
Evaluation of Different Activity of Lactobacillus spp. against Two Proteus mirabilis Isolated Clinical Strains in Different Anatomical Sites In Vitro: An Explorative Study to Improve the Therapeutic Approach
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Biofilm Formation Assay
2.3. Counting of Sessile Bacteria
2.4. Cells Cultures and Treatment
2.5. Analysis of Flagellar Gene Expression
2.6. Real-Time PCR
2.7. Bacterial Internalization Assay
2.8. ELISA Assay
2.9. Statistical Analysis
3. Results
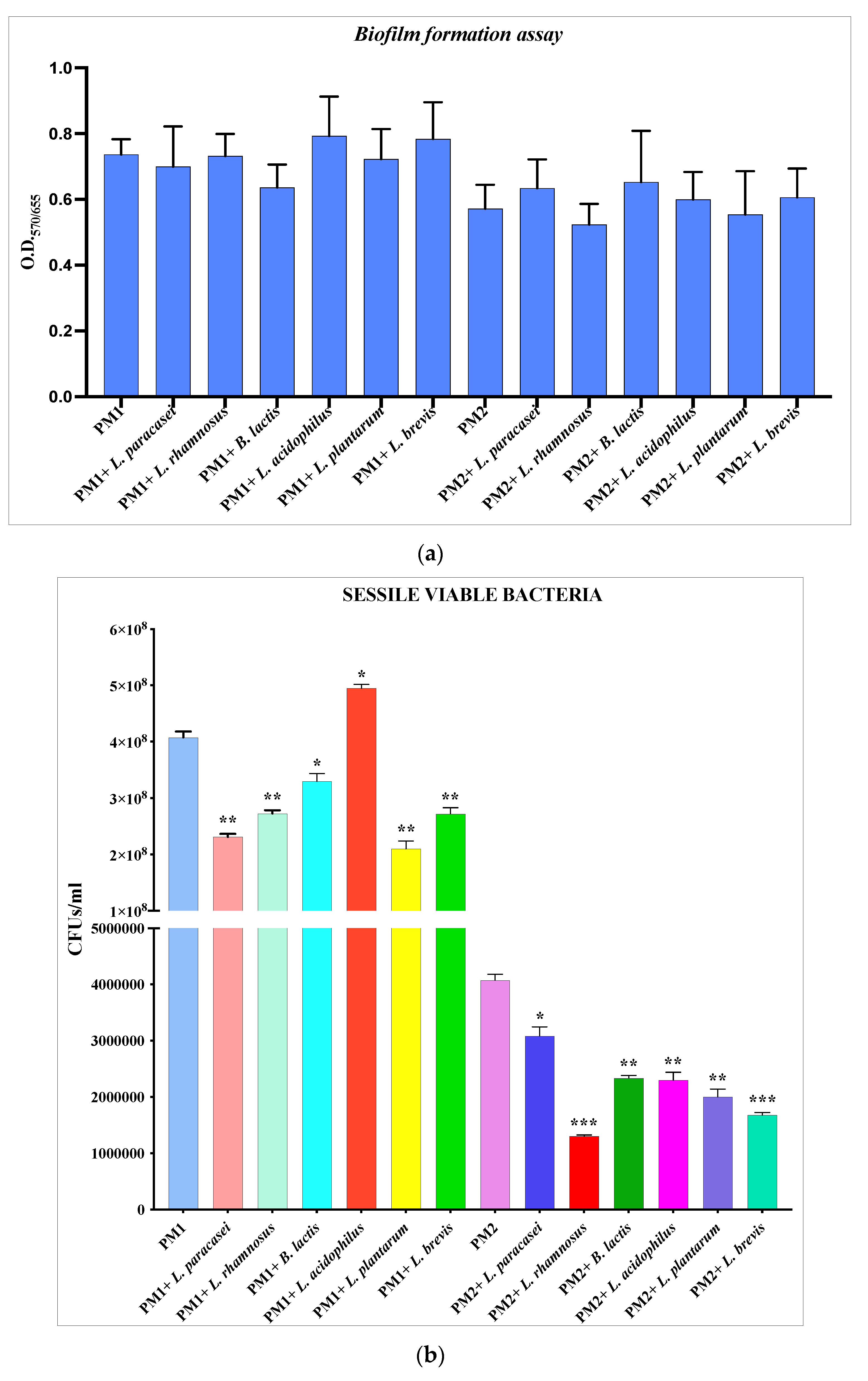
3.1. Antibiofilm activity of Lactobacillus spp. against P. mirabilis
3.2. Selection of Best Lactobacillus spp. Strain for Each Epithelium
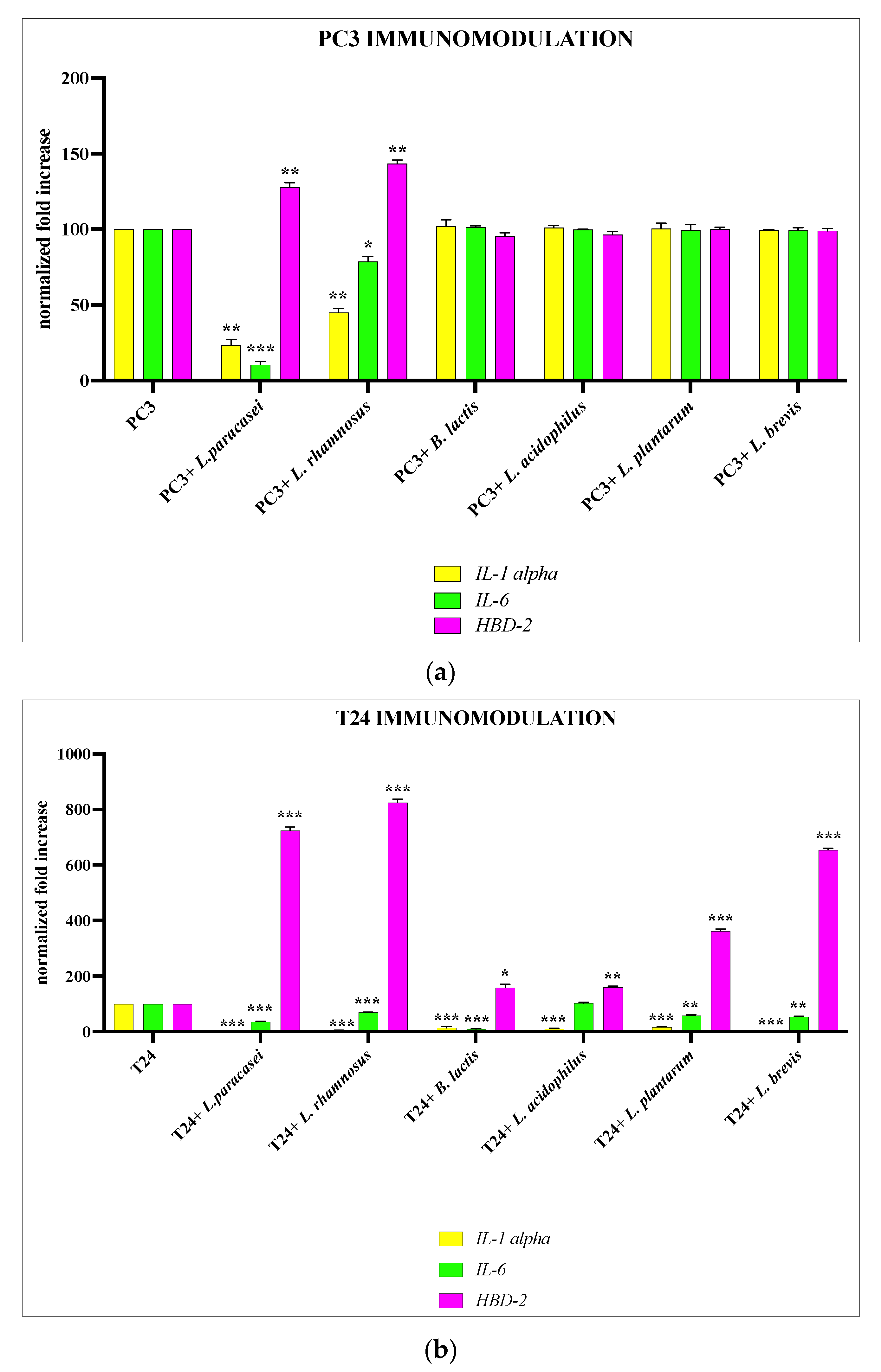
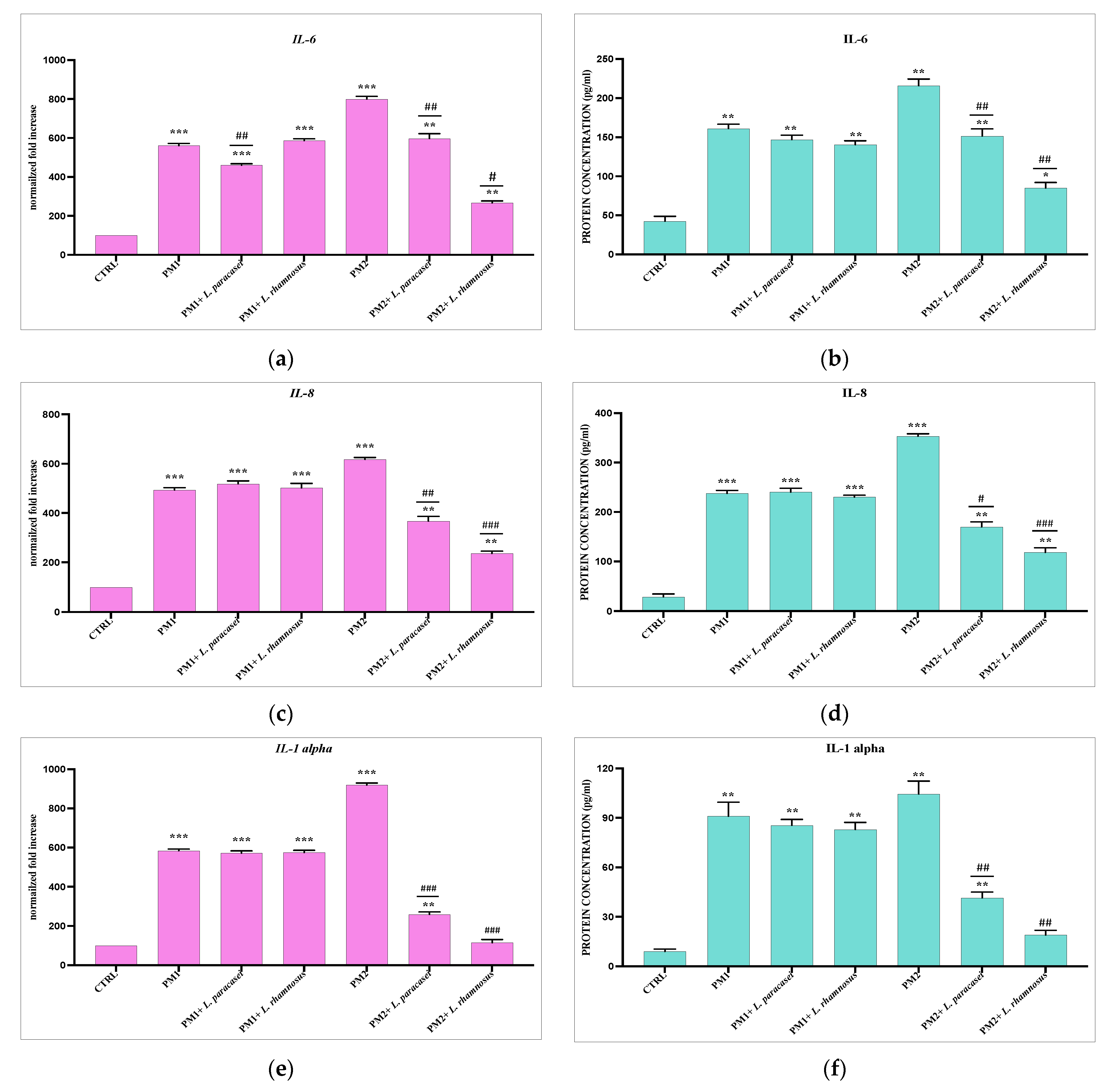
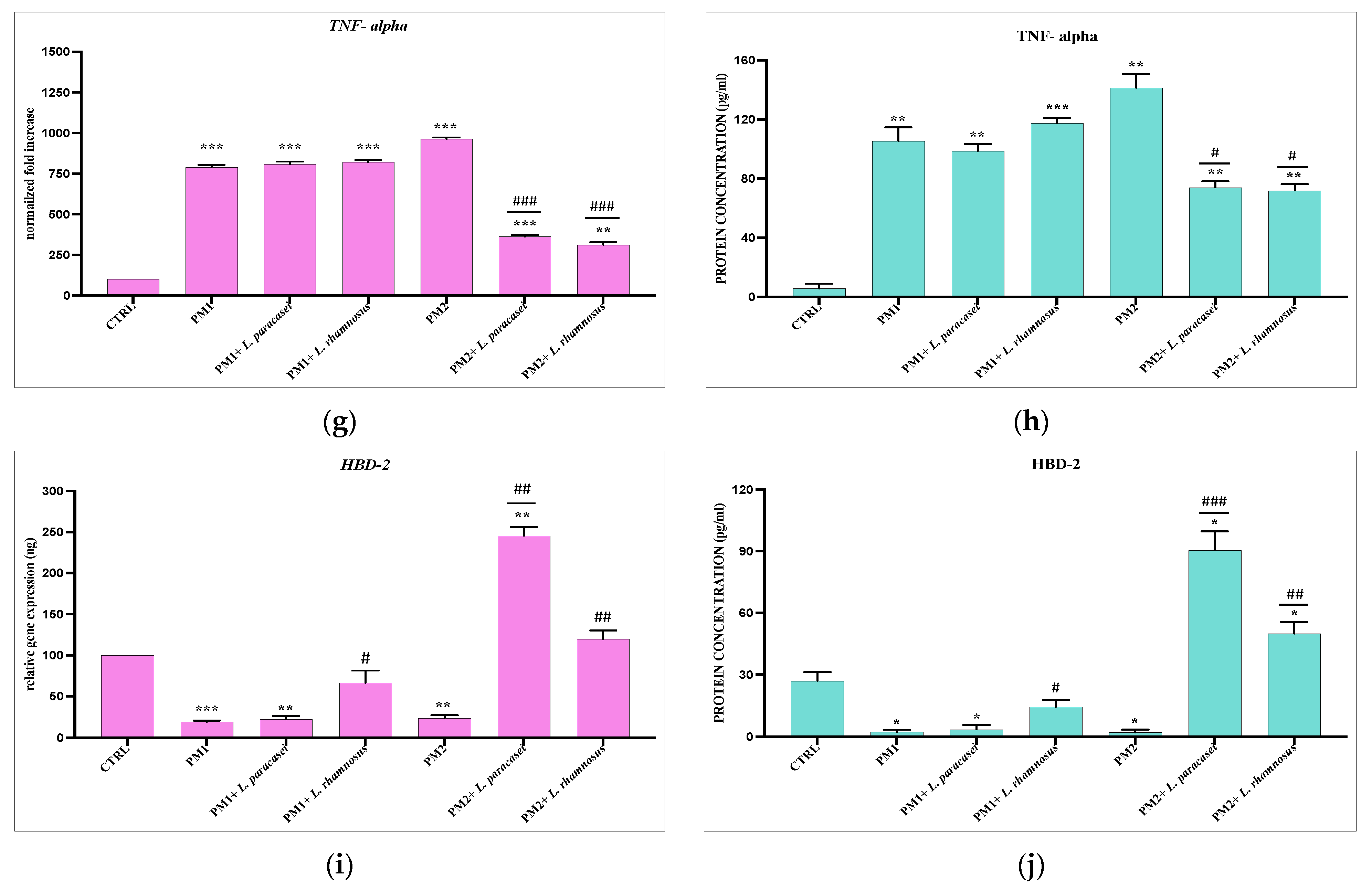
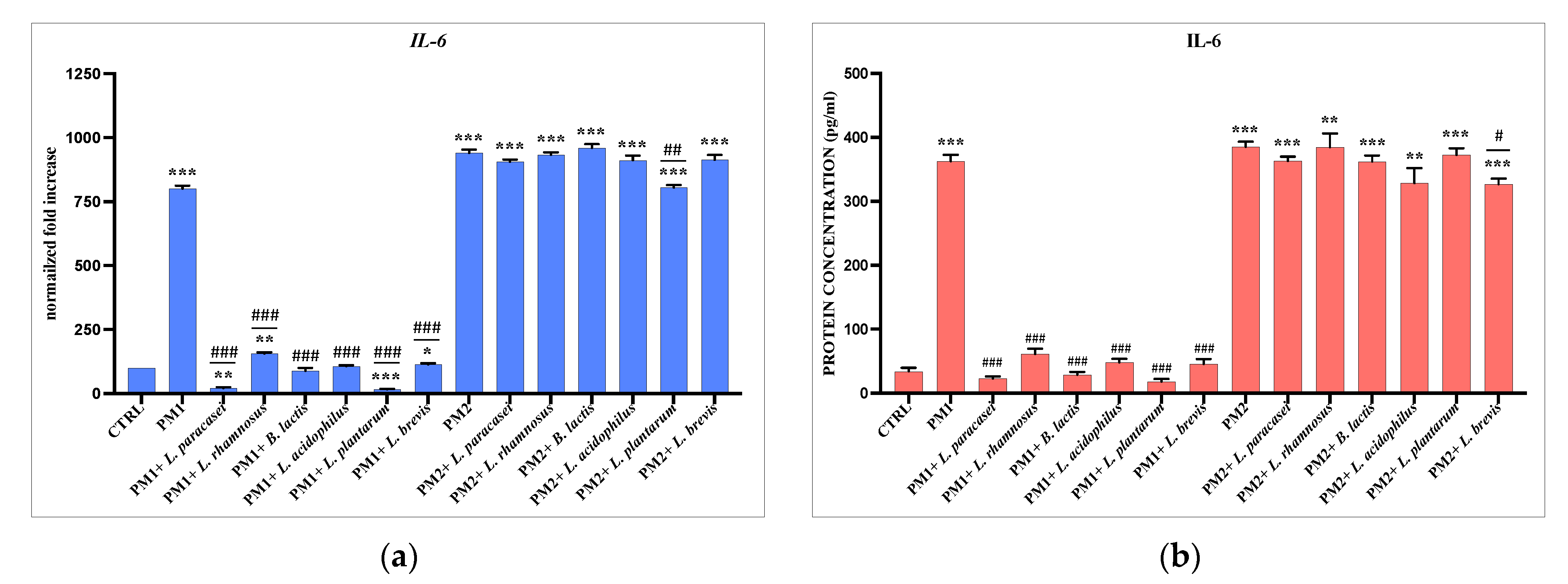
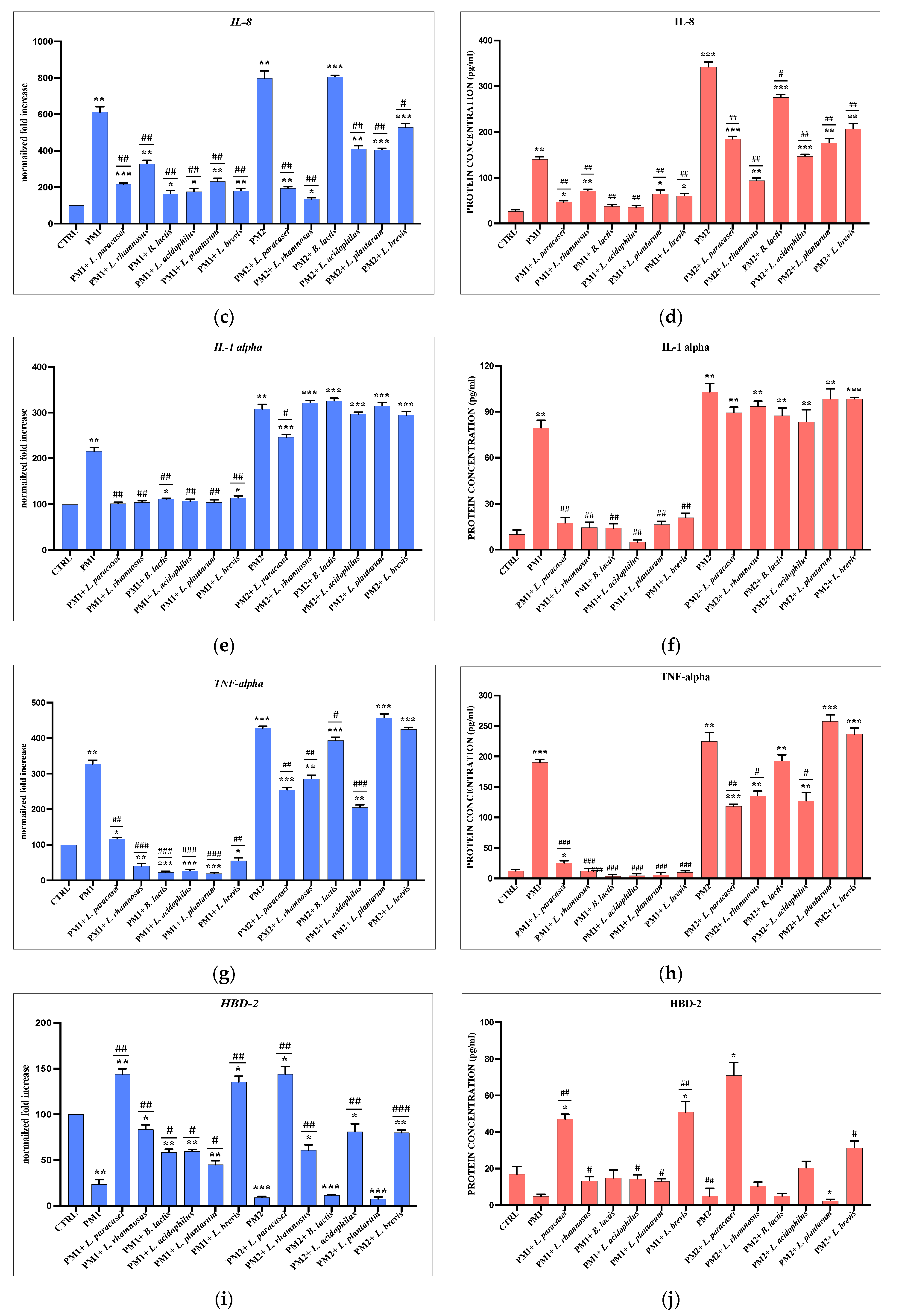
3.3. Analysis of Lactobacillus spp. Anti-Inflammatory Activity against Different P. mirabilis Strains Clinically Isolated in Different Epithelia
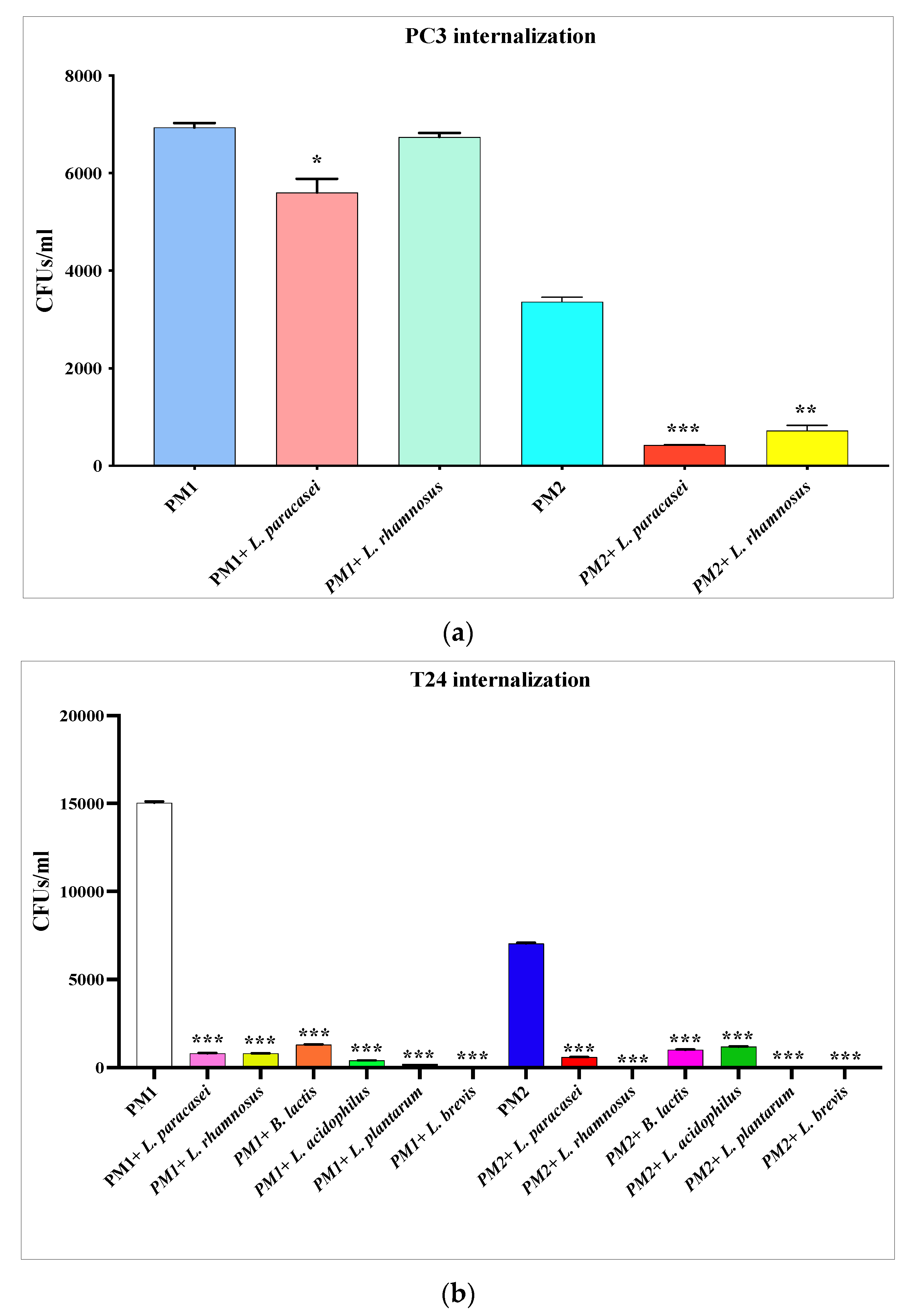
3.4. Inhibition of P. mirabilis Invasiveness
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mancuso, G.; Midiri, A.; Gerace, E.; Marra, M.; Zummo, S.; Biondo, C. Urinary Tract Infections: The Current Scenario and Future Prospects. Pathogens 2023, 12, 623. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Huang, Z.; Yang, T.; Wang, G.; Li, P.; Yang, B.; Li, J. Pathogenesis of Proteus mirabilis in Catheter-Associated Urinary Tract Infections. Urol. Int. 2021, 105, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Kranz, J.; Schmidt, S.; Lebert, C.; Schneidewind, L.; Mandraka, F.; Kunze, M.; Helbig, S.; Vahlensieck, W.; Naber, K.; Schmiemann, G.; et al. The 2017 Update of the German Clinical Guideline on Epidemiology, Diagnostics, Therapy, Prevention, and Management of Uncomplicated Urinary Tract Infections in Adult Patients: Part 1. Urol. Int. 2018, 100, 263–270. [Google Scholar] [CrossRef]
- Suárez Fernández, M.L.; Ridao Cano, N.; Álvarez Santamarta, L.; Gago Fraile, M.; Blake, O.; Díaz Corte, C. A Current Review of the Etiology, Clinical Features, and Diagnosis of Urinary Tract Infection in Renal Transplant Patients. Diagnostics 2021, 11, 1456. [Google Scholar] [CrossRef]
- Fusco, A.; Savio, V.; De Filippis, A.; Tufano, A.; Donnarumma, G. Induction of Different Apoptosis Pathways by Two Proteus mirabilis Clinical Isolates Strains in Prostatic Epithelial Cells. Front. Physiol. 2018, 9, 1855. [Google Scholar] [CrossRef] [PubMed]
- Edefonti, A.; Tel, F.; Testa, S.; De Palma, D. Febrile urinary tract infections: Clinical and laboratory diagnosis, imaging, and prognosis. Semin. Nucl. Med. 2014, 44, 123–128. [Google Scholar] [CrossRef]
- Schaeffer, A.J.; Nicolle, L.E. Clinical practice. urinary tract infections in older men. N. Engl. J. Med. 2016, 374, 562–571. [Google Scholar] [CrossRef]
- Pirola, G.M.; Verdacchi, T.; Rosadi, S.; Annino, F.; De Angelis, M. Chronic prostatitis: Current treatment options. Res. Rep. Urol. 2019, 11, 165–174. [Google Scholar] [CrossRef]
- Drzewiecka, D. Significance and Roles of Proteus spp. Bacteria in Natural Environments. Microb. Ecol. 2016, 72, 741–758. [Google Scholar] [CrossRef]
- Kwiecinska-Piróg, J.; Bogiel, T.; Skowron, K.; Wieckowska, E.; Gospodarek, E. Proteus mirabilis biofilm-qualitative and quantitative colorimetric methods-based evaluation. Braz. J. Microbiol. 2015, 45, 1423–1431. [Google Scholar] [CrossRef]
- Marques, C.; Belas, A.; Menezes, J.; Moreira da Silva, J.; Cavaco-Silva, P.; Trigueiro, G.; Gama, L.T.; Pomba, C. Human and Companion Animal Proteus mirabilis Sharing. Microbiol. Res. 2022, 13, 38–48. [Google Scholar] [CrossRef]
- Zhang, J.; Hoedt, E.C.; Liu, Q.; Berendsen, E.; Teh, J.J.; Hamilton, A.; O’ Brien, A.W.; Ching, J.Y.L.; Wei, H.; Yang, K.; et al. Elucidation of Proteus mirabilis as a Key Bacterium in Crohn’s Disease Inflammation. Gastroenterology 2021, 160, 317–330.e11. [Google Scholar] [CrossRef]
- Gmiter, D.; Pacak, I.; Nawrot, S.; Czerwonka, G.; Kaca, W. Genomes comparison of two Proteus mirabilis clones showing varied swarming ability. Mol. Biol. Rep. 2023, 50, 5817–5826. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and Urinary Tract Infections. Microbiol. Spectr. 2015, 3, 383–433. [Google Scholar] [CrossRef]
- Tabatabaei, A.; Ahmadi, K.; Shabestari, A.N.; Khosravi, N.; Badamchi, A. Virulence genes and antimicrobial resistance pattern in Proteus mirabilis strains isolated from patients attended with urinary infections to Tertiary Hospitals, in Iran. Afr. Health Sci. 2021, 21, 1677–1684. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus mirabilis Infection. EcoSal Plus 2018, 8, 8. [Google Scholar] [CrossRef]
- Wasfi, R.; Hamed, S.M.; Amer, M.A.; Fahmy, L.I. Proteus mirabilis Biofilm: Development and Therapeutic Strategies. Front. Cell. Infect. Microbiol. 2020, 10, 414. [Google Scholar] [CrossRef]
- Girlich, D.; Bonnin, R.A.; Dortet, L.; Naas, T. Genetics of acquired antibiotic resistance genes in Proteus spp. Front. Microbiol. 2020, 11, 256. [Google Scholar] [CrossRef]
- Tegegne, B.A.; Kebede, B. Probiotics, their prophylactic and therapeutic applications in human health development: A review of the literature. Heliyon 2022, 8, e09725. [Google Scholar] [CrossRef]
- Rabetafika, H.N.; Razafindralambo, A.; Ebenso, B.; Razafindralambo, H.L. Probiotics as Antibiotic Alternatives for Human and Animal Applications. Encyclopedia 2023, 3, 561–581. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva RCd Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Imperial, I.C.; Ibana, J.A. Addressing the Antibiotic Resistance Problem with Probiotics: Reducing the Risk of Its Double-Edged Sword Effect. Front. Microbiol. 2016, 7, 1983. [Google Scholar] [CrossRef] [PubMed]
- Varankovich, N.V.; Nickerson, M.T.; Korber, D.R. Probiotic-based strategies for therapeutic and prophylactic use against multiple gastrointestinal diseases. Front. Microbiol. 2015, 6, 685. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhu, G.; Chen, C.; Zheng, Y.; Ma, F.; Zhao, J.; Lee, Y.K.; Zhang, H.; Chen, W. Lactobacillus strains derived from human gut ameliorate metabolic disorders via modulation of gut microbiota composition and short-chain fatty acids metabolism. Benef. Microbes 2021, 12, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, S.; Cai, S.; Yu, H.; Wang, Y.; Zeng, X.; Qiao, S. Lactobacillus reuteri Ameliorates Intestinal Inflammation and Modulates Gut Microbiota and Metabolic Disorders in Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients 2020, 12, 2298. [Google Scholar] [CrossRef]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and Improving Function of Lactobacilli in the Prevention and Treatment of Cardiovascular-Related Diseases: A Novel Perspective From Gut Microbiota. Front. Nutr. 2021, 8, 693412. [Google Scholar] [CrossRef]
- Kwon, M.-S.; Lim, S.K.; Jang, J.-Y.; Lee, J.; Park, H.K.; Kim, N.; Yun, M.; Shin, M.-Y.; Jo, H.E.; Oh, Y.J.; et al. Lactobacillus sakei WIKIM30 Ameliorates Atopic Dermatitis-Like Skin Lesions by Inducing Regulatory T Cells and Altering Gut Microbiota Structure in Mice. Front. Immunol. 2018, 9, 1905. [Google Scholar] [CrossRef]
- Yang, Y.J.; Chuang, C.C.; Yang, H.B.; Lu, C.C.; Sheu, B.S. Lactobacillus acidophilus ameliorates H. pylori-induced gastric inflammation by inactivating the Smad7 and NFκB pathways. BMC Microbiol. 2012, 12, 38. [Google Scholar] [CrossRef]
- D’ambrosio, S.; Ventrone, M.; Fusco, A.; Casillo, A.; Dabous, A.; Cammarota, M.; Corsaro, M.M.; Donnarumma, G.; Schiraldi, C.; Cimini, D. Limosilactobacillus fermentum from buffalo milk is suitable for potential biotechnological process development and inhibits Helicobacter pylori in a gastric epithelial cell model. Biotechnol. Rep. 2022, 34, e00732. [Google Scholar] [CrossRef]
- Fusco, A.; Coretti, L.; Savio, V.; Buommino, E.; Lembo, F.; Donnarumma, G. Biofilm Formation and Immunomodulatory Activity of Proteus mirabilis Clinically Isolated Strains. Int. J. Mol. Sci. 2017, 18, 414. [Google Scholar] [CrossRef]
- Song, C.H.; Kim, Y.H.; Naskar, M.; Hayes, B.W.; Abraham, M.A.; Noh, J.H.; Suk, G.; Kim, M.J.; Cho, K.S.; Shin, M.; et al. Lactobacillus crispatus Limits Bladder Uropathogenic E. coli Infection by Triggering a Host Type I Interferon Response. Proc. Natl. Acad. Sci. USA 2022, 119, e2117904119. [Google Scholar] [CrossRef]
- Johnson, J.A.; Delaney, L.F.; Ojha, V.; Rudraraju, M.; Hintze, K.R.; Siddiqui, N.Y.; Sysoeva, T.A. Commensal Urinary Lactobacilli Inhibit Major Uropathogens In Vitro With Heterogeneity at Species and Strain Level. Front. Cell. Infect. Microbiol. 2022, 12, 870603. [Google Scholar] [CrossRef]
- Cadieux, P.A.; Burton, J.; Devillard, E.; Reid, G. Lactobacillus by-products inhibit the growth and virulence of uropathogenic Escherichia coli. J. Physiol. Pharmacol. 2009, 60 (Suppl. 6), 13–18. [Google Scholar]
- Cimini, D.; D’ambrosio, S.; Stellavato, A.; Fusco, A.; Corsaro, M.M.; Dabous, A.; Casillo, A.; Donnarumma, G.; Giori, A.M.; Schiraldi, C. Optimization of growth of Levilactobacillus brevis SP 48 and in vitro evaluation of the effect of viable cells and high molecular weight potential postbiotics on Helicobacter pylori. Front. Bioeng. Biotechnol. 2022, 10, 1007004. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. JoVE 2011, 47, 2437. [Google Scholar] [CrossRef]
- Saunders, R.K., Jr.; Infanti, J.; Ali, H.; Shuey, T.; Potteiger, C.; McNeilly, S.; Adams, C.S. Gram-Negative Bacteria Are Internalized Into Osteocyte-Like Cells. J. Orthop. Res. 2020, 38, 861–870. [Google Scholar] [CrossRef]
- Tang, H.; Yang, D.; Zhu, L.; Shi, F.; Ye, G.; Guo, H.; Deng, H.; Zhao, L.; Xu, Z.; Li, Y. Paeonol Interferes With Quorum-Sensing in Pseudomonas aeruginosa and Modulates Inflammatory Responses In Vitro and In Vivo. Front. Immunol 2022, 13, 896874. [Google Scholar] [CrossRef]
- Fusco, A.; Savio, V.; Cammarota, M.; Alfano, A.; Schiraldi, C.; Donnarumma, G. Beta-Defensin-2 and Beta-Defensin-3 Reduce Intestinal Damage Caused by Salmonella typhimurium Modulating the Expression of Cytokines and Enhancing the Probiotic Activity of Enterococcus faecium. J. Immunol. Res. 2017, 2017, 6976935. [Google Scholar] [CrossRef]
- Fusco, A.; Savio, V.; Perfetto, B.; Mattina, R.; Donnarumma, G. Antimicrobial peptide human β-defensin-2 improves in vitro cellular viability and reduces pro-inflammatory effects induced by enteroinvasive Escherichia coli in Caco-2 cells by inhibiting invasion and virulence factors’ expression. Front. Cell. Infect. Microbiol. 2022, 12, 1009415. [Google Scholar] [CrossRef]
- Liu, L.; Cui, J.; Zhao, Y.; Liu, X.; Chen, L.; Xia, Y.; Wang, Y.; Chen, S.; Sun, S.; Shi, B.; et al. KDM6A-ARHGDIB axis blocks metastasis of bladder cancer by inhibiting Rac1. Mol. Cancer 2021, 20, 77. [Google Scholar] [CrossRef]
- de Siqueira e Silva, M.F.; de Sena, F.C.R.; Agostinho, F.; Almeida Medeiros, K.K.; Miguel, C.B. Urinary tract infection related to hospitalized patients: A Review. Arch. Urol. Res. 2017, 1, 001–004. [Google Scholar] [CrossRef]
- Delcaru, C.; Podgoreanu, P.; Alexandru, I.; Popescu, N.; Măruţescu, L.; Bleotu, C.; Mogoşanu, G.D.; Chifiriuc, M.C.; Gluck, M.; Lazăr, V. Antibiotic Resistance and Virulence Phenotypes of Recent Bacterial Strains Isolated from Urinary Tract Infections in Elderly Patients with Prostatic Disease. Pathogens 2017, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Huang, C.; Yan, Y.; Sun, L.; Li, H. Urinary Tract Infection Etiological Profiles and Antibiotic Resistance Patterns Varied Among Different Age Categories: A Retrospective Study From a Tertiary General Hospital During a 12-Year Period. Front. Microbiol. 2022, 12, 813145. [Google Scholar] [CrossRef] [PubMed]
- Baraniak, A.; Izdebski, R.; Fiett, J.; Gawryszewska, I.; Bojarska, K.; Herda, M.; Literacka, E.; Żabicka, D.; Tomczak, H.; Pewińska, N.; et al. NDM-producing Enterobacteriaceae in Poland, 2012–2014: Inter-regional outbreak of Klebsiella pneumoniae ST11 and sporadic cases. J. Antimicrob. Chemother. 2016, 71, 85–91. [Google Scholar] [CrossRef]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: A systematic review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef]
- Literacka, E.; Izdebski, R.; Baraniak, A.; Żabicka, D.; Schneider, A.; Urbanowicz, P.; Herda, M.; Hryniewicz, W.; Gniadkowski, M. Proteus mirabilis Producing the OXA-58 Carbapenemase in Poland. Antimicrob. Agents Chemother. 2019, 63, e00106-19. [Google Scholar] [CrossRef]
- Zamojska, D.; Nowak, A.; Nowak, I.; Macierzyńska-Piotrowska, E. Probiotics and Postbiotics as Substitutes of Antibiotics in Farm Animals: A Review. Animals 2021, 11, 3431. [Google Scholar] [CrossRef]
- Leistikow, K.R.; Beattie, R.E.; Hristova, K.R. Probiotics beyond the farm: Benefits, costs, and considerations of using antibiotic alternatives in livestock. Front. Antibiot. 2022, 1, 1003912. [Google Scholar] [CrossRef]
- Fusco, A.; Savio, V.; Cimini, D.; D’Ambrosio, S.; Chiaromonte, A.; Schiraldi, C.; Donnarumma, G. In Vitro Evaluation of the Most Active Probiotic Strains Able to Improve the Intestinal Barrier Functions and to Prevent Inflammatory Diseases of the Gastrointestinal System. Biomedicines 2023, 11, 865. [Google Scholar] [CrossRef]
- Loubet, P.; Ranfaing, J.; Dinh, A.; Dunyach-Remy, C.; Bernard, L.; Bruyère, F.; Lavigne, J.P.; Sotto, A. Alternative Therapeutic Options to Antibiotics for the Treatment of Urinary Tract Infections. Front. Microbiol. 2020, 11, 1509. [Google Scholar] [CrossRef]
- Cai, T.; Gallelli, L.; Cione, E.; Perletti, G.; Ciarleglio, F.; Malossini, G.; De Pretis, G.; Palmieri, A.; Mirone, V.; Bartoletti, R.; et al. The use of Lactobacillus casei DG® prevents symptomatic episodes and reduces the antibiotic use in patients affected by chronic bacterial prostatitis: Results from a phase IV study. World J. Urol. 2021, 39, 3433–3440. [Google Scholar] [CrossRef] [PubMed]
- Ng, Q.X.; Peters, C.; Venkatanarayanan, N.; Goh, Y.Y.; Ho, C.Y.X.; Yeo, W.S. Use of Lactobacillus spp. to prevent recurrent urinary tract infections in females. Med. Hypotheses 2018, 114, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and therapeutic effectiveness of lactobacilli. J. Clin. Pathol. 2016, 69, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, J.N.; Norsworthy, A.N.; Sun, T.T.; Pearson, M.M. Proteus mirabilis fimbriae- and urease-dependent clusters assemble in an extracellular niche to initiate bladder stone formation. Proc. Natl. Acad. Sci. USA 2016, 113, 4494–4499. [Google Scholar] [CrossRef]
- Zabłotni, A.; Matusiak, D.; Arbatsky, N.P.; Moryl, M.; Maciejewska, A.; Kondakova, A.N.; Shashkov, A.S.; Ługowski, C.; Knirel, Y.A.; Różalski, A. Changes in the lipopolysaccharide of Proteus mirabilis 9B-m (O11a) clinical strain in response to planktonic or biofilm type of growth. Med. Microbiol. Immunol. 2018, 207, 129–139. [Google Scholar] [CrossRef]
- Moryl, M.; Torzewska, A.; Jałmuzna, P.; Rózalski, A. Analysis of Proteus mirabilis distribution in multi-species biofilms on urinary catheters and determination of bacteria resistance to antimicrobial agents. Pol. J. Microbiol. 2013, 62, 377–384. [Google Scholar] [CrossRef]
- Morgenstein, R.M.; Szostek, B.; Rather, P.N. Regulation of gene expression during swarmer cell differentiation in Proteus mirabilis. FEMS Microbiol. Rev. 2010, 34, 753–763. [Google Scholar] [CrossRef]
- Silva, D.R.; Sardi, J.d.C.O.; Pitangui, N.d.S.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Stapleton, A.E.; Au-Yeung, M.; Hooton, T.M.; Fredricks, D.N.; Roberts, P.L.; Czaja, C.A.; Yarova-Yarovaya, Y.; Fiedler, T.; Cox, M.; Stamm, W.E. Randomized, placebo-controlled phase 2 trial of a Lactobacillus crispatus probiotic given intravaginally for prevention of recurrent urinary tract infection. Clin. Infect. Dis. 2011, 52, 1212–1217. [Google Scholar] [CrossRef]
- Sadahira, T.; Wada, K.; Araki, M.; Mitsuhata, R.; Yamamoto, M.; Maruyama, Y.; Iwata, T.; Watanabe, M.; Watanabe, T.; Kariyama, R.; et al. Efficacy of Lactobacillus vaginal suppositories for the prevention of recurrent cystitis: A phase II clinical trial. Int. J. Urol. 2021, 28, 1026–1031. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Conditions | Product Size (bp) |
|---|---|---|---|
| IL-6 | 5′-ATGAACTCCTTCTCCACAAGCGC-3′ 5′-GAAGAGCCCTCAGGCTGGACTG-3′ | 5″ at 95 °C, 13″ at 56 °C, 25″ at 72 °C for 40 cycles | 628 |
| IL-8 | 5′-ATGACTTCCAAGCTGGCCGTG-3′ 5′-TGAATTCTCAGCCCTCTTCAAAAACTTCTC-3′ | 5″ at 94 °C, 6″ at 55 °C, 12″ at 72 °C for 40 cycles | 297 |
| IL-1a | 5′-GCATCCAGCTACGAATCTCC-3′ 5′-CCACATTCAGCACAGGACTC-3′ | 5″ at 95 °C, 14″ at 58 °C, 28″ at 72 °C for 40 cycles | 708 |
| TNF-a | 5′-CAGAGGGAAGAGTTCCCCAG-3′ 5′-CCTTGGTCTGGTAGGAGACG-3′ | 5″ at 95 °C, 6″ at 57 °C, 13″ at 72 °C for 40 cycles | 324 |
| HBD-2 | 5′-GGATCCATGGGTATAGGCGATCCTGTTA-3′ 5′-AAGCTTCTCTGATGAGGGAGCCCTTTCT-3′ | 5″ at 94 °C, 6″ at 63 °C, 10″ at 72 °C for 50 cycles | 198 |
| flhDC | 5′-CGCACATCAGCCTGCAAGT-3′ 5′-GCAGGATTGGCGGAAAGTT-3′ | 5″ at 94 °C, 6″ at 53 °C, 7″ at 72 °C for 40 cycles | 90 |
| β-actin | 5′-GACGACGACAAGATAGCCTAGCAGCTATGAGGATC-3′ 5′-GAGGAGAAGCCCGGTTAACTTCCGCAGCATTTTGCGCCA-3′ | 243 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, A.; Savio, V.; Chiaromonte, A.; Alfano, A.; D’Ambrosio, S.; Cimini, D.; Donnarumma, G. Evaluation of Different Activity of Lactobacillus spp. against Two Proteus mirabilis Isolated Clinical Strains in Different Anatomical Sites In Vitro: An Explorative Study to Improve the Therapeutic Approach. Microorganisms 2023, 11, 2201. https://doi.org/10.3390/microorganisms11092201
Fusco A, Savio V, Chiaromonte A, Alfano A, D’Ambrosio S, Cimini D, Donnarumma G. Evaluation of Different Activity of Lactobacillus spp. against Two Proteus mirabilis Isolated Clinical Strains in Different Anatomical Sites In Vitro: An Explorative Study to Improve the Therapeutic Approach. Microorganisms. 2023; 11(9):2201. https://doi.org/10.3390/microorganisms11092201
Chicago/Turabian StyleFusco, Alessandra, Vittoria Savio, Adriana Chiaromonte, Alberto Alfano, Sergio D’Ambrosio, Donatella Cimini, and Giovanna Donnarumma. 2023. "Evaluation of Different Activity of Lactobacillus spp. against Two Proteus mirabilis Isolated Clinical Strains in Different Anatomical Sites In Vitro: An Explorative Study to Improve the Therapeutic Approach" Microorganisms 11, no. 9: 2201. https://doi.org/10.3390/microorganisms11092201