
Degradation of Bisphenol A by Bacillus subtilis P74 Isolated from Traditional Fermented Soybean Foods


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Bacteria
2.2. Culture Conditions
2.3. Measurement of BPA Degradation
2.4. Detection of Metabolites Generated from BPA Degradation
2.5. Soybean Fermentation
2.6. Statistical Analysis
3. Results
3.1. Screening of Bacteria with High BPA Removal Ability Isolated from Fermented Soybean Products
3.2. Effects of Culture Conditions on BPA Degradation by B. subtilis P74
3.2.1. Effects of BPA Concentration and Temperature on BPA Degradation from B. subtilis P74
3.2.2. Effects of pH on BPA Degradation by B. subtilis P74
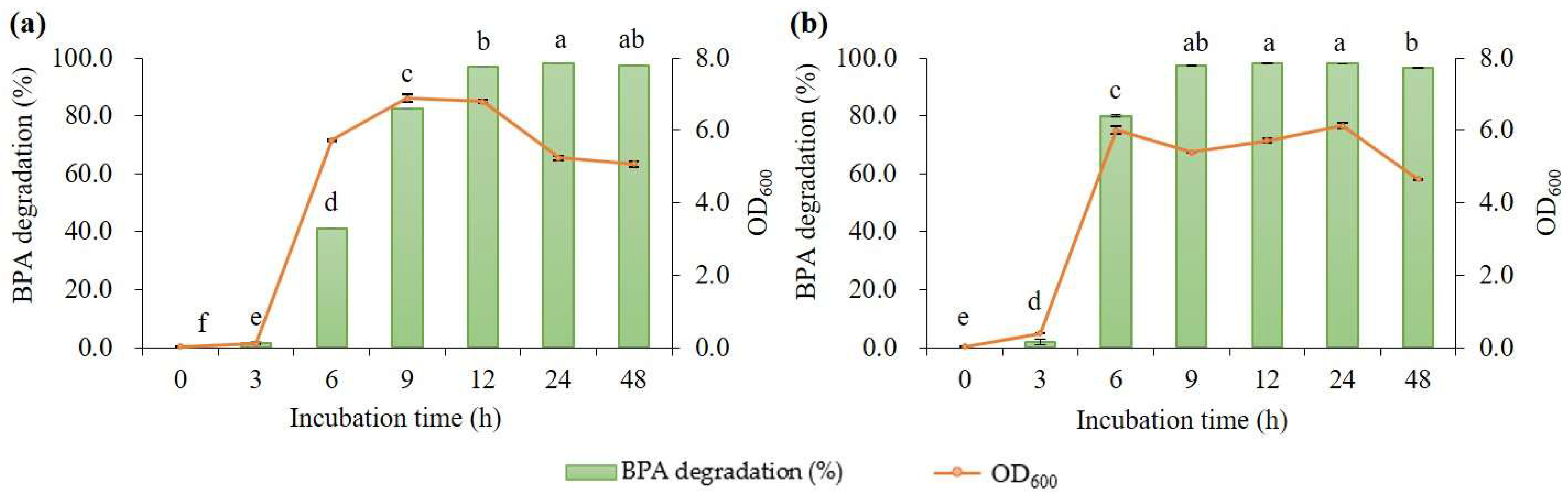
3.2.3. BPA Degradation Performance of B. subtilis P74
3.2.4. Effects of Initial Cell Concentration on BPA Degradation by B. subtilis P74
3.3. Mechanism of BPA Degradation by B. subtilis P74
3.4. BPA Degradation by B. subtilis P74 during Soybean Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Flint, S.; Markle, T.; Thompson, S.; Wallace, E. Bisphenol A exposure, effects, and policy: A wildlife perspective. J. Environ. Manag. 2012, 104, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Suzuki, T.; Sanoh, S.; Kohta, R.; Jinno, N.; Sugihara, K.; Yoshihara, S.i.; Fujimoto, N.; Watanabe, H.; Ohta, S. Comparative study of the endocrine-disrupting activity of bisphenol A and 19 related compounds. Toxicol. Sci. 2005, 84, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.G.; Correia, J.; Adiga, D.; Rai, P.S.; Dsouza, H.S.; Chakrabarty, S.; Kabekkodu, S.P. A comprehensive review on the carcinogenic potential of bisphenol A: Clues and evidence. Environ. Sci. Pollut. R. 2021, 28, 19643–19663. [Google Scholar] [CrossRef] [PubMed]
- Seachrist, D.D.; Bonk, K.W.; Ho, S.-M.; Prins, G.S.; Soto, A.M.; Keri, R.A. A review of the carcinogenic potential of bisphenol A. Reprod. Toxicol. 2016, 59, 167–182. [Google Scholar] [CrossRef]
- Engin, A.B.; Engin, A. The effect of environmental Bisphenol A exposure on breast cancer associated with obesity. Environ. Toxicol. Pharmacol. 2021, 81, 103544. [Google Scholar] [CrossRef]
- Jalal, N.; Surendranath, A.R.; Pathak, J.L.; Yu, S.; Chung, C.Y. Bisphenol A (BPA) the mighty and the mutagenic. Toxicol. Rep. 2018, 5, 76–84. [Google Scholar] [CrossRef]
- Ziv-Gal, A.; Flaws, J.A. Evidence for bisphenol A-induced female infertility: A review (2007–2016). Fertil. Steril. 2016, 106, 827–856. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, W.; Sun, S.; Du, X.; Han, Y.; Shi, W.; Liu, G. Immunotoxicity and neurotoxicity of bisphenol A and microplastics alone or in combination to a bivalve species, Tegillarca granosa. Environ. Pollut. 2020, 265, 115115. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, X.-S.; Cai, Z.-H. The impacts of bisphenol A (BPA) on abalone (Haliotis diversicolor supertexta) embryonic development. Chemosphere 2011, 82, 443–450. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, Z.; Meng, S.; Pan, C.; Yang, M.; Wu, X.; Yang, L.; Xu, H. Parental exposure to bisphenol A and its analogs influences zebrafish offspring immunity. Sci. Total Environ. 2018, 610, 291–297. [Google Scholar] [CrossRef]
- Koch, H.M.; Kolossa-Gehring, M.; Schröter-Kermani, C.; Angerer, J.; Brüning, T. Bisphenol A in 24 h urine and plasma samples of the German Environmental Specimen Bank from 1995 to 2009: A retrospective exposure evaluation. J. Exp. Sci. Environ. Epidemiol. 2012, 22, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Wiraagni, I.A.; Mohd, M.A.; bin Abd Rashid, R.; Haron, D.E.b.M. Validation of a simple extraction procedure for bisphenol A identification from human plasma. PLoS ONE 2019, 14, e0221774. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mao, W.; Yao, L.; Zhao, N.; Zhang, Y.; Zhao, M.; Jin, H. First report on occurrence of bisphenol A isomers in human serum and whole blood. J. Hazard. Mater. 2022, 424, 127549. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, G.; Rice, D.C. Does rapid metabolism ensure negligible risk from bisphenol A? Environ. Health Perspect. 2009, 117, 1639–1643. [Google Scholar] [CrossRef]
- Sun, Y.; Irie, M.; Kishikawa, N.; Wada, M.; Kuroda, N.; Nakashima, K. Determination of bisphenol A in human breast milk by HPLC with column-switching and fluorescence detection. Biomed. Chromatogr. 2004, 18, 501–507. [Google Scholar] [CrossRef]
- Quintella, C.M.; Mata, A.M.; Lima, L.C. Overview of bioremediation with technology assessment and emphasis on fungal bioremediation of oil contaminated soils. J. Environ. Manag. 2019, 241, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Sarma, H.; Nava, A.R.; Manriquez, A.M.E.; Dominguez, D.C.; Lee, W.-Y. Biodegradation of bisphenol A by bacterial consortia isolated directly from river sediments. Environ. Technol. Innov. 2019, 14, 100314. [Google Scholar] [CrossRef]
- Kyrila, G.; Katsoulas, A.; Schoretsaniti, V.; Rigopoulos, A.; Rizou, E.; Doulgeridou, S.; Sarli, V.; Samanidou, V.; Touraki, M. Bisphenol A removal and degradation pathways in microorganisms with probiotic properties. J. Hazard. Mater. 2021, 413, 125363. [Google Scholar] [CrossRef]
- López-Moreno, A.; Torres-Sánchez, A.; Acuña, I.; Suárez, A.; Aguilera, M. Representative Bacillus sp. AM1 from gut microbiota harbor versatile molecular pathways for bisphenol A biodegradation. Int. J. Mol. Sci. 2021, 22, 4952. [Google Scholar] [CrossRef]
- Mohan, H.; Lim, J.-M.; Lee, S.-W.; Cho, M.; Park, Y.-J.; Seralathan, K.-K.; Oh, B.-T. Enhanced removal of bisphenol A from contaminated soil by coupling Bacillus subtilis HV-3 with electrochemical system. Chemosphere 2020, 249, 126083. [Google Scholar] [CrossRef]
- Yamanaka, H.; Moriyoshi, K.; Ohmoto, T.; Ohe, T.; Sakai, K. Degradation of bisphenol A by Bacillus pumilus isolated from kimchi, a traditionally fermented food. Appl. Biochem. Biotechnol. 2007, 136, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Zühlke, M.-K.; Schlüter, R.; Henning, A.-K.; Lipka, M.; Mikolasch, A.; Schumann, P.; Giersberg, M.; Kunze, G.; Schauer, F. A novel mechanism of conjugate formation of bisphenol A and its analogues by Bacillus amyloliquefaciens: Detoxification and reduction of estrogenicity of bisphenols. Int. Biodeterior. Biodegrad. 2016, 109, 165–173. [Google Scholar] [CrossRef]
- Ju, J.; Shen, L.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Degradation potential of bisphenol A by Lactobacillus reuteri. LWT 2019, 106, 7–14. [Google Scholar] [CrossRef]
- Jung, W.Y.; Jung, J.Y.; Lee, H.J.; Jeon, C.O. Functional characterization of bacterial communities responsible for fermentation of Doenjang: A traditional Korean fermented soybean paste. Front. Microbiol. 2016, 7, 827. [Google Scholar] [CrossRef]
- Park, H.; Lee, M.; Jeong, D.; Park, S.; Ji, Y.; Todorov, S.D.; Holzapfel, W.H. Safety evaluation and in vivo strain-specific functionality of Bacillus strains isolated from Korean traditional fermented foods. Probiotics Antimicrob. Proteins 2021, 13, 60–71. [Google Scholar] [CrossRef]
- Mi, X.-J.; Tran, T.H.M.; Park, H.-R.; Xu, X.Y.; Subramaniyam, S.; Choi, H.S.; Kim, J.; Koh, S.C.; Kim, Y.J. Immune-enhancing effects of postbiotic produced by Bacillus velezensis Kh2-2 isolated from Korea Foods. Food Res. Int. 2022, 152, 110911. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.Y.; Daily, J.W.; Lee, G.H.; Ryu, M.S.; Yang, H.-J.; Jeong, S.-Y.; Qiu, J.Y.; Zhang, T.; Park, S. Short-term fermented soybeans with Bacillus amyloliquefaciens potentiated insulin secretion capacity and improved gut microbiome diversity and intestinal integrity to alleviate Asian type 2 diabetic symptoms. J. Agric. Food Chem. 2020, 68, 13168–13178. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Yi, S.-H.; Hong, S.-P. Characteristics of Bacterial Strains with Desirable Flavor Compounds from Korean Traditional Fermented Soybean Paste (Doenjang). Molecules 2021, 26, 5067. [Google Scholar] [CrossRef]
- Zhang, C.; Zeng, G.; Yuan, L.; Yu, J.; Li, J.; Huang, G.; Xi, B.; Liu, H. Aerobic degradation of bisphenol A by Achromobacter xylosoxidans strain B-16 isolated from compost leachate of municipal solid waste. Chemosphere 2007, 68, 181–190. [Google Scholar] [CrossRef]
- Meng, Z.; Tian, S.; Yan, J.; Jia, M.; Yan, S.; Li, R.; Zhang, R.; Zhu, W.; Zhou, Z. Effects of perinatal exposure to BPA, BPF and BPAF on liver function in male mouse offspring involving in oxidative damage and metabolic disorder. Environ. Pollut. 2019, 247, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Kimura, N.; Ikeda, I.; Fujimoto, K.; Kimoto, H. Adsorption of bisphenol A by lactic acid bacteria, Lactococcus, strains. Appl. Microbiol. Biotechnol. 2007, 74, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, N.; Shyamalagowri, S.; Nithya, T.; Aravind, J.; Govarthanan, M.; Kamaraj, M. Trends and thresholds on bacterial degradation of bisphenol A endocrine disruptor—A concise review. Environ. Monit. Assess. 2022, 194, 886. [Google Scholar] [CrossRef] [PubMed]
- Suyamud, B.; Inthorn, D.; Panyapinyopol, B.; Thiravetyan, P. Biodegradation of bisphenol A by a newly isolated Bacillus megaterium strain ISO-2 from a polycarbonate industrial wastewater. Water Air Soil Pollut. 2018, 229, 348. [Google Scholar] [CrossRef]
- Tian, K.; Yu, Y.; Qiu, Q.; Sun, X.; Meng, F.; Bi, Y.; Gu, J.; Wang, Y.; Zhang, F.; Huo, H. Mechanisms of BPA degradation and toxicity resistance in Rhodococcus equi. Microorganisms 2023, 11, 67. [Google Scholar] [CrossRef]
- Logan, N.; Vos, P. Bacillus. In Bergey’s Manual of Systematics of Archaea and Bacteria Bergey’s Manual Trust; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Gauvry, E.; Mathot, A.-G.; Couvert, O.; Leguérinel, I.; Coroller, L. Effects of temperature, pH and water activity on the growth and the sporulation abilities of Bacillus subtilis BSB1. Int. J. Food Microbiol. 2021, 337, 108915. [Google Scholar] [CrossRef]
- Li, G.; Zu, L.; Wong, P.-K.; Hui, X.; Lu, Y.; Xiong, J.; An, T. Biodegradation and detoxification of bisphenol A with one newly-isolated strain Bacillus sp. GZB: Kinetics, mechanism and estrogenic transition. Bioresour. Technol. 2012, 114, 224–230. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, K.; Chen, L. Bacteria-mediated bisphenol A degradation. Appl. Microbiol. Biotechnol. 2013, 97, 5681–5689. [Google Scholar] [CrossRef]
- Ike, M.; Chen, M.Y.; Jin, C.S.; Fujita, M. Acute toxicity, mutagenicity, and estrogenicity of biodegradation products of bisphenol A. Environ. Toxicol. Int. J. 2002, 17, 457–461. [Google Scholar] [CrossRef]
- Wang, W.; Yu, H.; Qin, H.; Long, Y.; Ye, J.; Qu, Y. Bisphenol A degradation pathway and associated metabolic networks in Escherichia coli harboring the gene encoding CYP450. J. Hazard. Mater. 2020, 388, 121737. [Google Scholar] [CrossRef]
- Jia, Y.; Eltoukhy, A.; Wang, J.; Li, X.; Hlaing, T.S.; Aung, M.M.; Nwe, M.T.; Lamraoui, I.; Yan, Y. Biodegradation of bisphenol A by Sphingobium sp. YC-JY1 and the essential role of cytochrome P450 monooxygenase. Int. J. Mol. Sci. 2020, 21, 3588. [Google Scholar] [CrossRef] [PubMed]
- Grelska, A.; Noszczyńska, M. White rot fungi can be a promising tool for removal of bisphenol A, bisphenol S, and nonylphenol from wastewater. Environ. Sci. Pollut. Res. 2020, 27, 39958–39976. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Source | BPA Degradation Performance a | Reference |
|---|---|---|---|
| B. pumilus BP-22DK | Kimchi | 10 mg/L (100%), 16 h | [21] |
| B. subtilis HV-3 | Livestock house sludge | 100 mg/L (98%), 120 h (electrochemical system) | [20] |
| B. amyloliquefaciens SBUG1837 | Activated sludge | 60 mg/L (77%), 48 h | [22] |
| B. subtilis | Soil | 50 mg/L (52%), 96 h | [18] |
| Lactococcus lactis RKG1-319 | Dairy product | 50 mg/L (39%), 96 h | |
| Lactobacillus plantarum 2035 | Dairy product | 50 mg/L (42%), 96 h | |
| Enterococcus faecalis ATCC 19433 | Lab strain | 50 mg/L (45%), 96 h | |
| L. reuteri ATCC55730 | Lab strain | 1 mg/L (70%), 48 h | [23] |
| Control (Extracellular) | Dead Cells | Intracellular | |
|---|---|---|---|
| Residual BPA (mg/L) | 0.27 ± 0.02 a | 9.88 ± 0.04 b | 0.28 ± 0.02 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.K.; Chin, Y.-W. Degradation of Bisphenol A by Bacillus subtilis P74 Isolated from Traditional Fermented Soybean Foods. Microorganisms 2023, 11, 2132. https://doi.org/10.3390/microorganisms11092132
Park YK, Chin Y-W. Degradation of Bisphenol A by Bacillus subtilis P74 Isolated from Traditional Fermented Soybean Foods. Microorganisms. 2023; 11(9):2132. https://doi.org/10.3390/microorganisms11092132
Chicago/Turabian StylePark, Young Kyoung, and Young-Wook Chin. 2023. "Degradation of Bisphenol A by Bacillus subtilis P74 Isolated from Traditional Fermented Soybean Foods" Microorganisms 11, no. 9: 2132. https://doi.org/10.3390/microorganisms11092132