Odorranain-H1/Frog * | FGKILGVGKKVLCGLS | 10.19 | 8.03 | 9.6 |
| Temporin-HN1/Frog | AILTTLANWARKFL | 10.23 | 8.01 | 9.3 |
| CPF-AM4/Frog | GLGSLVGNALRIGAKLL | 10.17 | 8.06 | 9.3 |
| Tachyplasin II/Crab | RWCFRVCYRGICYRKCR | 10.18 | 8.06 | 9.1 |
| Mastoparan-VT6/Wasp | INLKAIAALVKKLL | 10.21 | 8.04 | 9.0 |
| Temporin-1TSa/Frog | FLGALAKIISGIF | 11.00 | 8.02 | 8.9 |
| Temporin-1DRa/Frog | HFLGTLVNLAKKIL | 10.18 | 8.09 | 8.5 |
| Temporin-HN2/Frog | NILNTIINLAKKIL | 11.03 | 8.02 | 8.4 |
| NRC-15/Fish * | WGKLFKLGLHGIGLLH | 10.21 | 8.07 | 8.3 |
| CM-3/Designed | ALKAALLAILKIVRVI | 11.04 | 8.03 | 8.0 |
| Piscidin 1/Fish * | FFHHIFRGIVHVGKTI | 10.18 | 8.12 | 7.8 |
| Eumenitin/Wasp | LNLKGIFKKVASLLT | 11.00 | 8.08 | 7.6 |
| OdVP2/Wasp | ILGIITSLLKSLGKK | 10.22 | 8.11 | 7.3 |
| Clavanin C/Sea squirt | VFHLLGKIIHHVGNFV | 10.21 | 8.12 | 7.2 |
| P-04/Tunicate | FFPYAALKWLRKLLKK | 11.02 | 8.08 | 7.2 |
| Temporin L/Frog | FVQWFSKFLGRIL | 11.06 | 8.05 | 7.1 |
| Protegrin 4/Pig * | GRLCYCRGWICFCVGR | 11.11 | 8.01 | 7.1 |
| D28/Designed * | FLGVVFKLASKVFPAV | 11.02 | 8.09 | 7.0 |
| TsAP-1/Scorpion | FLSLIPSLVGGSISAFK | 11.07 | 8.05 | 7.0 |
| Brevinin-1Ta/Frog | FITLLLRKFICSITKKC | 11.07 | 8.05 | 7.0 |
| Bombolitin V/Bee | INVLGILGLLGKALSHL | 10.22 | 8.13 | 6.8 |
| Temporin-LTb/Frog | FIITGLVRGLTKLF | 11.07 | 8.06 | 6.7 |
| Temporin-1Cb/Frog | FLPLFASLIGKLL | 11.08 | 8.06 | 6.6 |
| Hylin a1/Frog | GAILPLALGALKNLIK | 11.14 | 8.02 | 6.3 |
| Arenicin-2/Lugworm | YAYVRIRGVLVRYRRC | 11.13 | 8.03 | 6.3 |
| Temporin-PTa/Frog | FFGSVLKLIPKIL | 11.07 | 8.08 | 6.3 |
| Decoralin/Wasp | SLLSLIRKLIT | 11.12 | 8.04 | 6.3 |
| Pd_mastoparan PDD-B/Wasp | INWLKLGKKILGAL | 11.06 | 8.09 | 6.3 |
| NRC-15/Flounder | GFWGKLFKLGLHGIGL | 11.02 | 8.14 | 5.9 |
| Clavanin E/Sea squirt | LFKLLGKIIHHVGNFV | 11.05 | 8.12 | 5.8 |
| Temporin 1Vb/Frog | FLSIIAKVLGSLF | 11.10 | 8.08 | 5.8 |
| Alyteserin-2b/Toad | ILGAILPLVSGLLSNKL | 11.18 | 8.02 | 5.6 |
| Polyphemusin/Crab * | WCFRVCYKGFCYRKCR | 11.05 | 8.13 | 5.6 |
| IsCT2/Scorpion | IFGAIWNGIKSLF | 10.23 | 8.21 | 4.9 |
| Indolicidin/Cow | ILPWKWPWWPWRR | 11.03 | 8.18 | 4.8 |
| Temporin-SN1/Frog | FFPFLLGALGSLLPKIF | 10.22 | 8.23 | 4.6 |
| Pelophylaxin-4/Frog | ILPFLAGLFSKIL | 11.05 | 8.18 | 4.5 |
| P9/Designed | KRRWRIWLV | 11.10 | 8.14 | 4.4 |
| Brevinin 1Tb/Frog | LVPLFLSKLICFITKKC | 11.10 | 8.15 | 4.2 |
| Casecidin 17/Cow | YQEPVLGPVRGPFPIIV | 11.12 | 8.14 | 4.1 |
| Brevinin-1DYc/Frog | LLAGLPKLLCLFFKKC | 11.10 | 8.16 | 4.0 |
| Temporin-1Oc/Frog | FLPLLASLFSRLF | 11.10 | 8.17 | 3.8 |
| Temporin A/Frog | FLPLIGRVLSGIL | 11.02 | 9.00 | 3.7 |
| Fallaxidin/Frog * | LSFLPKVIGVIGHLIH | 11.18 | 8.13 | 3.2 |
| Pantinin-3/Scorpion | FLSTIWNGIKSLL | 11.08 | 8.23 | 2.8 |
| Temporin-1Vc/Frog | FLPLVTMLLGKLF | 11.12 | 8.21 | 2.5 |
| P8/Designed * | FFIYVWRRR | 11.16 | 8.22 | 1.6 |
| Tritrpticin/Pig | VRRFPWWWPFLRR | 11.08 | 9.13 | −0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilpert, K.; Munshi, T.; López-Pérez, P.M.; Sequeira-Garcia, J.; Hofmann, S.; Bull, T.J. Discovery of Antimicrobial Peptides That Can Accelerate Culture Diagnostics of Slow-Growing Mycobacteria Including Mycobacterium tuberculosis. Microorganisms 2023, 11, 2225. https://doi.org/10.3390/microorganisms11092225
Hilpert K, Munshi T, López-Pérez PM, Sequeira-Garcia J, Hofmann S, Bull TJ. Discovery of Antimicrobial Peptides That Can Accelerate Culture Diagnostics of Slow-Growing Mycobacteria Including Mycobacterium tuberculosis. Microorganisms. 2023; 11(9):2225. https://doi.org/10.3390/microorganisms11092225
Chicago/Turabian StyleHilpert, Kai, Tulika Munshi, Paula M. López-Pérez, Joana Sequeira-Garcia, Sven Hofmann, and Tim J. Bull. 2023. "Discovery of Antimicrobial Peptides That Can Accelerate Culture Diagnostics of Slow-Growing Mycobacteria Including Mycobacterium tuberculosis" Microorganisms 11, no. 9: 2225. https://doi.org/10.3390/microorganisms11092225