
Genomic Analysis of Clostridioides difficile Recovered from Horses in Western Australia
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. Case Study Samples
2.3. Culture
2.4. C. difficile Characterisation
2.5. Antimicrobial Susceptibility Testing (AST)
2.6. Whole-Genome Sequencing (WGS) and Analysis
2.7. ST54 Whole-Genome SNP Phylogeny
2.8. Hybrid Assembly of Equine C. difficile RT 012 Closed Reference Genome
2.9. Statistical Analysis
3. Results
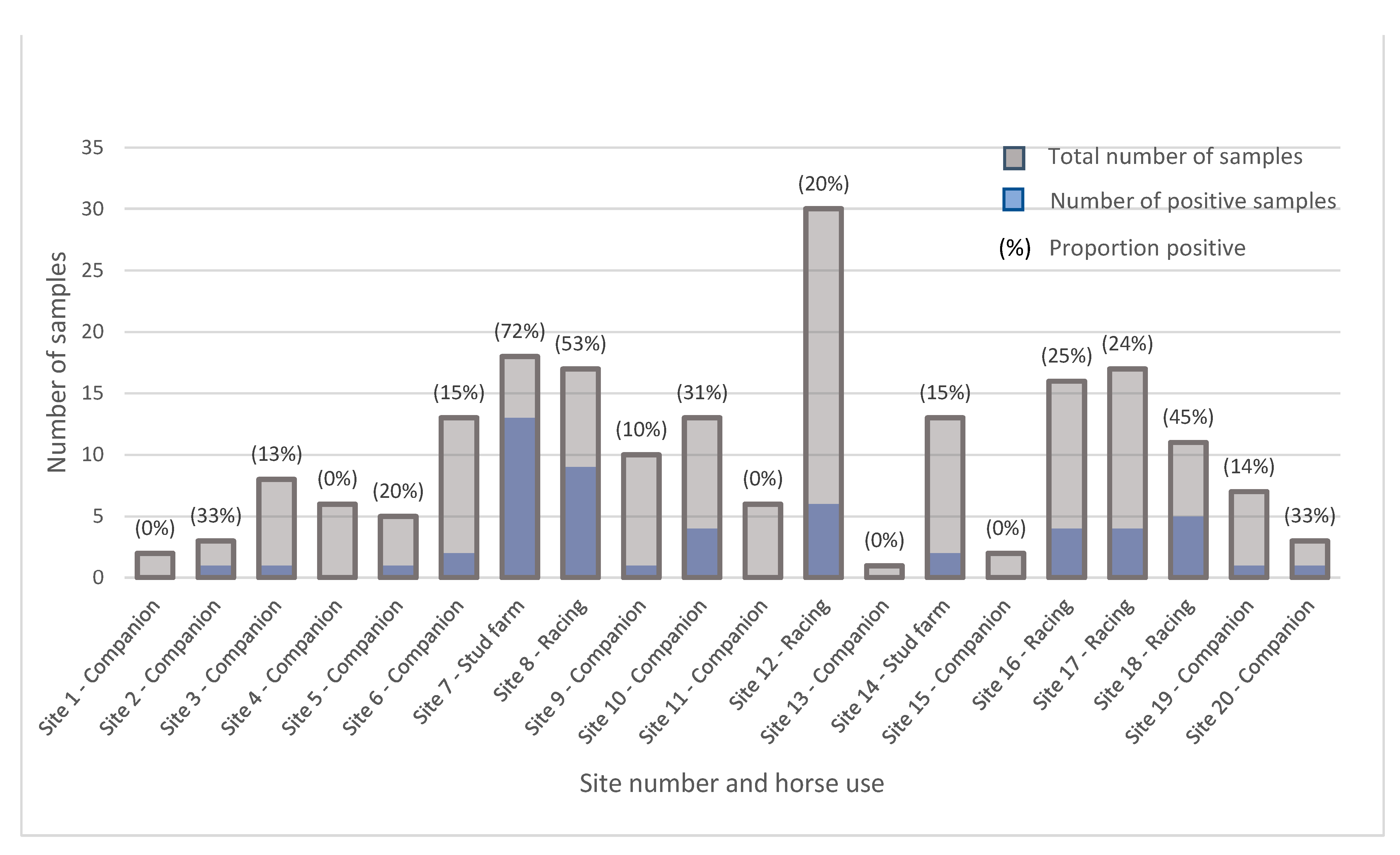
3.1. Epidemiology
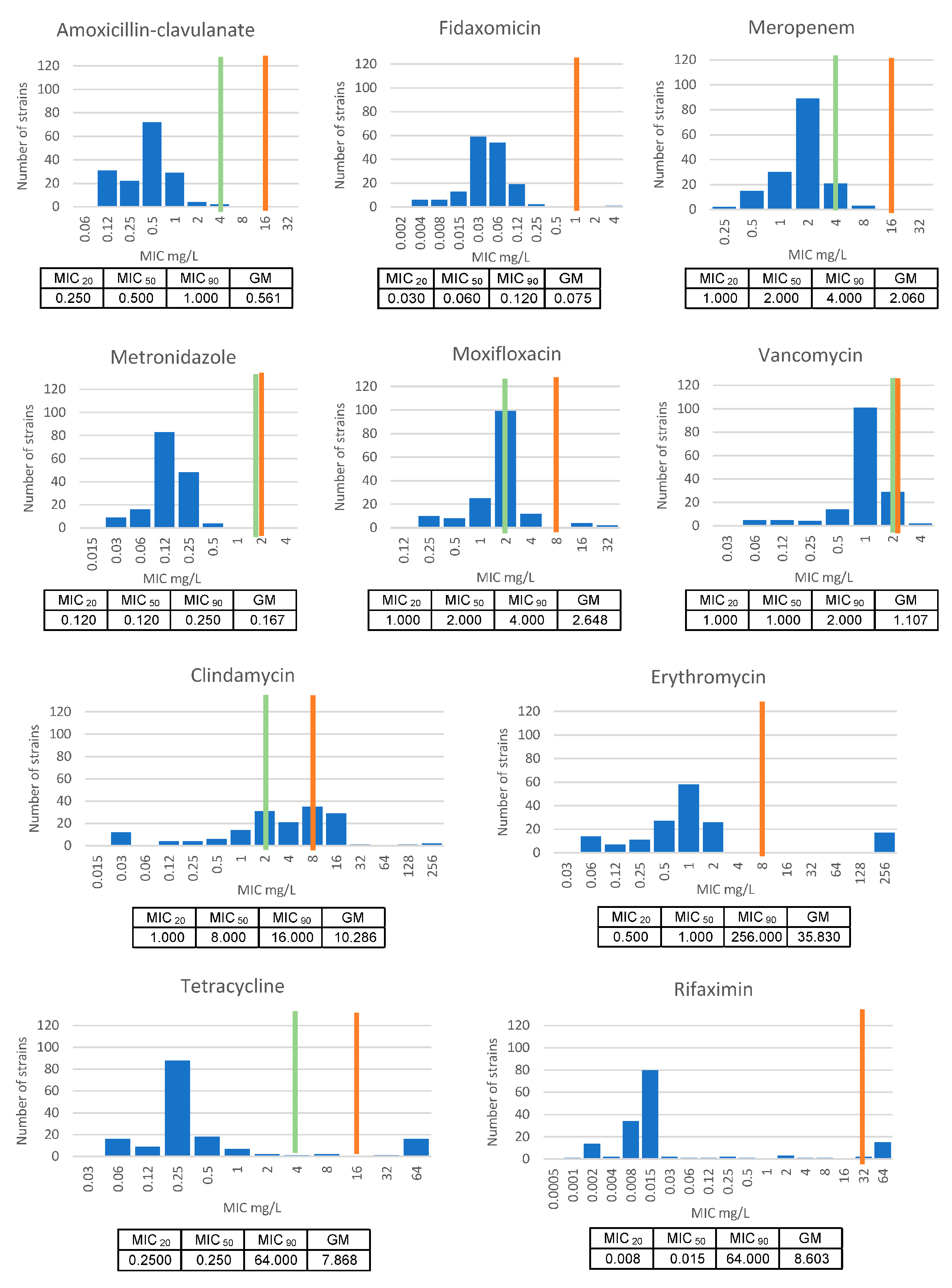
3.2. Antimicrobial Susceptibility Testing (AST)
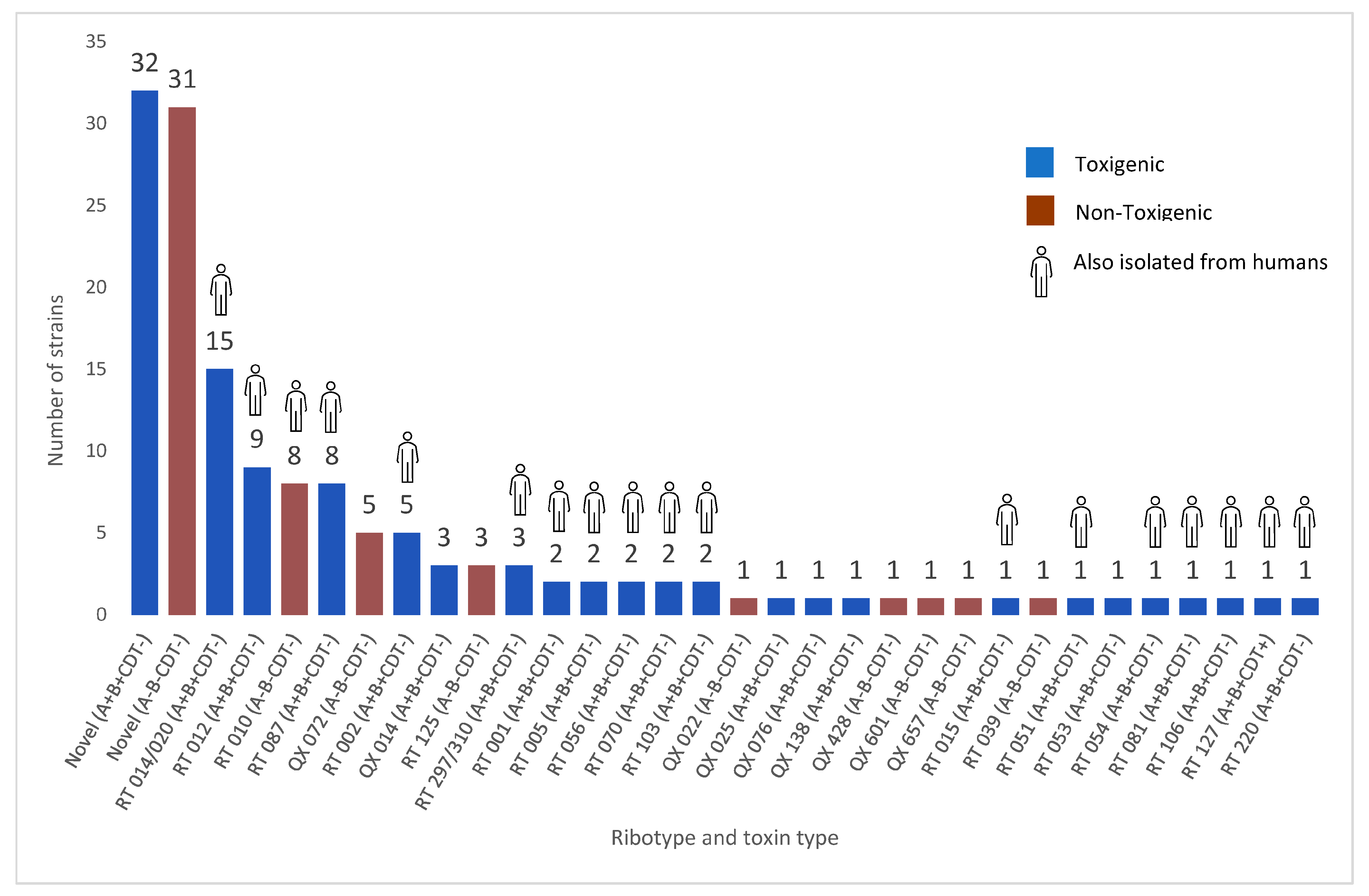
3.3. Molecular Characterisation
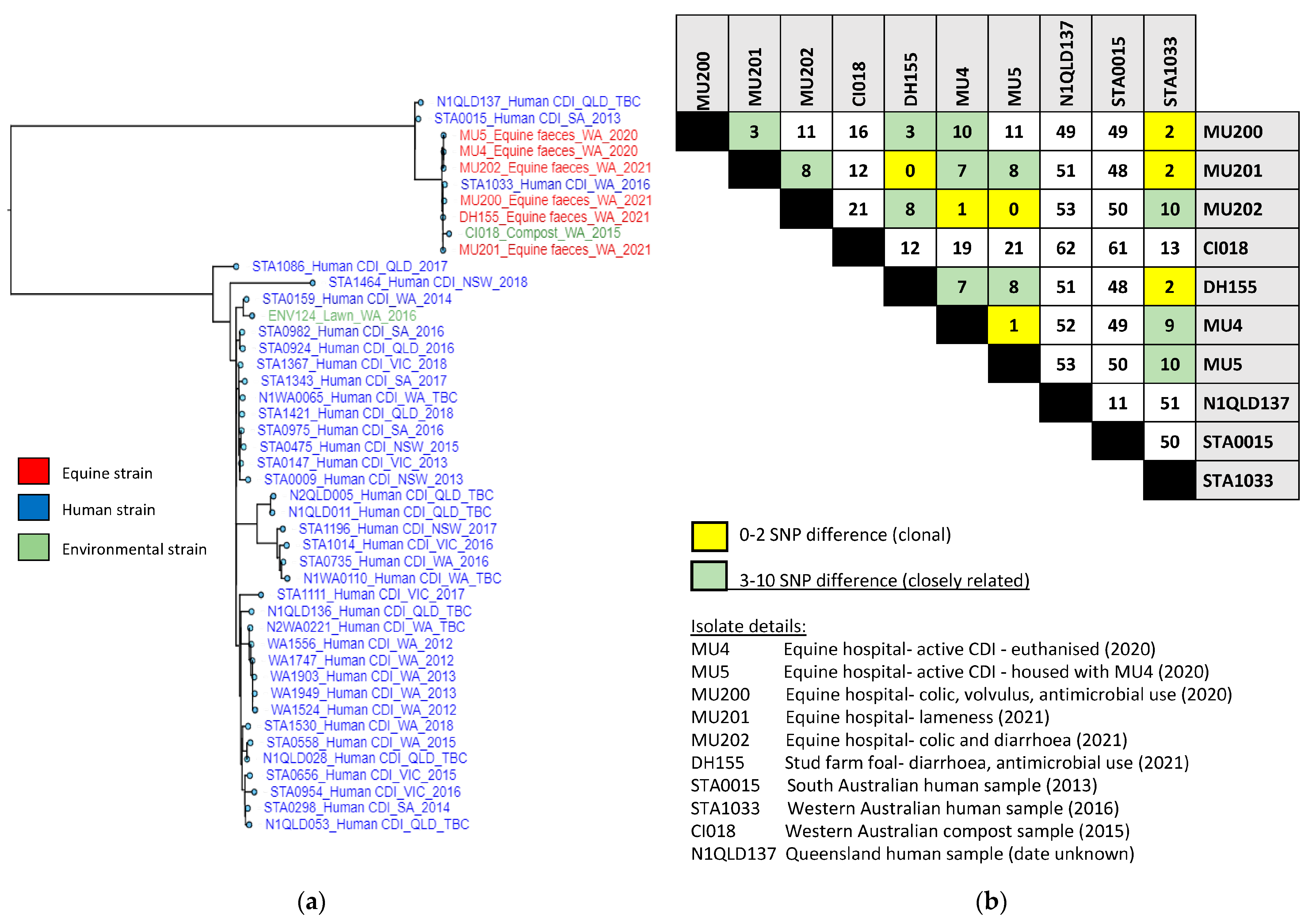
3.4. ST54 Whole-Genome SNP Phylogeny
3.5. Complete Genome for MU4
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ofori, E.; Ramai, D.; Dhawan, M.; Mustafa, F.; Gasperino, J.; Reddy, M. Community-acquired Clostridium difficile: Epidemiology, ribotype, risk factors, hospital and intensive care unit outcomes, and current and emerging therapies. J. Hosp. Infect. 2018, 99, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Feuerstadt, P.; Theriault, N.; Tillotson, G. The burden of CDI in the United States: A multifactorial challenge. BMC Infect. Dis. 2023, 23, 132. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 28 February 2023).
- Keel, M.K.; Songer, J.G. The comparative pathology of Clostridium difficile-associated disease. Vet. Path 2006, 43, 225–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, A. Clostridium difficile toxins: Mediators of inflammation. J. Innate Immun. 2012, 4, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyerly, D.M.; Lockwood, D.E.; Richardson, S.H.; Wilkins, T.D. Biological activities of toxins A and B of Clostridium difficile. Infect. Immun. 1982, 35, 1147–1150. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, S.A.; Cartman, S.T.; Heap, J.T.; Kelly, M.L.; Cockayne, A.; Minton, N.P. The role of toxin A and toxin B in Clostridium difficile infection. Nature 2010, 467, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Buddle, J.E.; Fagan, R.P. Pathogenicity and virulence of Clostridioides difficile. Virulence 2023, 14, 2150452. [Google Scholar] [CrossRef]
- Weese, J.S. Clostridium (Clostridioides) difficile in animals. J. Vet. Diagn. Investig. Off. Publ. Am. Assoc. Vet. Lab. Diagn. Inc. 2020, 32, 213–221. [Google Scholar] [CrossRef]
- Knight, D.R.; Riley, T.V. Genomic delineation of zoonotic origins of Clostridium difficile. Front. Public Health 2019, 7, 164. [Google Scholar] [CrossRef] [Green Version]
- Knetsch, C.W.; Connor, T.; Mutreja, A.; van Dorp, S.M.; Sanders, I.; Browne, H.; Harris, D.; Lipman, L.; Keessen, E.C.; Corver, J.; et al. Whole genome sequencing reveals potential spread of Clostridium difficile between humans and farm animals in the Netherlands, 2002 to 2011. Eurosurveillance 2014, 19, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Loo, V.G.; Brassard, P.; Miller, M.A. Household transmission of Clostridium difficile to family members and domestic pets. Infect. Control. Hosp. Epidemiol. 2016, 37, 1342. [Google Scholar] [CrossRef] [PubMed]
- Redding, L.E.; Habing, G.G.; Tu, V.; Bittinger, K.L.; O’Day, J.; Pancholi, P.; Wang, S.-H.; Alexander, A.; Kelly, B.J.; Weese, J.S.; et al. Infrequent intrahousehold transmission of Clostridioides difficile between pet owners and their pets. Zoonoses Public Health 2023, 70, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Diab, S.S.; Rodriguez-Bertos, A.; Uzal, F.A. Pathology and diagnostic criteria of Clostridium difficile enteric infection in horses. Vet. Pathol. 2013, 50, 1028–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Båverud, V.; Gustafsson, A.; Franklin, A.; Aspán, A.; Gunnarsson, A. Clostridium difficile: Prevalence in horses and environment, and antimicrobial susceptibility. Equine Vet. J. 2003, 35, 465–471. [Google Scholar] [CrossRef]
- Thean, S.; Elliott, B.; Riley, T.V. Clostridium difficile in horses in Australia—A preliminary study. J. Med. Microbiol. 2011, 60, 1188–1192. [Google Scholar] [CrossRef]
- Weese, J.S.; Staempfli, H.R.; Prescott, J.F. A prospective study of the roles of Clostridium difficile and enterotoxigenic Clostridium perfringens in equine diarrhoea. Equine Vet. J. 2001, 33, 403–409. [Google Scholar] [CrossRef]
- Shaughnessy, M.; Snider, T.; Sepulveda, R.; Boxrud, D.; Cebelinski, E.; Jawahir, S.; Holzbauer, S.; Johnston, B.; Smith, K.; Bender, J.; et al. Prevalence and molecular characteristics of Clostridium difficile in retail meats, food-producing and companion animals, and humans in Minnesota. J. Food Prot. 2018, 81, 1635–1642. [Google Scholar] [CrossRef]
- Schoster, A.; Staempfli, H.R. Epidemiology and antimicrobial resistance in Clostridium difficile with special reference to the horse. Curr. Clin. Microbiol. Rep. 2016, 3, 32–41. [Google Scholar] [CrossRef]
- Racing Australia. Racing Australia Annual Report 2020; Racing Australia Limited: Flemington, Australia, 2020. [Google Scholar]
- Helgi Library. Horse Meat Production. Available online: https://www.helgilibrary.com/indicators/horse-meat-production/ (accessed on 21 February 2023).
- Lim, S.C.; Knight, D.R.; Moono, P.; Foster, N.F.; Riley, T.V. Clostridium difficile in soil conditioners, mulches and garden mixes with evidence of a clonal relationship with historical food and clinical isolates. Environ. Microbiol. Rep. 2020, 12, 672–680. [Google Scholar] [CrossRef]
- Brazier, J.S. Role of the laboratory in investigations of Clostridium difficile diarrhea. Clin. Infect. Dis. 1993, 16, 228–233. [Google Scholar] [CrossRef]
- Lim, S.C.; Foster, N.F.; Elliott, B.; Riley, T.V. High prevalence of Clostridium difficile on retail root vegetables, Western Australia. J. Appl. Microbiol. 2018, 124, 585–590. [Google Scholar] [CrossRef]
- M11-A7; Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria, Seventh Edition. Clinical and Laboratory Standards Institute, CLSI: Wayne, PA, USA, 2011.
- The European Committee on Antimicrobial Susceptibility Testing, Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 13.0. 2023. Available online: http://www.eucast.org (accessed on 1 April 2023).
- EMA/857570/2011; Assessment Report, Dificlir Fidaxomicin. EMA, European Medicines Agency: Amsterdam, The Netherlands, 2011. Available online: https://www.ema.europa.eu (accessed on 1 April 2023).
- O’Connor, J.R.; Galang, M.A.; Sambol, S.P.; Hecht, D.W.; Vedantam, G.; Gerding, D.N.; Johnson, S. Rifampin and rifaximin resistance in clinical isolates of Clostridium difficile. Antimicrob. Agents Chemother. 2008, 52, 2813–2817. [Google Scholar] [CrossRef] [Green Version]
- M100-S23; Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Third Informational Supplement. CLSI, Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013.
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.W.; Fung, R.; Golubchik, T.; Harding, R.M.; Jeffery, K.J.; Jolley, K.A.; et al. Multilocus sequence typing of Clostridium difficile. J. Clin. Microbiol. 2010, 48, 770–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.R. SKA: Split Kmer analysis toolkit for bacterial genomic epidemiology. bioRxiv 2018, 453142. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Fawley, W.N.; Knetsch, C.W.; MacCannell, D.R.; Harmanus, C.; Du, T.; Mulvey, M.R.; Paulick, A.; Anderson, L.; Kuijper, E.J.; Wilcox, M.H. Development and validation of an internationally-standardized, high-resolution capillary gel-based electrophoresis PCR-ribotyping protocol for Clostridium difficile. PLoS ONE 2015, 10, e0118150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weese, J.S.; Rousseau, J.; Deckert, A.; Gow, S.; Reid-Smith, R.J. Clostridium difficile and methicillin-resistant Staphylococcus aureus shedding by slaughter-age pigs. BMC Vet. Res. 2011, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Torres, C.E.; Weese, J.S.; Staempfli, H.R. Validation of a commercial enzyme immunoassay for detection of Clostridium difficile toxins in feces of horses with acute diarrhea. J. Vet. Intern. Med. 2010, 24, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.C.; McGrath, J.W.; McMullan, G.; Marks, N.; Fairley, D.J. Development of an optimized broth enrichment culture medium for the isolation of Clostridium difficile. Anaerobe 2018, 54, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Dharmasena, M.; Jiang, X. Improving culture media for the isolation of Clostridium difficile from compost. Anaerobe 2018, 51, 1–7. [Google Scholar] [CrossRef]
- Blanco, J.L.; Álvarez-Pérez, S.; García, M.E. Is the prevalence of Clostridium difficile in animals underestimated? Vet. J. 2013, 197, 694–698. [Google Scholar] [CrossRef]
- Weese, J.S.; Avery, B.P.; Rousseau, J.; Reid-Smith, R.J. Detection and enumeration of Clostridium difficile spores in retail beef and pork. Appl. Environ. Microbiol. 2009, 75, 5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawley, T.D.; Clare, S.; Deakin, L.J.; Goulding, D.; Yen, J.L.; Raisen, C.; Brandt, C.; Lovell, J.; Cooke, F.; Clark, T.G. Use of purified Clostridium difficile spores to facilitate evaluation of health care disinfection regimens. Appl. Environ. Microbiol. 2010, 76, 6895–6900. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.; Taminiau, B.; Brévers, B.; Avesani, V.; Van Broeck, J.; Leroux, A.A.; Amory, H.; Delmée, M.; Daube, G. Carriage and acquisition rates of Clostridium difficile in hospitalized horses, including molecular characterization, multilocus sequence typing and antimicrobial susceptibility of bacterial isolates. Vet. Microbiol. 2014, 172, 309–317. [Google Scholar] [CrossRef]
- Schoster, A.; Staempfli, H.R.; Arroyo, L.G.; Reid-Smith, R.J.; Janecko, N.; Shewen, P.E.; Weese, J.S. Longitudinal study of Clostridium difficile and antimicrobial susceptibility of Escherichia coli in healthy horses in a community setting. Vet. Microbiol. 2012, 159, 364–370. [Google Scholar] [CrossRef]
- Bandelj, P.; Blagus, R.; Briski, F.; Frlic, O.; Vergles Rataj, A.; Rupnik, M.; Ocepek, M.; Vengust, M. Identification of risk factors influencing Clostridium difficile prevalence in middle-size dairy farms. Vet. Res. 2016, 47, 41. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Sebaihia, M.; Lawley, T.D.; Stabler, R.A.; Dawson, L.F.; Martin, M.J.; Holt, K.E.; Seth-Smith, H.M.B.; Quail, M.A.; Rance, R.; et al. Evolutionary dynamics of Clostridium difficile over short and long time scales. Proc. Natl. Acad. Sci. USA 2010, 107, 7527–7532. [Google Scholar] [CrossRef]
- Brouwer, M.S.M.; Roberts, A.P.; Hussain, H.; Williams, R.J.; Allan, E.; Mullany, P. Horizontal gene transfer converts non-toxigenic Clostridium difficile strains into toxin producers. Nat. Commun. 2013, 4, 2601. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, M.; Walk, S.T.; Young, V.B.; Aronoff, D.M. A clinical and epidemiological review of non-toxigenic Clostridium difficile. Anaerobe 2013, 22, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Gerding, D.N.; Sambol, S.P.; Johnson, S. Non-toxigenic Clostridioides (Formerly Clostridium) difficile for prevention of C. difficile infection: From bench to bedside back to bench and back to bedside. Front. Microbiol. 2018, 9, 1700. [Google Scholar] [CrossRef] [PubMed]
- Sambol, S.P.; Skinner, A.M.; Serna-Perez, F.; Owen, B.; Gerding, D.N.; Johnson, S. Effective colonization by nontoxigenic Clostridioides difficile REA strain M3 (NTCD-M3) spores following treatment with either Fidaxomicin or Vancomycin. Microbiol. Spectr. 2023, 11, e0051723. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.R.; Squire, M.M.; Collins, D.A.; Riley, T.V. Genome analysis of Clostridium difficile PCR ribotype 014 lineage in Australian pigs and humans reveals a diverse genetic repertoire and signatures of long-range interspecies Transmission. Front. Microbiol. 2016, 7, 2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moono, P.; Lim, S.C.; Riley, T.V. High prevalence of toxigenic Clostridium difficile in public space lawns in Western Australia. Sci. Rep. 2017, 7, 41196. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Putsathit, P.; George, N.; Hemphill, C.; Huntington, P.G.; Korman, T.M.; Kotsanas, D.; Lahra, M.; McDougall, R.; Moore, C.V.; et al. Laboratory-based surveillance of Clostridium difficile infection in Australian health care and community settings, 2013 to 2018. J. Clin. Microbiol. 2020, 58, e01552-20. [Google Scholar] [CrossRef]
- Collins, D.A.; Riley, T.V. Clostridium difficile in Asia: Opportunities for One Health management. Trop. Med. Infect. Dis. 2019, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.; Vernon, J.; Morris, K.; Nicholson, S.; Todhunter, S.; Longshaw, C.; Wilcox, M.H. Pan-European longitudinal surveillance of antibiotic resistance among prevalent Clostridium difficile ribotypes. Clin. Microbiol. Infect. 2015, 21, 248.e9–248.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weese, J.S.; Slovis, N.; Rousseau, J. Clostridioides (Clostridium) difficile in neonatal foals and mares at a referral hospital. J. Vet. Intern. Med. 2021, 35, 1140–1146. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Lee, K.; Byun, J.-W.; Kim, H.; So, B.; Ku, B.-K.; Kim, H.-Y.; Moon, B.-Y. Prevalence, genetic characteristics, and antimicrobial resistance of Clostridioides difficile isolates from horses in Korea. Anaerobe 2023, 80, 102700. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.R.; Kullin, B.; Androga, G.O.; Barbut, F.; Eckert, C.; Johnson, S.; Spigaglia, P.; Tateda, K.; Tsai, P.J.; Riley, T.V. Evolutionary and genomic insights into Sequence Type 11: A diverse zoonotic and antimicrobial-resistant lineage of global One Health importance. mBio 2019, 10, e00446-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janezic, S.; Garneau, J.R.; Monot, M. Comparative Genomics of Clostridium difficile. In Updates on Clostridium Difficile in Europe: Advances in Microbiology, Infectious Diseases and Public Health Volume 8; Mastrantonio, P., Rupnik, M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 59–75. [Google Scholar]
- Imwattana, K.; Kiratisin, P.; Riley, T.V.; Knight, D.R. Genomic basis of antimicrobial resistance in non-toxigenic Clostridium difficile in Southeast Asia. Anaerobe 2020, 66, 102290. [Google Scholar] [CrossRef]
- Hardefeldt, L.Y.; Gilkerson, J.R.; Billman-Jacobe, H.; Stevenson, M.A.; Thursky, K.; Bailey, K.E.; Browning, G.F. Barriers to and enablers of implementing antimicrobial stewardship programs in veterinary practices. J. Vet. Intern. Med. 2018, 32, 1092–1099. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Origin | Horses (N) | C. difficile +ve Horses | Proportion +ve (%) |
|---|---|---|---|
| Veterinary Hospital: | 186 | 68 | 36.6% |
| No GI signs | 142 | 52 | 36.6% |
| Foals 4–12 months | 0 | n/a | n/a |
| Adults > 12 months | 142 | 52 | 36.6% |
| GI signs | 44 | 16 | 36.4% |
| Foals 4–12 months | 0 | n/a | n/a |
| Adults > 12 months | 44 | 16 | 36.4% |
| Private domestic: | 201 | 55 | 27.4% |
| No GI signs | 194 | 52 | 26.8% |
| Foals 4–12 months | 15 | 7 | 46.7% |
| Adults > 12 months | 179 | 45 | 25.1% |
| GI signs | 8 | 3 | 37.5% |
| Foals 4–12 months | 5 | 3 | 60.0% |
| Adults > 12 months | 3 | 0 | 0% |
| Overall totals: | 387 | 123 | 31.8% |
| No GI signs | 336 | 104 | 30.9% |
| Foals 4–12 months | 15 | 7 | 46.7% |
| Adults > 12 months | 321 | 97 | 30.2% |
| GI signs | 51 | 19 | 37.2% |
| Foals 4–12 months | 5 | 3 | 60.0% |
| Adults > 12 months | 46 | 16 | 34.8% |
| Isolate MU4 | Features |
|---|---|
| Origin | Domestic Horse, Western Australia, 2020 |
| PCR ribotype | 012 |
| Multi-locus sequence type | 54 (clade C1) |
| Toxin profile | A+B+CDT- |
| Chromosome | |
| GenBank accession | CP102397.1 |
| Size (bp) | 4,492,337 |
| %GC | 29.29 |
| N CDS | 4079 |
| N tRNA/rRNA | 50/6 |
| N CRISPRs | 9 |
| Prophages | Clostridium Phage CDMH1 Clostridium Phage ΦMMP02 Clostridium Phage ΦCDHM19 |
| No. genes | 4309 |
| No. coding sequences | 4180 |
| Noncoding RNAs | 4 |
| AMR loci | Beta Lactam (blaCDD-1) Vancomycin (vanR-Cd, vanS-Cd, vanG-Cd, vanT-Cd, vanZ1) Macrolide (erm(B)) Streptomycin (aadE, ant(6)-Ia, sat4) Tetracycline (tet(M)) Amikacin; kanamycin (aph(3’)-IIIa) Trimethoprim (dfrF) (95.96% coverage) Amikacin; gentamicin; kanamycin; tobramycin (aph(2”)-Ih) (82.21% coverage) |
| Extrachromosomal features | |
| Novel plasmid | 7881 bp, accession CP102399.1 |
| Novel phage | 34,753 bp, accession CP102398.1 |
| Total genome size (bp) | 4,535,071 |
| N contigs | 3 |
| Total ONT reads | 978,529 |
| Average ONT read length (bp) | 3610 |
| Basecalled N50 | 11,200 |
| Total illumina reads | 2,079,414 |
| SRA accession | SAMN30090499 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hain-Saunders, N.M.R.; Knight, D.R.; Bruce, M.; Byrne, D.; Riley, T.V. Genomic Analysis of Clostridioides difficile Recovered from Horses in Western Australia. Microorganisms 2023, 11, 1743. https://doi.org/10.3390/microorganisms11071743
Hain-Saunders NMR, Knight DR, Bruce M, Byrne D, Riley TV. Genomic Analysis of Clostridioides difficile Recovered from Horses in Western Australia. Microorganisms. 2023; 11(7):1743. https://doi.org/10.3390/microorganisms11071743
Chicago/Turabian StyleHain-Saunders, Natasza M. R., Daniel R. Knight, Mieghan Bruce, David Byrne, and Thomas V. Riley. 2023. "Genomic Analysis of Clostridioides difficile Recovered from Horses in Western Australia" Microorganisms 11, no. 7: 1743. https://doi.org/10.3390/microorganisms11071743