

Isolation, Identification and Evaluation of the Effects of Native Entomopathogenic Fungi from Côte d’Ivoire on Galleria mellonella



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Fungi
2.3. Molecular Identification of Isolated Strains
2.4. Assessing Pathogenicity of Isolates on Galleria mellonella
2.5. Statistical Analysis
3. Results
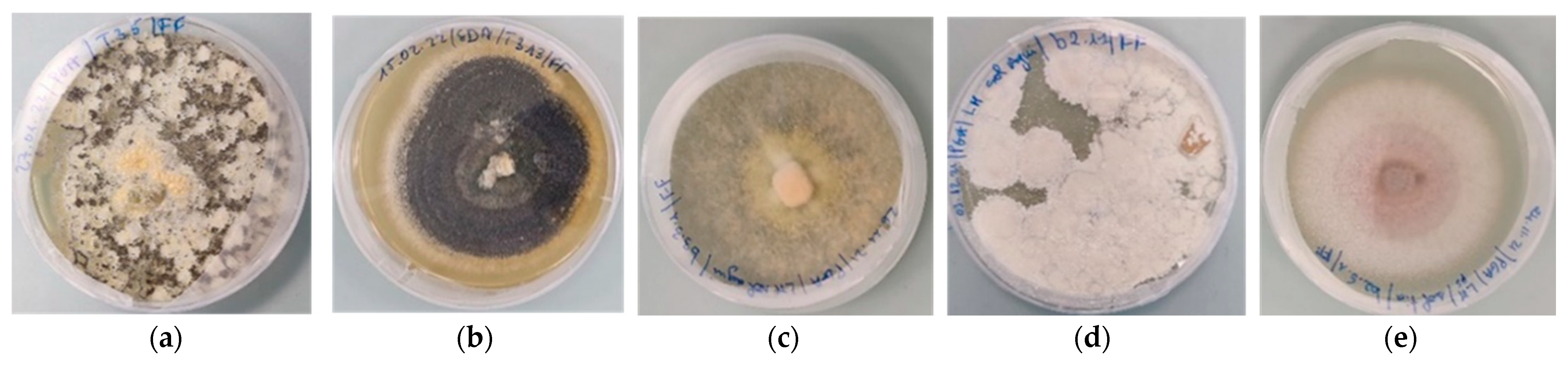
3.1. Fungi Isolated
3.2. Pathogenicity of Isolates on Galleria mellonella Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CABI. Scientists Discover New Crop-Destroying Armyworm Is Now “Spreading Rapidly” in Africa; CABI: Wallingford, UK, 2017; Available online: http://www.cabi.org/news-and-media/2017/scientistsdiscover-new-crop-destroying-armyworm-is-now-spreading-rapidly-in-africa (accessed on 10 September 2021).
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–301. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huessing, J.E.; Eddy; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management; First Edition Mexico; USAID: Washington, DC, USA; CIMMYT: El Batán, Mexico, 2018; 120p.
- Maiga, I. Note d’Informations Générales sur la Noctuelle du Maïs Spodoptera frugiperda J.E. Smith. In Un Ravageur Redoutable et très Polyphage à Surveiller; Centre Régional AGRHYMET/CILSS: Niamey, Niger, 2017; 16p. [Google Scholar]
- CILSS. La Chenille D’automne Spodoptera frugiperda, Nouveau Ravageur du Maïs en Afrique de l’Ouest, a Atteint le Niger; Bulletin Spécial; Centre Régional AGRHYMET: Niamey, Niger, 2017; 7p. [Google Scholar]
- Ruiz-Nájera, R.E.; Ruiz-Estudillo, R.A.; Sánchez-Yáñez, J.M.; Molina-Ochoa, J.; Skoda, S.R.; Coutiño-Ruiz, R.; Pinto-Ruiz, R.; Guevara-Hernández, F.; Foster, J.E. Occurrence of entomopathologic fungi and parasitic nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae collected in central Chiapes. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Hama, A.; Adamou, H.; Adamou, B.; Salifou, A.; Delams, P. Alerte: Spodoptera frugiperda, une Nouvelle Chenille Ravageur de Maïs; Note d’Information/Traitements Phytosanitaires et Ravageurs; Réseau National des Chambres d’Agriculture du Niger: Niamey, Niger, 2016; 3p. [Google Scholar]
- Tizié, Y.G.; N’Guessan, C. Atelier de Renforcement des Capacités des Etats Membres de la CEDEAO Pour Lutter Contre L’invasion de Spodoptera frugiperda en Afrique de L’ouest; Compte-Rendu d’AtelierL: Bordeaux, France, 2017; 35p. [Google Scholar]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J.; Nguyen, S.N.; Abo-Elgharg, E. Biochemical characteristics of insecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Physiol. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Bawin, T.; Seye, F.; Boukraa, S.; Zimmer, J.Y.; Delvigne, F.; Francis, F. La lutte contre les moustiques (Diptera: Culicidae): Diversité des approches et application du contrôle biologique. Can. Entomol. 2015, 147, 476–500. [Google Scholar] [CrossRef]
- Alves, S.B.; Lopes, R.B.; Vieira, S.A.; Tamai, M.A. Fungos entomopatogênicos usados no controle de pragas na América Latina. In Controle Microbiano de Pragas na América Latina: Avanços e Desafios; Alves, S.B., Ed.; FEALQ: Piracicaba, Brazil, 2008; pp. 69–110. [Google Scholar]
- Islam, M.S.; Subbiah, V.K.; Siddiquee, S. Efficacy of Entomopathogenic Trichoderma Isolates against Sugarcane Woolly Aphid, Ceratovacuna lanigera Zehntner (Hemiptera: Aphididae). Horticulturae 2022, 8, 2. [Google Scholar] [CrossRef]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Demirci, S.N.Ş.; Altuntaş, H. Entomopathogenic potential of Purpureocillium lilacinum against the model insect Galleria mellonella (Lepidoptera: Pyralidae). Environ. Exp. Biol. 2019, 17, 71–74. [Google Scholar]
- Thaochan, N.; Sausa-Ard, W. Occurrence and effectiveness of indigenous Metarhizium anisopliae against adults Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae) in Southern Thailand. Songklanakarin. J. Sci. Technol. 2017, 39, 325–334. [Google Scholar]
- Davet, P.; Rouxel, F. Detection and Isolation of Fungi from the Soil; Edition INRA: Paris, France, 1997; p. 194. [Google Scholar]
- Zimmerman, G. The Galleria bait method’ for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Laurent, L.; Crovadore, J.; Deneux, P.; Cochard, B.; Ravat, P.C.; Chablais, R.; Bovigny, P.Y.; Lefort, F. New strains of entomopathogenic fungi isolated from organic orchards in Geneva. Commun. Agric. Appl. Biol. 2017, 82, 207–212. [Google Scholar]
- Ripoll, J.; Bon, M.C.; Jones, W. Optimization of genomic DNA extraction from yellow nightshade (Solanum elaeagnifolium Cav.), an invasive plant in cultivated areas in the Mediterranean region. Biotechnol. Agron. Soc. Environ. 2011, 15, 95–100. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Sohrabi, F.; Jamali, F.; Morammazi, S.; Saber, M.; Kamita, S.G. Evaluation of the compatibility of entomopathogenic fungi and two botanical insecticides tondexir and palizin for controlling Galleria mellonella L. (Lepidoptera: Pyralidae). Crop Prot. 2019, 117, 20–25. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Bojke, A.; Tkaczuk, C. Effects of the entomopathogenic fungi Metarhizium robertsii, Metarhizium flavoviride, and Isaria fumosorosea on the lipid composition of Galleria mellonella larvae. Mycologia 2021, 113, 525–535. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Sabbahi, R.; Merzouki, A.; Guertin, C. Efficacy of Beauveria bassiana (Bals.) Vuill. Against the tarnished plant bug, Lygus lineolaris L., in strawberries. J. Appl. Entomol. 2008, 132, 124–134. [Google Scholar] [CrossRef]
- Sengul, A.K.; Serdar, U. Isolation, identification and seasonal distribution of soilborne fungi in tea growing areas of Iyidere-Ikizdere vicinity (Rize-Turkey). J. Basic Microbiol. 2006, 46, 208–218. [Google Scholar]
- Ibrahim, A.A.; Mohamed, H.F.; El-Naggar, S.E.M.; Swelim, M.A.; Elkhawaga, O.E. Isolation and selection of entomopathogenic fungi as biocontrol agent against the Greater Wax Moth, Galleria mellonella L. (Lepidoptera: Pyralidae). Egypt. J. Biol. Pest Control 2016, 26, 249–253. [Google Scholar]
- Correa, T.A.; Santos, F.S.; Camargo, M.G.; Quinelato, S.; Bittencourt, V.R.E.P.; Golo, P.S. Comparison of methods for isolating entomopathogenic fungi from soil samples. JOVE 2022, 179, e63353. [Google Scholar]
- Meyling, N.V. Methods for Isolation of Entomopathogenic Fungi from the Soil Environment, Laboratory Manual. Deliverable 5.1, VegQure, DARCOF III: Research in Organic Food and Farming (FØJO III). 2007. Available online: https://orgprints.org/id/eprint/11200/ (accessed on 1 April 2022).
- Baydar, R.; Güven, Ö.; Karaca, I. Occurrence of Entomopathogenic Fungi in Agriculture Soils Isparta Provence in Turkey and their Pathogenicity to Galleria mellonella (L.) (Lepidoptera: Pyralidae) larvae. Egypt. J. Biol. Pest Control 2016, 26, 323–327. [Google Scholar]
- Saleh, M.M.E.; Abdel-Raheem, M.A.; Ebadah, I.M.; Huda, H.E. Natural abundance of entomopathogenic fungi in fruit orchards and their virulence against Galleria mellonella larvae. Egypt. J. Biol. Pest Control 2016, 26, 203–207. [Google Scholar]
- Klingen, I.; Meadow, R.; Aandal, T. Mortality of Delia floralis, Galleria mellonella and Mamestra brassicae treated with insect pathogenic hyphomycetous fungi. J. Appl. Entomol. 2002, 126, 231–237. [Google Scholar] [CrossRef]
- Rajula, J.; Pittarate, S.; Suwannarach, N.; Kumla, J.; Ptaszynska, A.A.; Thungrabeab, M.; Mekchay, S.; Krutmuang, P. Evaluation of native entomopathogenic fungi for the control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A sustainable way for eco-friendly agriculture. J. Fungi 2021, 7, 1073. [Google Scholar] [CrossRef] [PubMed]
- Alfiky, A. Screening and identification of indigenous entomopathogenic fungal isolates from agricultural farmland soils in Nile Delta, Egypt. J. Fungi 2022, 8, 54. [Google Scholar] [CrossRef]
- Oreste, M.; Bubici, G.; Poliseno, M.; Triggiani, O.; Tarasco, E. Pathogenicity of Beauveria bassiana (Bals.-Criv.) Vuill. and Metarhizium anisopliae (Metschn.) Sorokin against Galleria mellonella L. and Tenebrio molitor L. in laboratory assays. Redia 2012, 95, 43–48. [Google Scholar]
- Castrillo, L.A.; Griggs, M.H.; Liu, H.; Bauer, L.S.; Vandenberg, J.D. Assessing deposition and persistence of Beauveria bassiana GHA (Ascomycota: Hypocreales) applied for control of the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae), in a commercial tree nursery. Biol. Control 2010, 54, 61–67. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Isolates | Accessions Numbers |
|---|---|---|
| B. bassiana | A211 | ON121958 |
| B. bassiana | A214a | ON121959 |
| B. bassiana | A214b | ON121960 |
| M. anisopliae | T35 | ON121962 |
| M. anisopliae | T331 | ON121963 |
| Metarhizium sp. | T141 | ON121961 |
| Metarhizium sp. | T34 | N.A. |
| Metarhizium sp. | T121 | N.A. |
| Metarhizium sp. | T132 | N.A. |
| Metarhizium sp. | T313 | N.A. |
| Trichoderma asperellum | Ou5 | ON121957 |
| Trichoderma longibrachiatum | Ko4 | ON121954 |
| Species | Isolates | UASWS’s Numbers | Number of Larvae Tested | Number of Dead Larvae | Re-Isolation Rates (%) |
|---|---|---|---|---|---|
| B. bassiana | A211 | UASWS2633 | 30 | 30 | 90 |
| B. bassiana | A214a | UASWS2634 | 30 | 30 | 100 |
| B. bassiana | A214b | UASWS2635 | 30 | 30 | 100 |
| Metarhizium sp. | T34 | N.A. | 30 | 27 | 80 |
| M. anisopliae | T35 | UASWS2637 | 30 | 28 | 100 |
| Metarhizium sp. | T121 | N.A. | 30 | 27 | 100 |
| Metarhizium sp. | T132 | N.A. | 30 | 21 | 100 |
| Metarhizium sp. | T141 | UASWS2636 | 30 | 24 | 70 |
| Metarhizium sp. | T313 | N.A. | 30 | 26 | 90 |
| M. anisopliae | T331 | UASWS2638 | 30 | 30 | 90 |
| T. longibrachiatum | Ko4 | UASWS2629 | 30 | 5 | 0 |
| T. asperellum | Ou5 | UASWS2632 | 30 | 3 | 0 |
| Fungi | Isolates | LC50 (Conidia mL−1) | LT50 (Days) |
|---|---|---|---|
| B. bassiana | A214b | 1.12 × 104 | 2.64 |
| M. anisopliae | T331 | 3.47 × 104 | 3.08 |
| Metarhizium sp. | T34 | 2.19 × 105 | 3.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fofana, F.; Descombes, C.; Kouamé, A.P.; Lefort, F. Isolation, Identification and Evaluation of the Effects of Native Entomopathogenic Fungi from Côte d’Ivoire on Galleria mellonella. Microorganisms 2023, 11, 2104. https://doi.org/10.3390/microorganisms11082104
Fofana F, Descombes C, Kouamé AP, Lefort F. Isolation, Identification and Evaluation of the Effects of Native Entomopathogenic Fungi from Côte d’Ivoire on Galleria mellonella. Microorganisms. 2023; 11(8):2104. https://doi.org/10.3390/microorganisms11082104
Chicago/Turabian StyleFofana, Fatoumatou, Corentin Descombes, Assiri Patrice Kouamé, and François Lefort. 2023. "Isolation, Identification and Evaluation of the Effects of Native Entomopathogenic Fungi from Côte d’Ivoire on Galleria mellonella" Microorganisms 11, no. 8: 2104. https://doi.org/10.3390/microorganisms11082104