
Clostridioides difficile, a New “Superbug”


Abstract
:1. Introduction
2. Material and Methods
3. Discussion
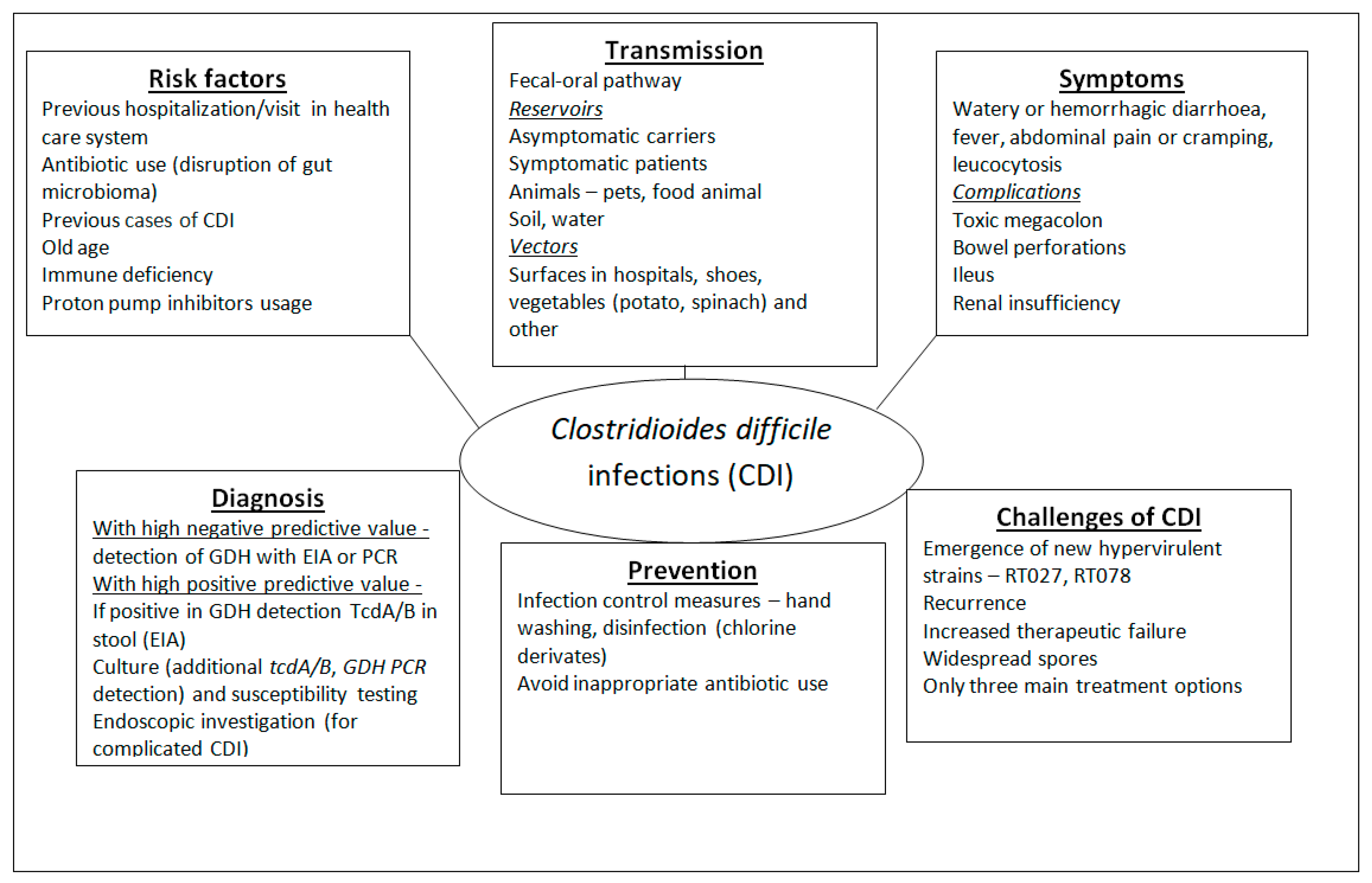
3.1. Factors of Virulence and Pathogenesis
3.2. C. difficile Infections (CDIs) and Associated Risk Factors
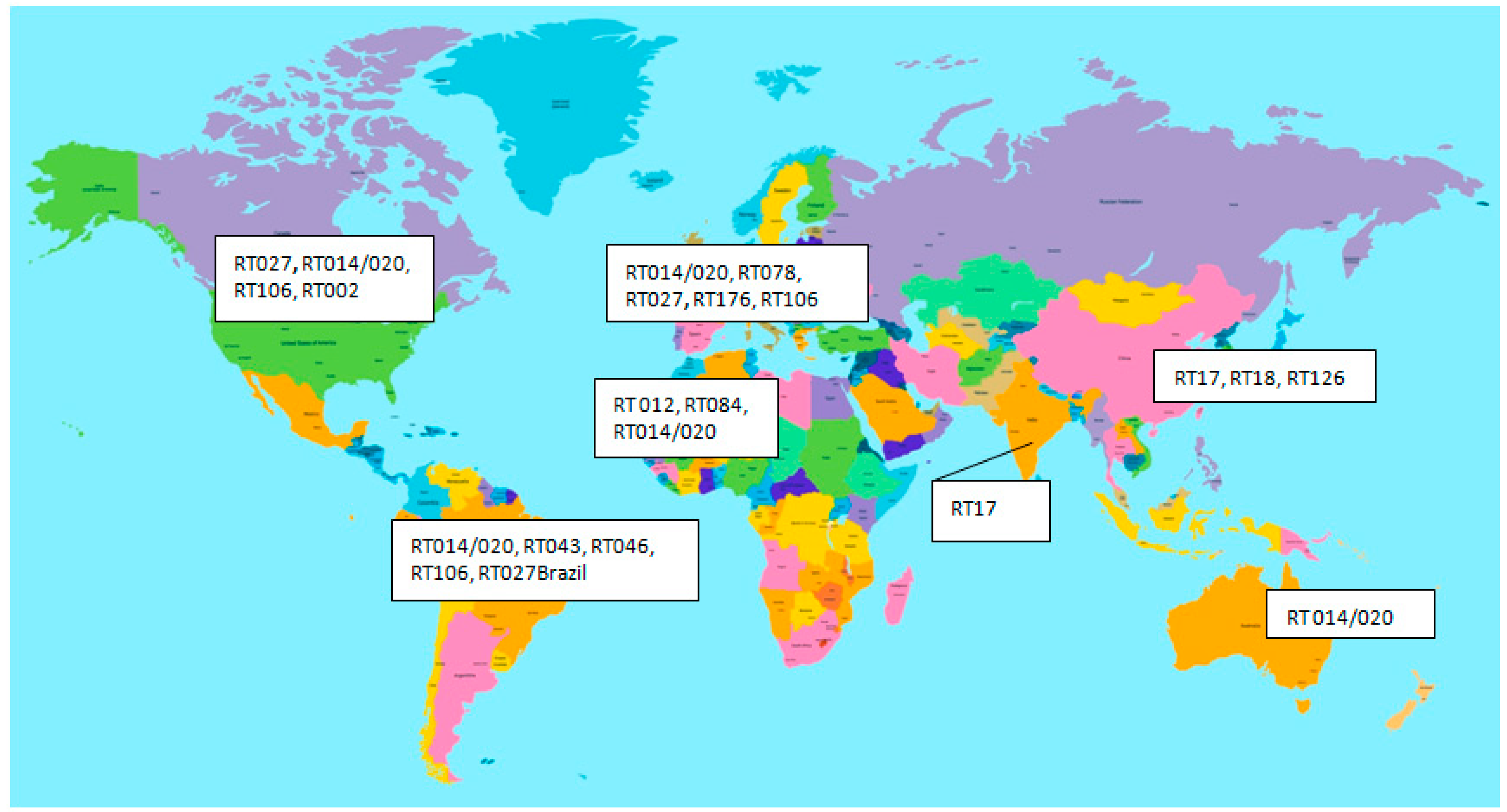
3.3. Epidemiology
3.4. CDI Treatment
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hall, I.C.; O’Toole, E. Intestinal flora in newborn infants with description of a new pathogenic anaerobe. Am. J. Dis. Child. 1935, 49, 390–402. [Google Scholar] [CrossRef]
- Tedesco, F.J.; Barton, R.W.; Alpers, D.H. Clindamycin-Associated Colitis. Ann. Intern. Med. 1974, 81, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Lawson, P.A.; Citron, D.M.; Tyrrell, K.L.; Finegold, S.M. Reclassification of Clostridium difficile as Clostridioides difficile (Hall and O’Toole 1935) Preevot 1938. Anaerobe 2016, 40, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Lanzoni-Mangutchi, P.; Banerji, O.; Wilson, J.; Barwinska-Sendra, A.; Kirk, J.A.; Vaz, F.; O’Beirne, S.; Baslé, A.; El Omari, K.; Wagner, A.; et al. Structure and assembly of the S-layer in C. difficile. Nat. Commun. 2022, 13, 970. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, W.J.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. The structure of the S-layer of Clostridium difficile. J. Cell Commun. Signal. 2018, 12, 319–331. [Google Scholar] [CrossRef] [Green Version]
- CDC. Clostridioides difficile. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/clostridioides-difficile-508.pdf (accessed on 16 January 2023).
- Boyanova, L.; Markovska, R.; Hadzhiyski, P.; Yordanov, D.; Yaneva, P.; Mitov, I. Recurrent Clostridioides (Clostridium) difficile infection in a patient suffering from inflammatory bowel disease and benefits of resistotyping. Diagn. Microbiol. Infect. Dis. 2019, 94, 334–336. [Google Scholar] [CrossRef]
- Just, I.; Selzer, J.; von Eichel-Streiber, C.; Aktories, K. The low molecular mass GTP-binding protein Rh is affected by toxin a from Clostridium difficile. J. Clin. Investig. 1995, 95, 1026–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Buddle, J.E.; Fagan, R.P. Pathogenicity and virulence of Clostridioides difficile. Virulence 2023, 14, 2150452. [Google Scholar] [CrossRef]
- Jank, T.; Belyi, Y.; Aktories, K. Bacterial glycosyltransferase toxins. Cell. Microbiol. 2015, 17, 1752–1765. [Google Scholar] [CrossRef] [Green Version]
- Burke, K.E.; Lamont, J.T. Clostridium difficile Infection: A Worldwide Disease. Gut Liver 2014, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Prim. 2016, 2, 16020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drudy, D.; Fanning, S.; Kyne, L. Toxin A-negative, toxin B-positive Clostridium difficile. Int. J. Infect. Dis. 2007, 11, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Mani, N.; Dupuy, B. Regulation of toxin synthesis in Clostridium difficile by an alternative RNA polymerase sigma factor. Proc. Natl. Acad. Sci. USA 2001, 98, 5844–5849. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, A.; Govind, R. Regulation of Clostridioides difficile toxin production. Curr. Opin. Microbiol. 2021, 65, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Mengoli, M.; Barone, M.; Fabbrini, M.; D’Amico, F.; Brigidi, P.; Turroni, S. Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile. Genes 2022, 13, 2200. [Google Scholar] [CrossRef]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; Di Masi, A. Clostridium difficile Toxins A and B: Insights into Pathogenic Properties and Extraintestinal Effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.P.; Lyras, D.; Allen, D.L.; Mackin, K.E.; Howarth, P.M.; O’Connor, J.R.; Rood, J.I. Binary toxin production in Clostridium difficile is regulated by CdtR, a LytTR family response regulator. J. Bacteriol. 2007, 189, 7290–7301. [Google Scholar] [CrossRef] [Green Version]
- Anwar, F.; Roxas, B.A.P.; Shehab, K.W.; Ampel, N.M.; Viswanathan, V.K.; Vedantam, G. Low-toxin Clostridioides difficile RT027 strains exhibit robust virulence. Emerg. Microbes Infect. 2022, 11, 1982–1993. [Google Scholar] [CrossRef]
- Aitken, S.L.; Alam, M.J.; Khaleduzzuman, M.; Walk, S.T.; Musick, W.L.; Pham, V.P.; Christensen, J.L.; Atmar, R.L.; Garey, Y.X.A.K.W. In the Endemic Setting, Clostridium difficile Ribotype 027 Is Virulent But Not Hypervirulent—ERRATUM. Infect. Control Hosp. Epidemiol. 2015, 37, 124. [Google Scholar] [CrossRef] [Green Version]
- Fatima, R.; Aziz, M. The Hypervirulent Strain of Clostridium Difficile: NAP1/B1/027—A Brief Overview. Cureus 2019, 11, e3977. [Google Scholar] [CrossRef] [Green Version]
- Chilton, C.; Pickering, D.; Freeman, J. Microbiologic factors affecting Clostridium difficile recurrence. Clin. Microbiol. Infect. 2017, 24, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Taggart, M.G.; Snelling, W.J.; Naughton, P.J.; La Ragione, R.M.; Dooley, J.S.G.; Ternan, N.G. Biofilm regulation in Clostrid-ioides difficile: Novel systems linked to hypervirulence. PLoS Pathog. 2021, 17, e1009817. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1. [Google Scholar] [CrossRef]
- Bagdasarian, N.; Rao, K.; Malani, P.N. Diagnosis and treatment of Clostridium difficile in adults: A systematic review. JAMA 2015, 313, 398. [Google Scholar] [CrossRef] [PubMed]
- Vaishnavi, C. Clinical spectrum & pathogenesis of Clostridium difficile associated diseases. Indian J. Med. Res. 2010, 131, 487–499. [Google Scholar] [PubMed]
- Kelly, C.P. Can we identify patients at high risk of recurrent Clostridium difficile infection? Clin. Microbiol. Infect. 2012, 18 (Suppl. S6). [Google Scholar] [CrossRef] [Green Version]
- Song, J.H.; Kim, Y.S. Recurrent Clostridium difficile infection: Risk factors, treatment, and prevention. Gut Liver 2019, 13, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.N.; Karim, S.T.; Pascual, R.A.; Jowhar, L.M.; Anderson, S.E.; McBride, S.M. Chemical and Stress Resistances of Clostridium difficile Spores and Vegetative Cells. Front. Microbiol. 2016, 7, 1698. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, D.M.; Nerandzic, M.M.; Jury, L.A.; Jinno, S.; Chang, S.; Donskey, C.J. Acquisition of spores on gloved hands after contact with the skin of patients with Clostridium difficile infection and with environmental surfaces in their rooms. Am. J. Infect. Control 2012, 40, 556–558. [Google Scholar] [CrossRef]
- Furuya-Kanamori, L.; Marquess, J.; Yakob, L.; Riley, T.V.; Paterson, D.L.; Foster, N.F.; Huber, C.A.; Clements, A.C.A. Asymptomatic Clostridium difficile colonization: Epidemiology and clinical implications. BMC Infect. Dis. 2015, 15, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leffler, D.A.; Lamont, J.T. Clostridium difficile infection. N. Engl. J. Med. 2015, 373, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Liubakka, A.; Vaughn, B.P. Clostridium difficile Infection and Fecal Microbiota Transplant. AACN Adv. Crit. Care 2016, 27, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Khanafer, N.; Vanhems, P.; Bennia, S.; Martin-Gaujard, G.; Juillard, L.; Rimmelé, T.; Argaud, L.; Martin, O.; Huriaux, L.; Marcotte, G.; et al. Factors Associated with Clostridioides (Clostridium) difficile Infection and Colonization: Ongoing Prospec-tive Cohort Study in a French University Hospital. Int. J. Environ. Res. Public Health 2021, 18, 7528. [Google Scholar] [CrossRef]
- Hensgens, M.P.M.; Goorhuis, A.; Dekkers, O.M.; Kuijper, E.J. Time interval of increased risk for Clostridium difficile infection after exposure to antibiotics. J. Antimicrob. Chemother. 2011, 67, 742–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccioni, A.; Rosa, F.; Manca, F.; Pignataro, G.; Zanza, C.; Savioli, G.; Covino, M.; Ojetti, V.; Gasbarrini, A.; Franceschi, F.; et al. Gut Microbiota and Clostridium difficile: What We Know and the New Frontiers. Int. J. Mol. Sci. 2022, 23, 13323. [Google Scholar] [CrossRef]
- Pensinger, D.A.; Fisher, A.T.; Dobrila, H.A.; Van Treuren, W.; Gardner, J.O.; Higginbottom, S.K.; Carter, M.M.; Schumann, B.; Bertozzi, C.R.; Anikst, V.; et al. Butyrate Differentiates Permissiveness to Clostridioides difficile Infection and Influences Growth of Diverse C. difficile Isolates. Infect. Immun. 2023, 91, e00570-22. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Ross, R.P.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Chopra, T.; Hecht, G.; Tillotson, G. Gut microbiota and microbiota-based therapies for Clostridioides difficile infection. Front. Med. 2023, 9, 1093329. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.M.; Kelly, C.P.; Farraye, F. Clostridium difficile Infection in the Inflammatory Bowel Disease Patient. Inflamm. Bowel Dis. 2013, 19, 194–204. [Google Scholar] [CrossRef]
- Chitnis, A.S.; Holzbauer, S.M.; Belflower, R.M.; Winston, L.G.; Bamberg, W.M.; Lyons, C.; Farley, M.M.; Dumyati, G.K.; Wilson, L.E.; Beldavs, Z.G.; et al. Epidemiology of community-associated Clostridium difficile infection, 2009 through 2011. JAMA Intern. Med. 2013, 173, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePestel, D.D.; Aronoff, D.M. Epidemiology of Clostridium difficile Infection. J. Pharm. Pr. 2013, 26, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullish, B.H.; Williams, H.R. Clostridium difficile infection and antibiotic-associated diarrhoea. Clin. Med. 2018, 18, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Department of Health and Human Services. FDA Drug Safety Communication: Clostridium Difficile-Associated Diarrhea Can Be Associated with Stomach Acid Drugs Known as Proton Pump Inhibitors (Ppis). Available online: http://www.fda.gov/drugs/drugsafety/ucm290510.htm (accessed on 26 February 2023).
- Imhann, F.; Bonder, M.J.; Vila, A.V.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.M.; et al. Proton pump inhibitors affect the gut microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finney, J.M.T. Gastro-enterostomy for cicatrizing ulcer of the pylorus. Bull. Johns Hopkins Hosp. 1983, 4, 53. [Google Scholar]
- Redelings, M.D.; Sorvillo, F.; Mascola, L. Increase in Clostridium difficile-related mortality rates, United States, 1999–2004. Emerg. Infect. Dis. 2007, 13, 1417–1419. [Google Scholar] [CrossRef] [Green Version]
- Kazakova, S.V.; Ware, K.; Baughman, B.; Bilukha, O.; Paradis, A.; Sears, S.; Thompson, A.; Jensen, B.; Wiggs, L.; Bessette, J.; et al. A Hospital Outbreak of Diarrhea Due to an Emerging Epidemic Strain of Clostridium difficile. Arch. Intern. Med. 2006, 166, 2518–2524. [Google Scholar] [CrossRef] [Green Version]
- Pépin, J.L.; Valiquette, M.E.; Clossette, B. Mortality attributed to nosocomial Clostridium difficile-associated disease during an epidemic caused by a hyperviluent strain in Quebec. CMAJ 2005, 173, 1037. [Google Scholar] [CrossRef] [Green Version]
- Labbé, A.C.; Poirier, L.; Maccannell, D.; Louie, T.; Savoie, M.; Béliveau, C.; Laverdière, M.; Pépin, J. Clostridium difficile in-fections in a Canadian tertiary care hospital before and during a regional epidemic associated with the BI/NAP1/027 strain. Antimicrob. Agents Chemother. 2008, 52, 3180–3187. [Google Scholar] [CrossRef] [Green Version]
- Kuijper, E.J.; Coignard, B.; Brazier, J.S.; Suetens, C.; Drudy, D.; Wiuff, C.; Pituch, H.; Reichert, P.; Schneider, F.; Widmer, A.F.; et al. Update of Clostridium difficile-associated disease due to PCR ribotype 027 in Europe. Eurosurveillance 2007, 12, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Luo, Y.; Grinspan, A.M. Epidemiology of community-acquired and recurrent Clostridioides difficile infection. Ther. Adv. Gastroenterol. 2021, 14, 17562848211016248. [Google Scholar] [CrossRef]
- Guh, A.Y.; Mu, Y.; Winston, L.G.; Johnston, H.; Olson, D.; Farley, M.M.; Wilson, L.E.; Holzbauer, S.M.; Phipps, E.C.; Dumyati, G.K.; et al. Emerging Infections Program Clostridioides difficile Infection Working Group. Trends in U.S. Burden of Clostridioides difficile Infection and Outcomes. N. Engl. J. Med. 2020, 382, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- van Dorp, S.M.; Kinross, P.; Gastmeier, P.; Behnke, M.; Kola, A.; Delmée, M.; Pavelkovich, A.; Mentula, S.; Barbut, F.; Hajdu, A.; et al. For the European Clostridium difficile Infection Surveillance Network (ECDIS-Net) on behalf of all participants. Standardised surveillance of Clostridium difficile infection in European acute care hospitals: A pilot study, 2013. Eurosurveillance 2016, 21, 30293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control. Clostridioides (Clostridium) difficile Infections; Annual epidemiological report for 2016–2017; ECDC: Stockholm, Sweden, 2022. [Google Scholar]
- Kracík, M.; Dolinová, I.; Žemličková, H. Ribotyping of Clostridioides difficile in the Liberec Regional Hospital: A tertiary health care facility. Folia Microbiol. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aptekorz, M.; Szczegielniak, A.; Wiechuła, B.; Harmanus, C.; Kuijper, E.; Martirosian, G. Occurrence of Clostridium difficile ribotype 027 in hospitals of Silesia, Poland. Anaerobe 2017, 45, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Stojanovic, P.; Harmanus, C.; Kuijper, E.J. Community-onset Clostridioides difficile infection in south Serbia. Anaerobe 2023, 79, 102669. [Google Scholar] [CrossRef]
- Novakova, E.; Stefkovicova, M.; Kopilec, M.G.; Novak, M.; Kotlebova, N.; Kuijper, E.; Krutova, M. The emergence of Clos-tridium difficile ribotypes 027 and 176 with a predominance of the Clostridium difficile ribotype 001 recognized in Slovakia following the European standardized Clostridium difficile infection surveillance of 2016. Int. J. Infect. Dis. 2020, 90, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.; Vernon, J.; Pilling, S.; Morris, K.; Nicolson, S.; Shearman, S.; Clark, E.; Palacios-Fabrega, J.A.; Wilcox, M. Pan-European Longitudinal Surveillance of Antibiotic Resistance among Prevalent Clostridium difficile Ribotypes’ Study Group. Five-year Pan-European, longitudinal surveillance of Clostridium difficile ribotype prevalence and antimicrobial re-sistance: The extended ClosER study. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 169–177. [Google Scholar]
- Persson, S.; Nielsen, H.L.; Coia, J.E.; Engberg, J.; Olesen, B.S.; Engsbro, A.L.; Petersen, A.M.; Holt, H.M.; Lemming, L.; Marmolin, E.S.; et al. Sentinel surveillance and epidemiology of Clostridioides difficile in Denmark, 2016 to 2019. Eurosurveillance 2022, 27, 2200244. [Google Scholar] [CrossRef]
- Dirks, E.E.; Luković, J.A.; Peltroche-Llacsahuanga, H.; Herrmann, A.; Mellmann, A.; Arvand, M. Molecular Epidemiology, Clinical Course, and Implementation of Specific Hygiene Measures in Hospitalised Patients with Clostridioides difficile Infec-tion in Brandenburg, Germany. Microorganisms 2022, 11, 44. [Google Scholar] [CrossRef]
- Kachrimanidou, M.; Metallidis, S.; Tsachouridou, O.; Harmanus, C.; Lola, V.; Protonotariou, E.; Skoura, L.; Kuijper, E. Predominance of Clostridioides difficile PCR ribotype 181 in northern Greece, 2016–2019. Anaerobe 2022, 76, 102601. [Google Scholar] [CrossRef] [PubMed]
- Dobreva, E.G.; Ivanov, I.N.; Vathcheva-Dobrevska, R.S.; Ivanova, K.I.; Asseva, G.D.; Petrov, P.K.; Kantardjiev, T.V. Ad-vances in molecular surveillance of Clostridium difficile in Bulgaria. J. Med. Microbiol. 2013, 62 Pt 9, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Brajerova, M.; Zikova, J.; Krutova, M. Clostridioides difficile epidemiology in the Middle and the Far East. Anaerobe 2022, 74, 102542. [Google Scholar] [CrossRef] [PubMed]
- Senoh, M.; Kato, H. Molecular epidemiology of endemic Clostridioides difficile infection in Japan. Anaerobe 2022, 74, 102510. [Google Scholar] [CrossRef] [PubMed]
- Kullin, B.; Abratt, V.R.; Reid, S.J.; Riley, T.V. Clostridioides difficile infection in Africa: A narrative review. Anaerobe 2022, 74, 102549. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.-Y.; Bai, L.-L.; Zhang, W.-Z.; Li, G.-W.; Lu, J.-X. A narrative review of Clostridioides difficile infection in China. Anaerobe 2022, 74, 102540. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Amador, L.; Quesada-Gómez, C.; Rodríguez, C. Clostridioides difficile in Latin America: A comprehensive review of literature (1984–2021). Anaerobe 2022, 74, 102547. [Google Scholar] [CrossRef]
- Hall, A.J.; Curns, A.T.; McDonald, L.C.; Parashar, U.D.; Lopman, B.A. The Roles of Clostridium difficile and Norovirus Among Gastroenteritis-Associated Deaths in the United States, 1999–2007. Clin. Infect. Dis. 2012, 55, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile Infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, B.G.; Vinithakumari, A.A.; Sponseller, B.; Tangudu, C.; Mooyottu, S. Prevalence, Colonization, Epidemiology, and Public Health Significance of Clostridioides difficile in Companion Animals. Front. Veter. Sci. 2020, 7, 512551. [Google Scholar] [CrossRef]
- Walczak, R.; Paek, M.; Suran, J.; Amory, J.T.; Specchi, S.; Sanchez, M. Radiography and ultrasonography of pneumatosis intestinalis in a cat. Veter. Radiol. Ultrasound 2018, 61, E26–E30. [Google Scholar] [CrossRef] [PubMed]
- Perrin, J.; Buogo, C.; Gallusser, A.; Burnens, A.P.; Nicolet, J. Intestinal Carriage of Clostridium difficile in Neonate Dogs. J. Veter. Med. Ser. B 1993, 40, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.E.; Nunnally, A.E.; Jimenez, V., Jr.; Cope, E.K.; Sahl, J.W.; Sheridan, K.; Hornstra, H.M.; Vinocur, J.; Settles, E.W.; Headley, K.C.; et al. Domestic canines do not display evidence of gut microbial dysbiosis in the presence of Clostridioides (Clostridium) difficile, despite cellular susceptibility to its toxins. Anaerobe 2019, 58, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.S.; Markel, M.E.; Garcia-Mazcorro, J.F.; Unterer, S.; Heilmann, R.M.; Dowd, S.E.; Kachroo, P.; Ivanov, I.; Mi-namoto, Y.; Dillman, E.M.; et al. The fecal microbiome in dogs with acute diarrhea and idiopathic inflammatory bowel dis-ease. PLoS ONE 2012, 7, e51907. [Google Scholar] [CrossRef] [Green Version]
- lvarez-Pérez, S.; Blanco, J.L.; Peláez, T.; Lanzarot, M.P.; Harmanus, C.; Kuijper, E.; García, M.E. Faecal shedding of anti-microbial-resistant Clostridium difficile strains by dogs. J. Small Anim. Pract. 2015, 56, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Alves, F.; Castro, R.; Pinto, M.; Nunes, A.; Pomba, C.; Oliveira, M.; Silveira, L.; Gomes, J.P.; Oleastro, M. Molecular epi-demiology of Clostridioides difficile in companion animals: Genetic overlap with human strains and public health concerns. Front. Public Health 2023, 10, 1070258. [Google Scholar] [CrossRef]
- Stoesser, N.; Eyre, D.W.; Quan, T.P.; Godwin, H.; Pill, G.; Mbuvi, E.; Vaughan, A.; Griffiths, D.; Martin, J.; Fawley, W.; et al. Modernising Medical Microbiology Informatics Group (MMMIG). Epidemiology of Clostridium difficile in infants in Oxford-shire, UK: Risk factors for colonization and carriage, and genetic overlap with regional C. difficile infection strains. PLoS ONE 2017, 12, e0182307. [Google Scholar] [CrossRef] [Green Version]
- Loo, V.G.; Brassard, P.; Miller, M.A. Household Transmission of Clostridium difficile to Family Members and Domestic Pets. Infect. Control Hosp. Epidemiol. 2016, 37, 1342–1348. [Google Scholar] [CrossRef]
- Lefebvre, S.L.; Arroyo, L.G.; Weese, J.S. Epidemic Clostridium difficile Strain in Hospital Visitation Dog. Emerg. Infect. Dis. 2006, 12, 1036–1037. [Google Scholar] [CrossRef]
- Rodriguez, C.; Taminiau, B.; Bouchafa, L.; Romijn, S.; Van Broeck, J.; Delmée, M.; Clercx, C.; Daube, G. Clostridium difficile beyond stools: Dog nasal discharge as a possible new vector of bacterial transmission. Heliyon 2019, 5, e01629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orden, C.; Neila, C.; Blanco, J.L.; Álvarez-Pérez, S.; Harmanus, C.; Kuijper, E.J.; García, M.E. Recreational sandboxes for children and dogs can be a source of epidemic ribotypes of Clostridium difficile. Zoonoses Public Health 2017, 65, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janezic, S.; Mlakar, S.; Rupnik, M. Dissemination of Clostridium difficile spores between environment and households: Dog paws and shoes. Zoonoses Public Health 2018, 65, 669–674. [Google Scholar] [CrossRef]
- Diniz, A.N.; Moura, L.N.F.; Cruz, D.S.G.; Oliveira Junior, C.A.; Figueiredo, H.C.P.; Cunha, J.L.R.; Vilela, E.G.; Kuijper, E.J.; Wilcox, M.H.; Lobato, F.C.F.; et al. Characterization of the virulence of three novel clade 2 Clostridioides (Clostridium) difficile strains and a two-year screening in animals and humans in Brazil. PLoS ONE 2022, 17, e0273013. [Google Scholar] [CrossRef]
- Rabold, D.; Espelage, W.; Abu Sin, M.; Eckmanns, T.; Schneeberg, A.; Neubauer, H.; Möbius, N.; Hille, K.; Wieler, L.H.; Seyboldt, C.; et al. The zoonotic potential of Clostridium difficile from small companion animals and their owners. PLoS ONE 2018, 13, e0193411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orden, C.; Blanco, J.L.; Álvarez-Pérez, S.; García-Sancho, M.E.; Blanco, J.L.; Garcia-Sancho, M.; Rodriguez-Franco, F.; Sainz, A.; Villaescusa, A.; Garcia, M.E.; et al. Isolation of Clostridium difficile from dogs with digestive disorders, including stable metronidazole-resistant strains. Anaerobe 2017, 43, 78–81. [Google Scholar] [CrossRef]
- Spigaglia, P.; Drigo, I.; Barbanti, F.; Mastrantonio, P.; Bano, L.; Bacchin, C.; Puiatti, C.; Tonon, E.; Berto, G.; Agnoletti, F. Antibiotic resistance patterns and PCR-ribotyping of Clostridium difficile strains isolated from swine and dogs in Italy. Anaerobe 2015, 31, 42–46. [Google Scholar] [CrossRef]
- Knetsch, C.W.; Connor, T.R.; Mutreja, A.; van Dorp, S.M.; Sanders, I.M.; Browne, H.P.; Harris, D.; Lipman, L.; Keessen, E.C.; Corver, J.; et al. Whole genome sequencing reveals potential spread of Clostridium difficile between humans and farm animals in the Netherlands, 2002 to 2011. Eurosurveillance 2014, 19, 20954. [Google Scholar] [CrossRef] [Green Version]
- Debast, S.B.; van Leengoed, L.A.M.G.; Goorhuis, A.; Harmanus, C.; Kuijper, E.J.; Bergwerff, A.A. Clostridium difficile PCR ribotype 078 toxinotype V found in diarrhoeal pigs identical to isolates from affected humans. Environ. Microbiol. 2009, 11, 505–511. [Google Scholar] [CrossRef]
- Moloney, G.; Eyre, D.W.; Mac Aogáin, M.; McElroy, M.C.; Vaughan, A.; Peto, T.E.A.; Crook, D.W.; Rogers, T.R. Human and Porcine Transmission of Clostridioides difficile Ribotype 078, Europe. Emerg. Infect. Dis. 2021, 27, 2294–2300. [Google Scholar] [CrossRef]
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.W.; Fung, R.; Golubchik, T.; Harding, R.M.; Jeffery, K.J.M.; Jolley, K.A.; et al. Multilocus Sequence Typing of Clostridium difficile. J. Clin. Microbiol. 2010, 48, 770–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.T.; Mulvey, M.R.; Zhanel, G.G.; Bay, D.C.; Reid-Smith, R.J.; Janecko, N.; Golding, G.R. A Clostridioides difficile sur-veillance study of Canadian retail meat samples from 2016–2018. Anaerobe 2022, 74, 102551. [Google Scholar] [CrossRef] [PubMed]
- Kachrimanidou, M.; Tzika, E.; Filioussis, G. Clostridioides (Clostridium) difficile in Food-Producing Animals, Horses and Household Pets: A Comprehensive Review. Microorganisms 2019, 7, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kecerova, Z.; Cizek, A.; Nyc, O.; Krutova, M. Clostridium difficile isolates derived from Czech horses are resistant to en-rofloxacin; cluster to clades 1 and 5 and ribotype 033 predominates. Anaerobe 2019, 56, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.R.; Seyboldt, C.; Rupnik, M. Non-human C. difficile Reservoirs and Sources: Animals, Food, Environment. Updates Clostridium Difficile Eur. Adv. Microbiol. Infect. Dis. Public Health 2018, 8, 227–243. [Google Scholar] [CrossRef]
- Keessen, E.C.; Gaastra, W.; Lipman, L.J. Clostridium difficile infection in humans and animals, differences and similarities. Vet. Microbiol. 2011, 153, 205–217. [Google Scholar] [CrossRef]
- Tkalec, V.; Viprey, V.; Davis, G.; Janezic, S.; Sente, B.; Devos, N.; Wilcox, M.; Davies, K.; Rupnik, M.; on behalf of the COMBACTE-CDI consortium. Clostridioides difficile positivity rate and PCR ribotype distribution on retail potatoes in 12 European countries, January to June 2018. Eurosurveillance 2022, 27, 2100417. [Google Scholar] [CrossRef]
- Tkalec, V.; Janezic, S.; Skok, B.; Simonic, T.; Mesaric, S.; Vrabic, T.; Rupnik, M. High Clostridium difficile contamination rates of domestic and imported potatoes compared to some other vegetables in Slovenia. Food Microbiol. 2018, 78, 194–200. [Google Scholar] [CrossRef]
- Lim, S.C.; Foster, N.F.; Elliott, B.; Riley, T.V. High prevalence of Clostridium difficile on retail root vegetables, Western Aus-tralia. J. Appl. Microbiol. 2018, 124, 585–590. [Google Scholar] [CrossRef]
- Eckert, C.; Burghoffer, B.; Barbut, F. Contamination of ready-to-eat raw vegetables with Clostridium difficile in France. J. Med. Microbiol. 2013, 62, 1435–1438. [Google Scholar] [CrossRef]
- Janezic, S.; Potocnik, M.; Zidaric, V.; Rupnik, M. Highly Divergent Clostridium difficile Strains Isolated from the Environment. PLoS ONE 2016, 11, e0167101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.J.; Walk, S.T.; Endres, B.T.; Basseres, E.; Khaleduzzaman, M.; Amadio, J.; Musick, W.L.; Christensen, J.L.; Kuo, J.; Atmar, R.L.; et al. Community Environmental Contamination of Toxigenic Clostridium difficile. Open Forum Infect. Dis. 2017, 4, ofx018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, P.; Whyte, P.; Burgess, C.; Ekhlas, D.; Bolton, D. Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields. Pathogens 2022, 11, 1310. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gonzales-Luna, A.J.; Lancaster, C.K.; McPherson, J.K.; Begum, K.; Jahangir, A.M.; Garey, K.W. Multi-country sur-veillance of Clostridioides difficile demonstrates high prevalence of spores in non-healthcare environmental settings. Anaerobe 2022, 75, 102543. [Google Scholar] [CrossRef] [PubMed]
- Klein, H. 2021 Shoe Soles Are a Major Cause of C. difficile Transmission. AJMC. Available online: https://www.ajmc.com/view/shoe-soles-are-a-major-cause-of-c-difficile-transmission (accessed on 24 January 2023).
- Petrosillo, N. Clostridioides difficile Infection: A Never-Ending Challenge. J. Clin. Med. 2022, 11, 4115. [Google Scholar] [CrossRef] [PubMed]
- FDA. FDA Approves First Fecal Microbiota Product. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-fecal-microbiota-product (accessed on 19 March 2023).
- BiomeBank. BiomeBank Announces World First Regulatory Approval for Donor Derived Microbiome Drug. Available online: https://www.biomebank.com/news/biomebank-announces-world-first-regulatory-approval-for-donor-derived-microbiome-drug/ (accessed on 19 March 2023).
- Giesemann, T.; Guttenberg, G.; Aktories, K. Human alpha-defensins inhibit Clostridium difficile toxin B. Gastroenterology 2008, 134, 2049–2058. [Google Scholar] [CrossRef] [PubMed]
- Heber, S.; Barthold, L.; Baier, J.; Papatheodorou, P.; Fois, G.; Frick, M.; Barth, H.; Fischer, S. Inhibition of Clostridioides difficile Toxins TcdA and TcdB by Ambroxol. Front. Pharmacol. 2022, 12, 809595. [Google Scholar] [CrossRef]
- Qian, X.; Yanagi, K.; Kane, A.V.; Alden, N.; Lei, M.; Snydman, D.R.; Vickers, R.J.; Lee, K.; Thorpe, C.M. Ridinilazole, a narrow spectrum antibiotic for treatment of Clostridioides difficile infection, enhances preservation of microbiota-dependent bile acids. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 319, G227–G237. [Google Scholar] [CrossRef]
- Oksi, J.; Anttila, V.J.; Mattila, E. Treatment of Clostridioides (Clostridium) difficile infection. Ann Med. 2020, 52, 12–20. [Google Scholar]
- Alshrari, A.S.; Hudu, S.A.; Elmigdadi, F.; Imran, M. The Urgent Threat of Clostridioides difficile Infection: A Glimpse of the Drugs of the Future, with Related Patents and Prospects. Biomedicines 2023, 11, 426. [Google Scholar] [CrossRef]
- Garey, K.W.; McPherson, J.; Dinh, A.Q.; Hu, C.; Jo, J.; Wang, W.; Lancaster, C.K.; Gonzales-Luna, A.J.; Loveall, C.; Begum, K.; et al. Efficacy, safety, pharmacokinetics, and microbiome changes of ibezapolstat in adults with Clostridioides difficile In-fection: A phase 2a multicenter clinical trial. Clin. Infect. Dis. 2022, 75, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Hyte, M.L.; Arphai, L.J.; Vaughn, C.J.; Durham, S.H. The Role of Bezlotoxumab for the Prevention of Recurrent Clostridioides difficile Infections: A Review of the Current Literature and Paradigm Shift after 2021. Antibiotics 2022, 11, 1211. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.M.; Hegarty, J.W.; Healy, B.; Schulz, S.; Walsh, C.J.; Hill, C.; Ross, R.P.; Rea, M.C.; Farquhar, R.; Chesnel, L. Identification of ADS024, a newly characterized strain of Bacillus velezensis with direct Clostridiodes difficile killing and toxin degradation bio-activities. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- van Prehn, J.; Reigadas, E.; Vogelzang, E.H.; Bouza, E.; Hristea, A.; Guery, B.; Krutova, M.; Norén, T.; Allerberger, F.; Coia, J.E.; et al. Guideline Committee of the European Study Group on Clostridioides difficile. European Society of Clinical Mi-crobiology and Infectious Diseases: 2021 update on the treatment guidance document for Clostridioides difficile infection in adults. Clin. Microbiol. Infect. 2021, 27 (Suppl. S2), S1–S21. [Google Scholar] [CrossRef] [PubMed]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Soucy, J.R.; Westwood, D.; Daneman, N.; MacFadden, D.R. Antibiotic prescribing in patients with COVID-19: Rapid review and meta-analysis. Clin. Microbiol. Infect. 2021, 27, 520–531. [Google Scholar] [CrossRef]
- Fukushige, M.; Ngo, N.H.; Lukmanto, D.; Fukuda, S.; Ohneda, O. Effect of the COVID-19 pandemic on antibiotic con-sumption: A systematic review comparing 2019 and 2020 data. Front. Public Health 2022, 10, 946077. [Google Scholar] [CrossRef]
- Hamilton, A.; Poleon, S.; Cherian, J.; Cosgrove, S.; Laxminarayan, R.; Klein, E. COVID-19 and Outpatient Antibiotic Prescriptions in the United States: A County-level Analysis. Open Forum Infect. Dis. 2023, 10, ofad096. [Google Scholar] [CrossRef]
- CDC. COVID-19 Impact on HAIs. Available online: https://www.cdc.gov/hai/data/portal/covid-impact-hai.html (accessed on 19 March 2023).
- Kovačević, N.; Petrić, V.; Pete, M.; Popović, M.; Plećaš-Đurić, A.; Pejaković, S.; Tomić, S.; Damjanov, D.; Kosijer, D.; Lekin, M. Clostridioides Difficile Infection before and during Coronavirus Disease 2019 Pandemic—Similarities and Differences. Microorganisms 2022, 10, 2284. [Google Scholar] [CrossRef]
- Choi, K.B.; Du, T.; Silva, A.; Golding, G.R.; Pelude, L.; Mitchell, R.; Rudnick, W.; Hizon, R.; Al-Rawahi, G.N.; Chow, B.; et al. Trends in Clostridioides difficile infection rates in Canadian hospitals during the coronavirus disease 2019 (COVID-19) pandemic. Infect. Control Hosp. Epidemiol. 2022, 1–4. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, D.; Sun, X. Development of an Effective Nontoxigenic Clostridioides difficile–Based Oral Vaccine against C. difficile Infection. Microbiol. Spectr. 2022, 10, e00263-22. [Google Scholar] [CrossRef]
- Chandra, H.; Kovall, R.A.; Yadav, J.S.; Sun, X. Host Immune Responses to Surface S-Layer Proteins (SLPs) of Clostridioides difficile. Microorganisms 2023, 11, 380. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Drug | Mechanisms of Action | Advantages | Disadvantage |
|---|---|---|---|
| Standard Antibiotic Therapy | |||
| Vancomycin | Blocks peptidoglycan synthesis | Superior than metronidazole, can be applied orally and is not absorbed through gut mucosa | Pressure on the intestinal microbiota and risks for recurrence |
| Fidaxomicin | inhibits bacterial RNA polymerase | Can be used as first-line treatment for primary CDI, reduces recurrence—especially if used in an extended-pulsed regimen | Not superior to vancomycin for RT027 C. difficile strains |
| Metronidazole | Damages DNA in reduced form | Can be used in mild CDIs, low cost | Possibility for recurrence, side effects, such as allergic reactions, gastrointestinal irritation and neuropathy and interactions with ethanol and some common drugs (e.g., warfarin); increase of vancomycin-resistant enterococci |
| Antibiotic Therapy—In Investigation | |||
| Ridinilazole | Inhibition of cell division (bactericidal) and toxin production | Narrow-spectrum; non-absorbable; high inhibitory activity against C. difficile and minimal activity against both Gram-positive and Gram-negative aerobic and anaerobic intestinal microorganisms. Decreases recurrent infections In phase II/III trials, FDA fast-tracking status | More studies are required |
| Ibezapolstat | DNA polymerase IIIC inhibitor | Activity against C. difficile and preservation of Actinobacteria and bile acid equilibrium. | Clinical benefits of the agent should be more widely investigated |
| Toxin-blocking treatment (antibody) | |||
| Bezlotoxumab | Fully humanised monoclonal antibody against C. difficile—blocks TcdB toxin | Good adjunctive to other therapy, decrease recurrent CDIs | Risks for patients with history of heart failure. Parenteral administration, cost |
| Microbiome-modulating therapy | |||
| Faecal microbiota transplantation (FMT) | Reverts normal balance in gut microbiota | 75–90% success, decrease recurrence | FMT is not available everywhere, and all patients are not eligible for the procedure. Long-term safety- the main risk is transfer of infectious pathogens from the donor to the recipient, difficulties in introducing donor faeces, need for repeat transplantation, autoimmunological disorders |
| Purified Firmicutes spores ((SER-109) | Modulates gut microbiota | Reduce the risk of CDI | There are no important side effects, but the investigation is in progress |
| Live biotherapeutic (a strain of Bacillus velezensis) (ADS024) | Modulates gut microbiota; produces proteases that break down the CD toxins | Potential candidate for preventing the recurrence of CDI | Additional in vitro and clinical studies are needed |
| Quality-controlled stool product (RBX2660) | Modulates gut microbiota | Potential candidate for preventing the recurrence of CDI | |
| Probiotics- Saccharomyces boulardii, Lactobacillus spp. | Modulate gut microbiota | Prophylaxix of C. difficile infections | Controversial results, not included in the guidelines |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markovska, R.; Dimitrov, G.; Gergova, R.; Boyanova, L. Clostridioides difficile, a New “Superbug”. Microorganisms 2023, 11, 845. https://doi.org/10.3390/microorganisms11040845
Markovska R, Dimitrov G, Gergova R, Boyanova L. Clostridioides difficile, a New “Superbug”. Microorganisms. 2023; 11(4):845. https://doi.org/10.3390/microorganisms11040845
Chicago/Turabian StyleMarkovska, Rumyana, Georgi Dimitrov, Raina Gergova, and Lyudmila Boyanova. 2023. "Clostridioides difficile, a New “Superbug”" Microorganisms 11, no. 4: 845. https://doi.org/10.3390/microorganisms11040845