
Deionococcus proteotlycius Genomic Library Exploration Enhances Oxidative Stress Resistance and Poly-3-hydroxybutyrate Production in Recombinant Escherichia coli
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids and Culture Conditions
2.2. Screening of Oxidative-Resistant Clones Using the Genomic DNA Library
2.3. Construction of Deipr_0871 Expression Vector
2.4. Determination of Growth and Survival of the Recombinant E. coli Strains
2.5. Real-Time PCR (qPCR)
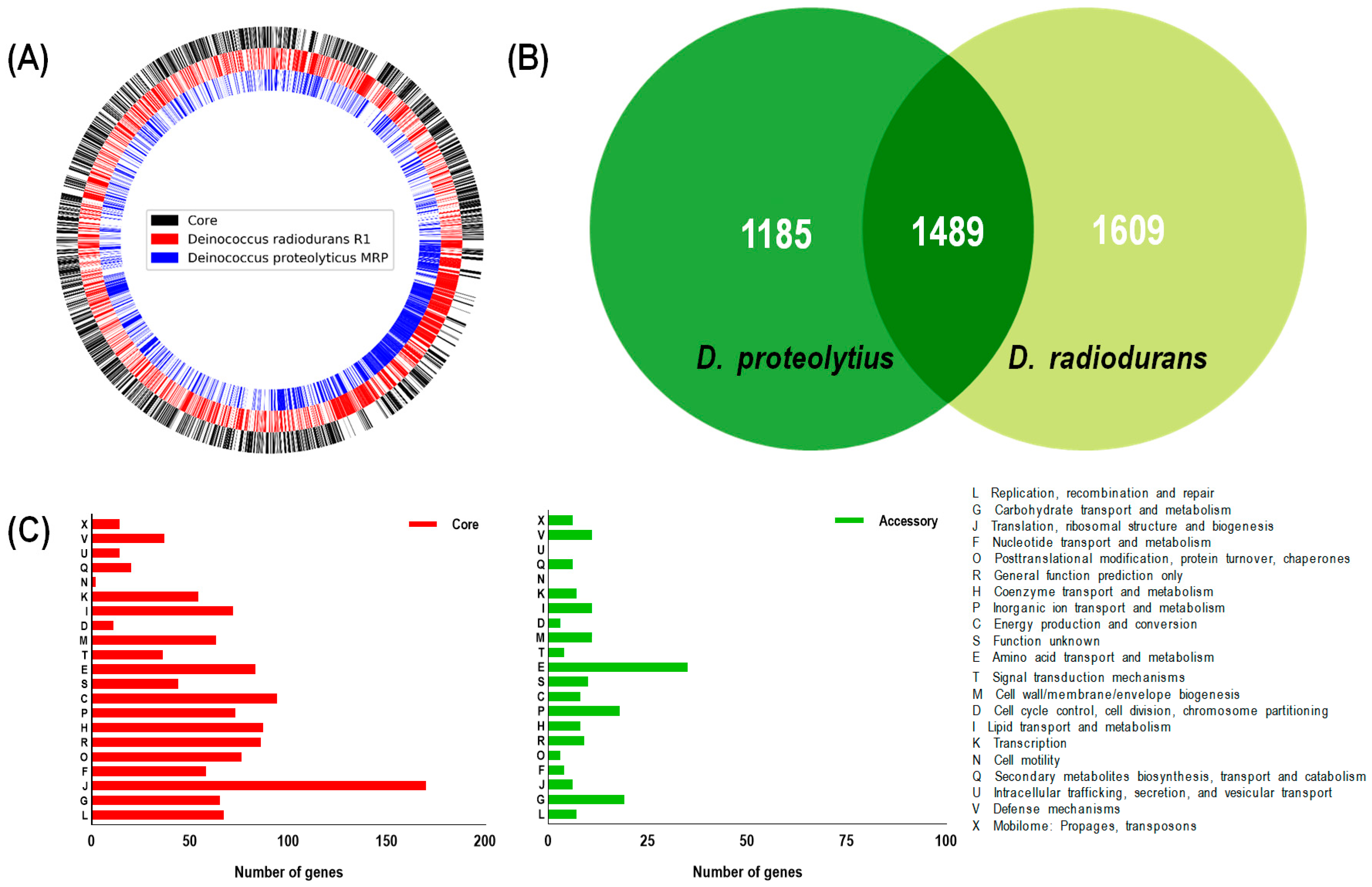
2.6. Genome Analysis
2.7. Cultivation for PHB Production
2.8. Measurement of Amount of PHB
2.9. Sequence Alignment
3. Results
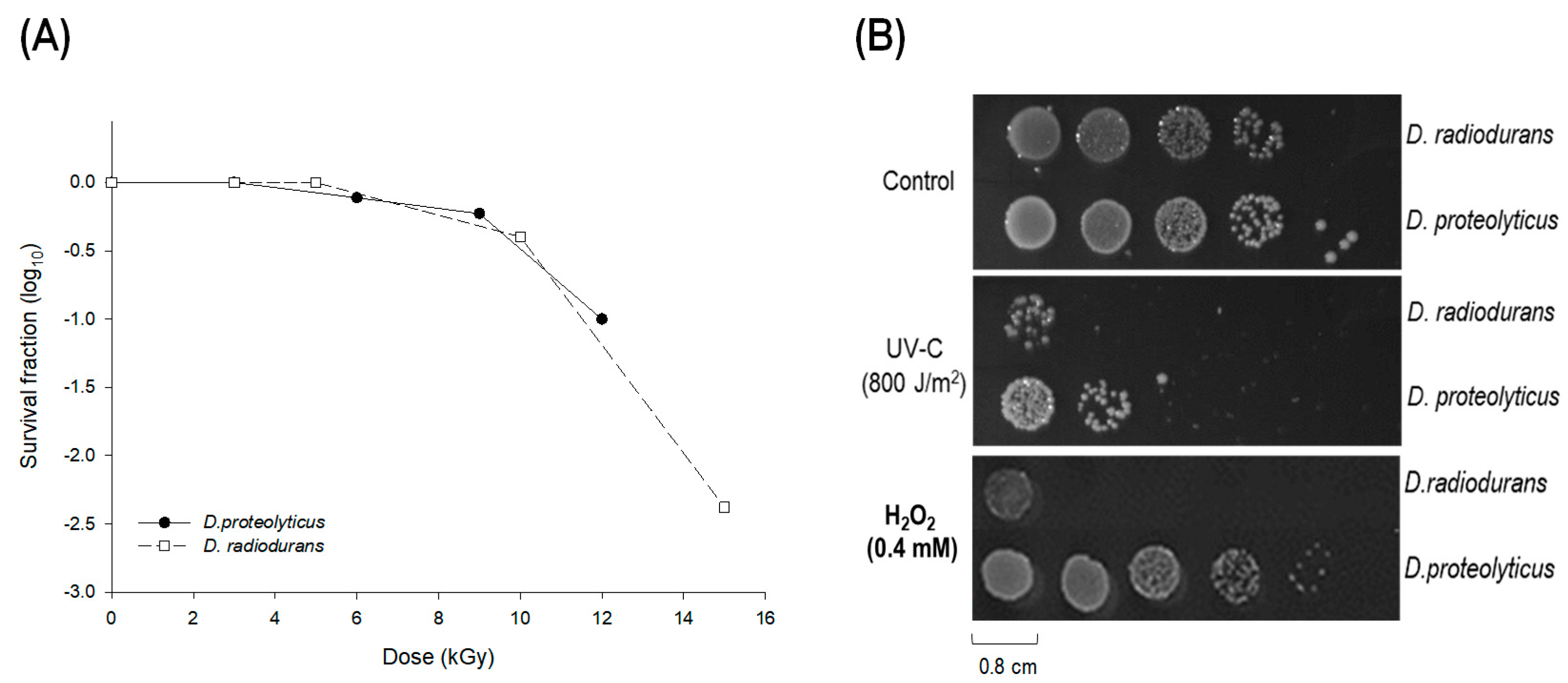
3.1. The Oxidative Stress Resistance Properties of Deinococcus Proteolyticus
3.2. Screening Genomic Libraries of D. proteolyticus for Genes Improving Oxidative Stress Resistance to E. coli
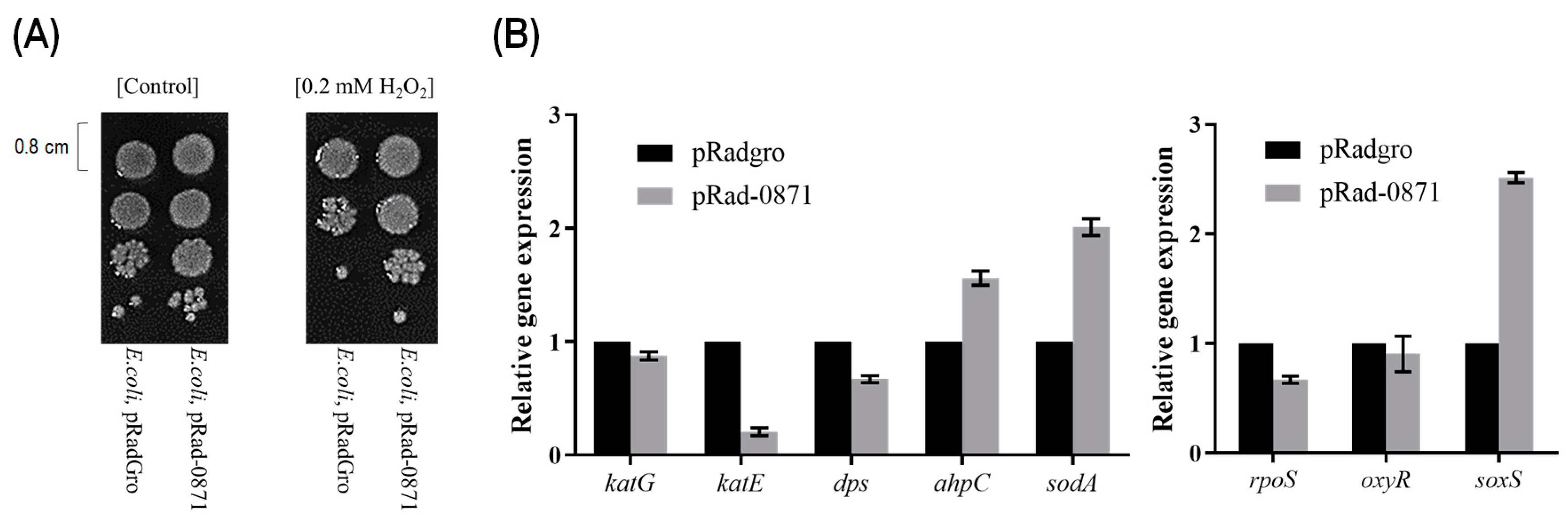
3.3. Effect of Deipr_0871 on the Oxidative Stress Resistance of E. coli Strain
3.4. Effect of Deipr_0871 on Growth of E. coli Strain
3.5. Effect of Deipr_0871 on Valuable Metabolite, PHB, Production in the E. coli Strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrer-Miralles, N.; Domingo-Espin, J.; Corchero, J.L.; Vazquez, E.; Villaverde, A. Microbial factories for recombinant pharmaceuticals. Microb. Cell Factories 2009, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- McElwain, L.; Phair, K.; Kealey, C.; Brady, D. Current trends in biopharmaceuticals production in Escherichia coli. Biotechnol. Lett. 2022, 44, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, G.A. From scratch to value: Engineering Escherichia coli wild type cells to the production of L-phenylalanine and other fine chemicals derived from chorismate. Appl. Microbiol. Biotechnol. 2007, 75, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-X.; Xiao, W.-H.; Liu, D.; Zhang, J.-L.; Ding, M.-Z.; Yuan, Y.-J. Biosynthesis of odd-chain fatty alcohols in Escherichia coli. Metab. Eng. 2015, 29, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lee, S.Y. Microbial production of short-chain alkanes. Nature 2013, 502, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Shiloach, J.; Fass, R. Growing E. coli to high cell density—A historical perspective on method development. Biotechnol. Adv. 2005, 23, 345–357. [Google Scholar] [CrossRef]
- Castan, A.; Näsman, A.; Enfors, S.-O. Oxygen enriched air supply in Escherichia coli processes: Production of biomass and recombinant human growth hormone. Enzyme Microb. Technol. 2002, 30, 847–854. [Google Scholar] [CrossRef]
- Baez, A.; Shiloach, J. Escherichia coli avoids high dissolved oxygen stress by activation of SoxRS and manganese-superoxide dismutase. Microb. Cell Factories 2013, 12, 23. [Google Scholar] [CrossRef]
- Ahn, Y.-J.; Im, E. Heterologous expression of heat shock proteins confers stress tolerance in Escherichia coli, an industrial cell factory: A short review. Biocatal. Agric. Biotechnol. 2020, 29, 101833. [Google Scholar] [CrossRef]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef]
- Basak, S.; Jiang, R. Enhancing E. coli tolerance towards oxidative stress via engineering its global regulator cAMP receptor protein (CRP). PLoS ONE 2012, 7, e51179. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wang, H.; Wang, J.; Jiang, W.; Jiang, Y.; Xin, F.; Zhang, W.; Jiang, M. Strategies to improve the stress resistance of Escherichia coli in industrial biotechnology. Biofuel Bioprod. Biorefining 2022, 16, 1130–1141. [Google Scholar] [CrossRef]
- Park, S.J.; Lim, S.; Choi, J.I. Improved tolerance of Escherichia coli to oxidative stress by expressing putative response regulator homologs from Antarctic bacteria. J. Microbiol. 2020, 58, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, S.A.; Fast, A.G.; Nakamaru-Ogiso, E.; Papoutsakis, E.T. Overexpression of fetA (ybbL) and fetB (ybbM), Encoding an Iron Exporter, Enhances Resistance to Oxidative Stress in Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 7210–7219. [Google Scholar] [CrossRef]
- Krisko, A.; Radman, M. Biology of extreme radiation resistance: The way of Deinococcus radiodurans. Cold Spring Harb. Perspect. Biol. 2013, 5, a012765. [Google Scholar] [CrossRef]
- Earl, A.M.; Mohundro, M.M.; Mian, I.S.; Battista, J.R. The IrrE protein of Deinococcus radiodurans R1 is a novel regulator of recA expression. J. Bacteriol. 2002, 184, 6216–6224. [Google Scholar] [CrossRef]
- Khairnar, N.P.; Misra, H.S.; Apte, S.K. Pyrroloquinoline–quinone synthesized in Escherichia coli by pyrroloquinoline–quinone synthase of Deinococcus radiodurans plays a role beyond mineral phosphate solubilization. Biochem. Biophys. Res. Commun. 2003, 312, 303–308. [Google Scholar] [CrossRef]
- Pan, J.; Wang, J.; Zhou, Z.; Yan, Y.; Zhang, W.; Lu, W.; Ping, S.; Dai, Q.; Yuan, M.; Feng, B.; et al. IrrE, a global regulator of extreme radiation resistance in Deinococcus radiodurans, enhances salt tolerance in Escherichia coli and Brassica napus. PLoS ONE 2009, 4, e4422. [Google Scholar] [CrossRef]
- Dong, X.; Tian, B.; Dai, S.; Li, T.; Guo, L.; Tan, Z.; Jiao, Z.; Jin, Q.; Wang, Y.; Hua, Y. Expression of PprI from Deinococcus radiodurans Improves Lactic Acid Production and Stress Tolerance in Lactococcus lactis. PLoS ONE 2015, 10, e0142918. [Google Scholar] [CrossRef]
- Appukuttan, D.; Singh, H.; Park, S.H.; Jung, J.H.; Jeong, S.; Seo, H.S.; Choi, Y.J.; Lim, S. Engineering Synthetic Multistress Tolerance in Escherichia coli by Using a Deinococcal Response Regulator, DR1558. Appl. Environ. Microbiol. 2016, 82, 1154–1166. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Zhao, Z.; Zhou, Z.; Lu, W.; Zhang, W.; Chen, M. irrE, an exogenous gene from Deinococcus radiodurans, improves the growth of and ethanol production by a Zymomonas mobilis strain under ethanol and acid stress. J. Microbiol. Biotechnol. 2010, 20, 1156–1162. [Google Scholar] [CrossRef]
- Guo, S.; Yi, X.; Zhang, W.; Wu, M.; Xin, F.; Dong, W.; Zhang, M.; Ma, J.; Wu, H.; Jiang, M. Inducing hyperosmotic stress resistance in succinate-producing Escherichia coli by using the response regulator DR1558 from Deinococcus radiodurans. Process Biochem. 2017, 61, 30–37. [Google Scholar] [CrossRef]
- Park, S.H.; Sohn, Y.J.; Park, S.J.; Choi, J.I. Effect of DR1558, a Deinococcus radiodurans response regulator, on the production of GABA in the recombinant Escherichia coli under low pH conditions. Microb. Cell Factories 2020, 19, 64. [Google Scholar] [CrossRef]
- Kang, S.-b.; Choi, J.-i. Enhanced cadaverine production by recombinant Corynebacterium glutamicum with a heterologous DR1558 regulator at low pH condition. Process Biochem. 2021, 111, 63–70. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, G.B.; Kim, H.U.; Park, S.J.; Choi, J.I. Enhanced production of poly-3-hydroxybutyrate (PHB) by expression of response regulator DR1558 in recombinant Escherichia coli. Int. J. Biol. Macromol. 2019, 131, 29–35. [Google Scholar] [CrossRef]
- Lim, S.; Jung, J.H.; Blanchard, L.; de Groot, A. Conservation and diversity of radiation and oxidative stress resistance mechanisms in Deinococcus species. FEMS Microbiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Vera Alvarez, R.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic Acids Res. 2021, 49, D274–D281. [Google Scholar] [CrossRef]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R. A rapid gas chromatographic method for the determination of poly-β-hydroxybutyric acid in microbial biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. The Clustal Omega multiple alignment package. Methods Mol. Biol. 2021, 2231, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Copeland, A.; Zeytun, A.; Yassawong, M.; Nolan, M.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.F.; Han, C.; Tapia, R.; et al. Complete genome sequence of the orange-red pigmented, radioresistant Deinococcus proteolyticus type strain (MRP(T)). Stand. Genom. Sci. 2012, 6, 240–250. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef]
- Seo, S.W.; Kim, D.; Szubin, R.; Palsson, B.O. Genome-wide reconstruction of OxyR and SoxRS transcriptional regulatory networks under oxidative stress in Escherichia coli K-12 MG1655. Cell Rep. 2015, 12, 1289–1299. [Google Scholar] [CrossRef]
- Mitrophanov, A.Y.; Groisman, E.A. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef]
- Rodríguez-Moya, J.; Argandoña, M.; Reina-Bueno, M.; Nieto, J.J.; Iglesias-Guerra, F.; Jebbar, M.; Vargas, C. Involvement of EupR, a response regulator of the NarL/FixJ family, in the control of the uptake of the compatible solutes ectoines by the halophilic bacterium Chromohalobacter salexigens. BMC Microbiol. 2010, 10, 256. [Google Scholar] [CrossRef]
- Triggs-Raine, B.L.; Doble, B.W.; Mulvey, M.R.; Sorby, P.A.; Loewen, P.C. Nucleotide sequence of katG, encoding catalase HPI of Escherichia coli. J. Bacteriol. 1988, 170, 4415–4419. [Google Scholar] [CrossRef]
- Henard, C.A.; Bourret, T.J.; Song, M.; Vazquez-Torres, A. Control of redox balance by the stringent response regulatory protein promotes antioxidant defenses of Salmonella. J. Biol. Chem. 2010, 285, 36785–36793. [Google Scholar] [CrossRef]
- Varghese, S.; Tang, Y.; Imlay, J.A. Contrasting sensitivities of Escherichia coli aconitases A and B to oxidation and iron depletion. J. Bacteriol. 2003, 185, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Rungrassamee, W.; Liu, X.; Pomposiello, P.J. Activation of glucose transport under oxidative stress in Escherichia coli. Arch. Microbiol. 2008, 190, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Chandani, N.; Mazumder, P.; Bhattacharjee, A. Production of polyhydroxybutyrate (biopolymer) by Bacillus tequilensis NCS-3 isolated from municipal waste areas of Silchar, Assam. Int. J. Sci. Res. 2014, 3, 198–203. [Google Scholar]
- Jiang, Y.; Marang, L.; Kleerebezem, R.; Muyzer, G.; van Loosdrecht, M.C. Polyhydroxybutyrate production from lactate using a mixed microbial culture. Biotechnol. Bioeng. 2011, 108, 2022–2035. [Google Scholar] [CrossRef]
- Sekar, K.; Tyo, K.E.J. Regulatory effects on central carbon metabolism from poly-3-hydroxybutryate synthesis. Metab. Eng. 2015, 28, 180–189. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | DCW (g/L) | Yield (gPHB/gGlucose) | PHB Conc. (g/L) | PHB Content (wt%) |
|---|---|---|---|---|
| E. coli XL1-Blue (pKMCAB+pRad-0871) | 3.88 ± 0.09 | 0.12 ± 0.004 | 3.139 ± 0.07 | 80 ± 3 |
| E. coli XL1-Blue (pKMCAB+pRadGro) | 1.96 ± 0.05 | 0.03 ± 0.007 | 0.731 ± 0.03 | 38 ± 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-K.; Jeong, S.; Baek, I.; Choi, J.-i.; Lim, S.; Jung, J.-H. Deionococcus proteotlycius Genomic Library Exploration Enhances Oxidative Stress Resistance and Poly-3-hydroxybutyrate Production in Recombinant Escherichia coli. Microorganisms 2023, 11, 2135. https://doi.org/10.3390/microorganisms11092135
Yang S-K, Jeong S, Baek I, Choi J-i, Lim S, Jung J-H. Deionococcus proteotlycius Genomic Library Exploration Enhances Oxidative Stress Resistance and Poly-3-hydroxybutyrate Production in Recombinant Escherichia coli. Microorganisms. 2023; 11(9):2135. https://doi.org/10.3390/microorganisms11092135
Chicago/Turabian StyleYang, Seul-Ki, Soyoung Jeong, Inwoo Baek, Jong-il Choi, Sangyong Lim, and Jong-Hyun Jung. 2023. "Deionococcus proteotlycius Genomic Library Exploration Enhances Oxidative Stress Resistance and Poly-3-hydroxybutyrate Production in Recombinant Escherichia coli" Microorganisms 11, no. 9: 2135. https://doi.org/10.3390/microorganisms11092135