

Beneficial Effects of Bacillus amyloliquefaciens D1 Soy Milk Supplementation on Serum Biochemical Indexes and Intestinal Health of Bearded Chickens
 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
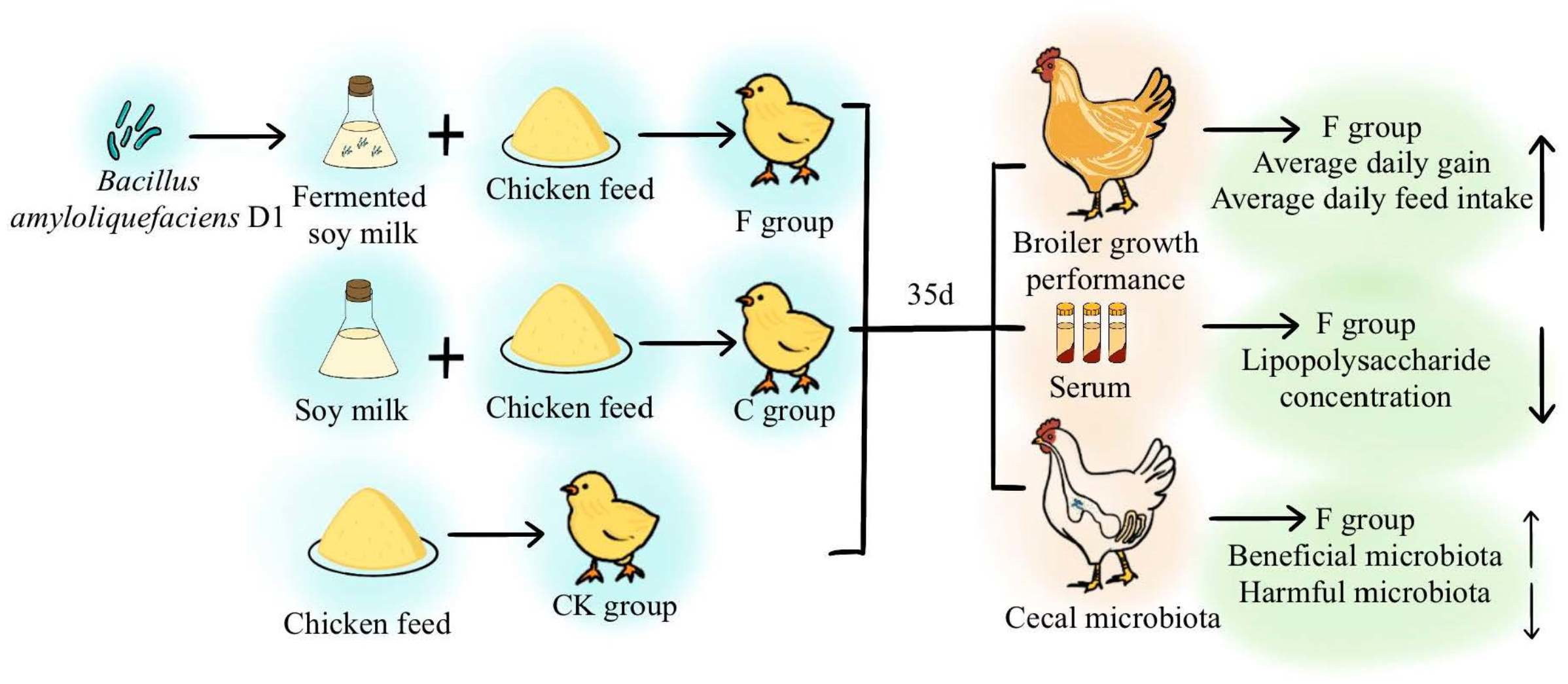
2.1. Bearded Chickens and Experimental Design
2.2. Preparation of Soy Milk
2.3. Calculation of Production Performance Indexes
2.4. Sample Collection and Treatment
2.5. Statistical Analysis
3. Results
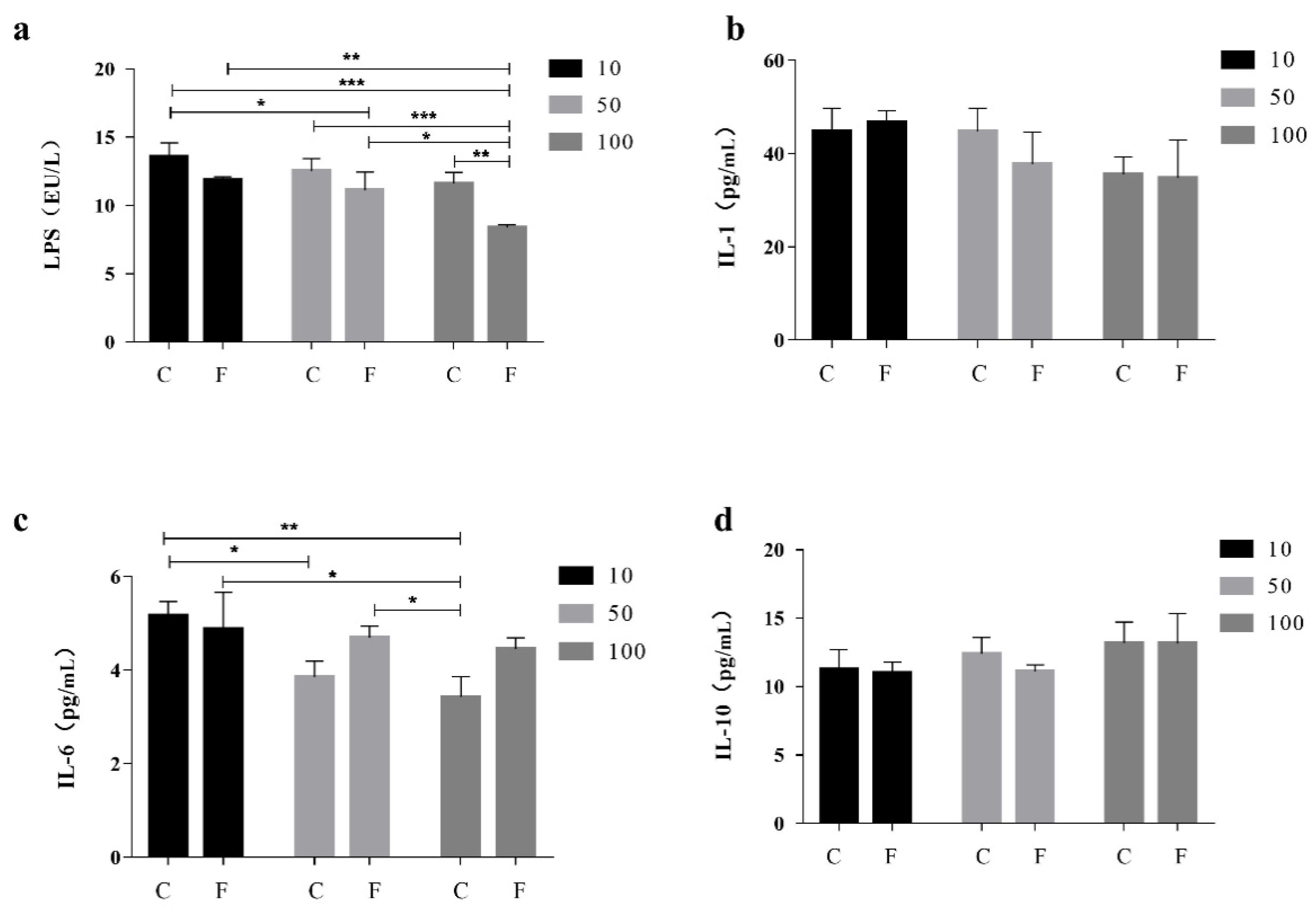
3.1. Effect of B. amyloliquefaciens D1 Fermented Soy Milk on the Productive Growth Performance and Serum Biochemical Indexes of Bearded Chickens
3.2. Effect of alpha Diversity Index of Fermented Soy Milk Broiler Cecum Contents by B. amyloliquefaciens D1
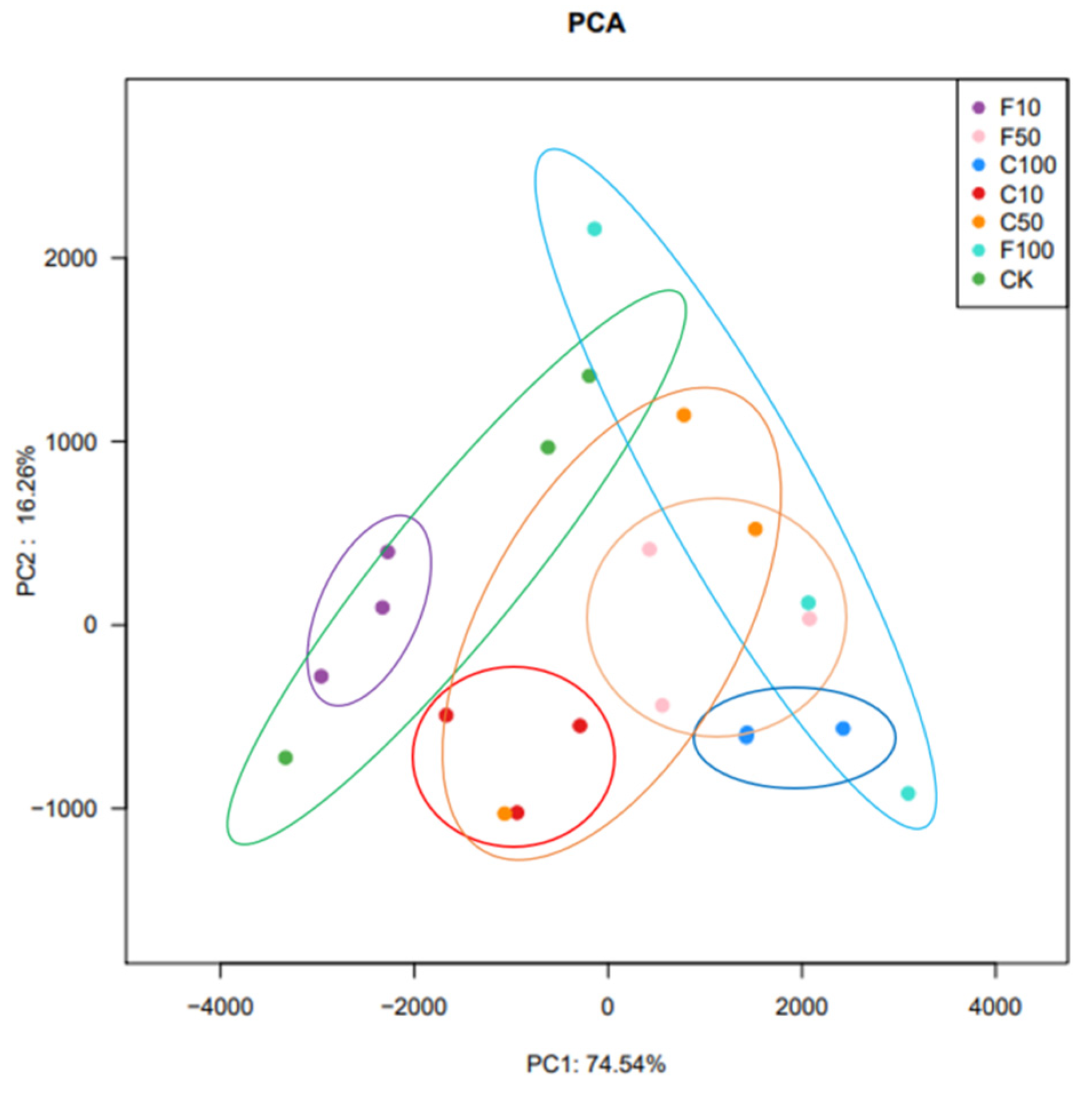
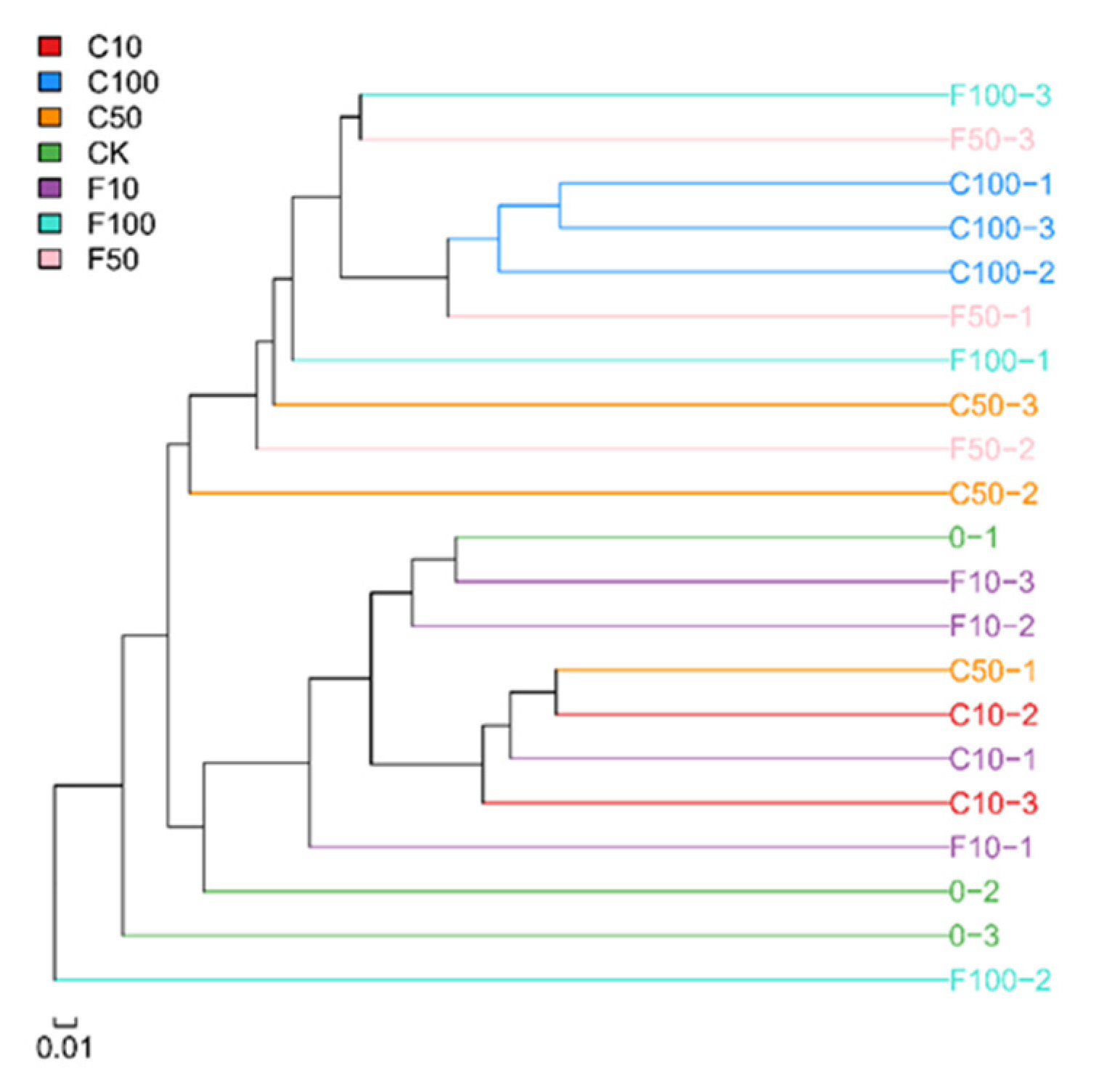
3.3. Effect of B. amyloliquefaciens D1 Fermentation of Soy Milk Broiler Cecum Contents on β-Diversity of Bacterial Microbiota
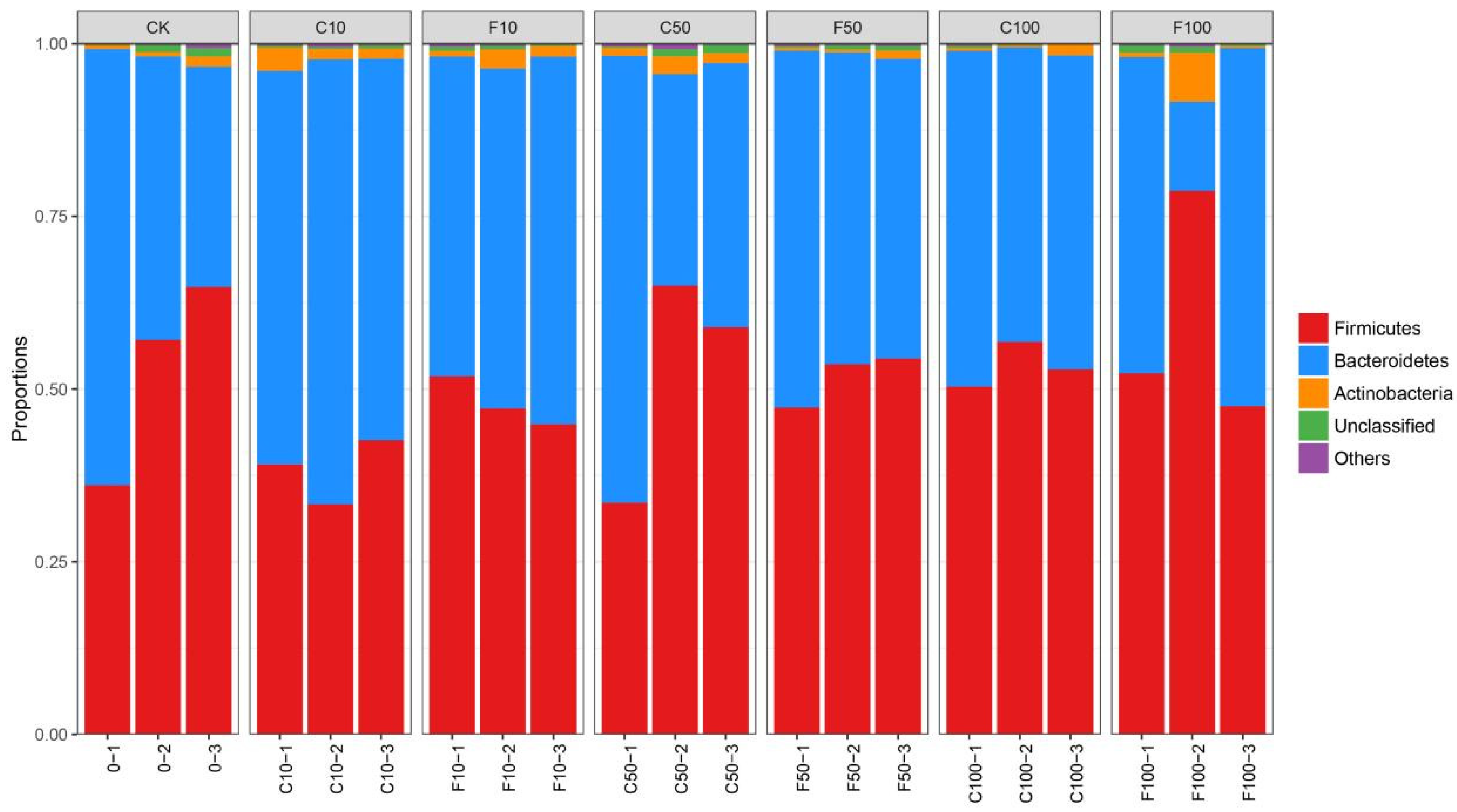
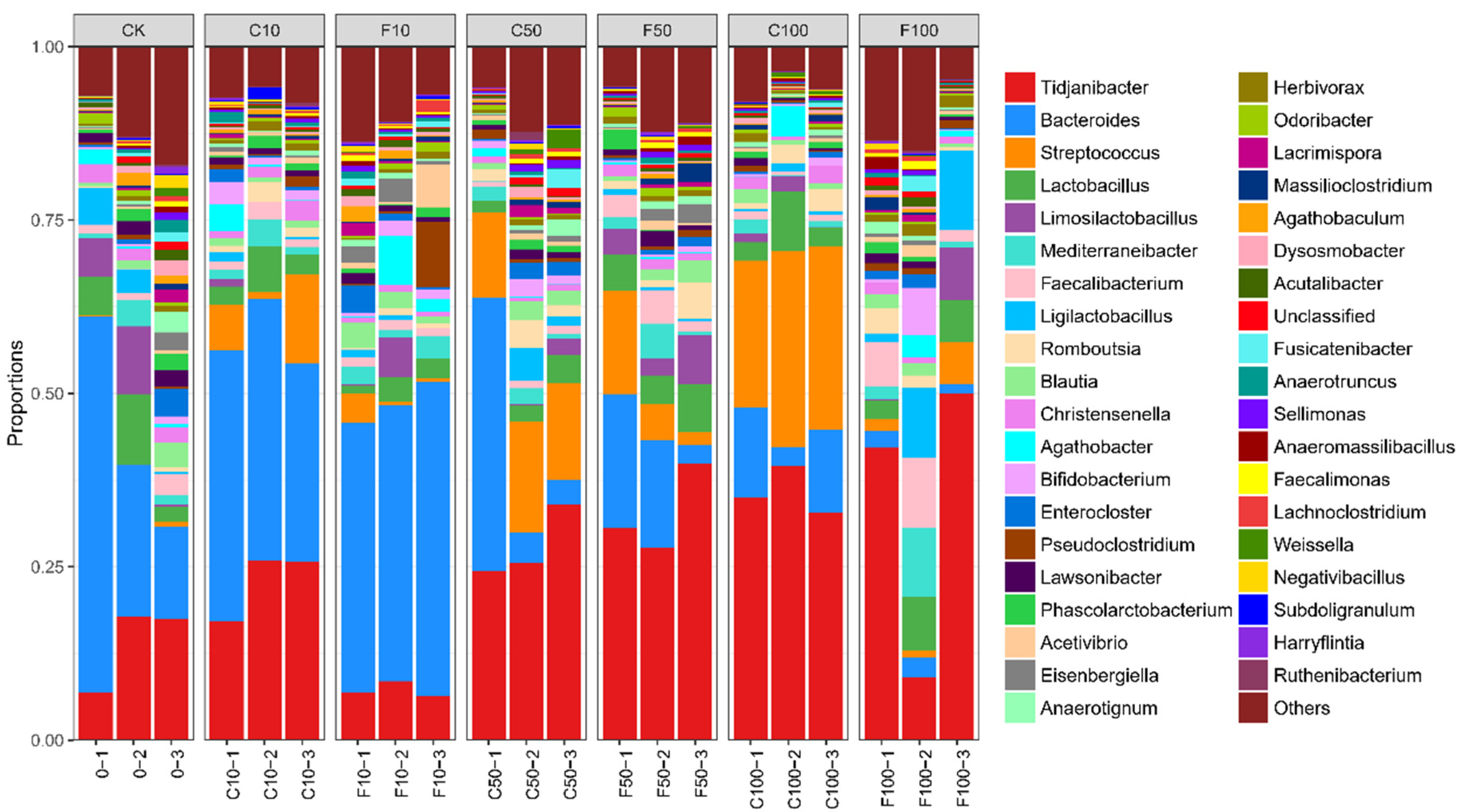
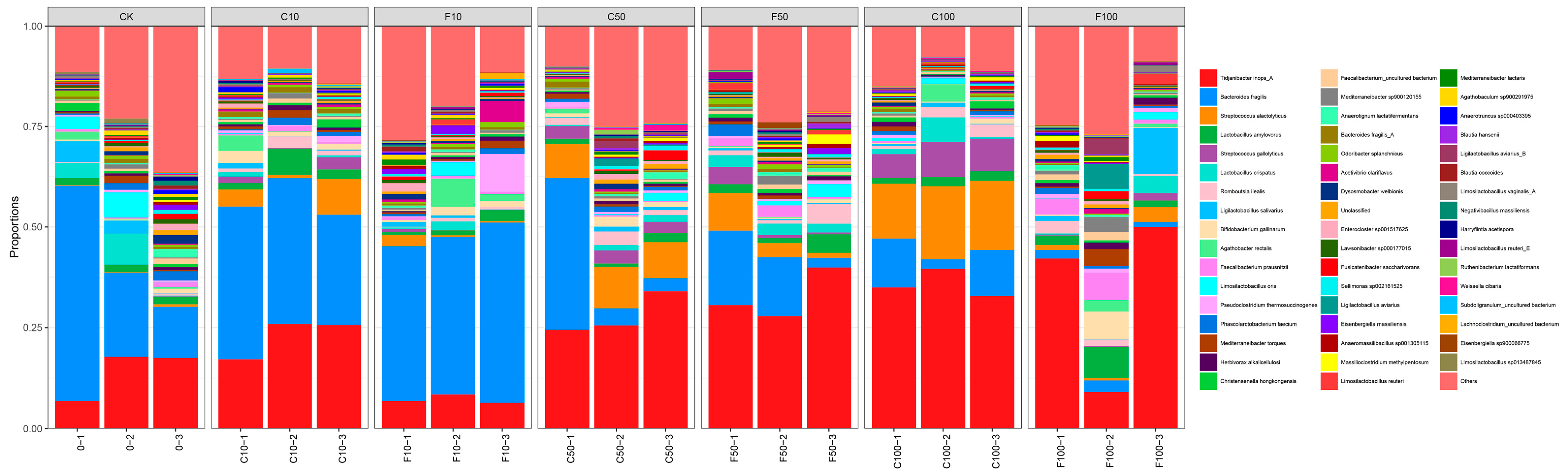
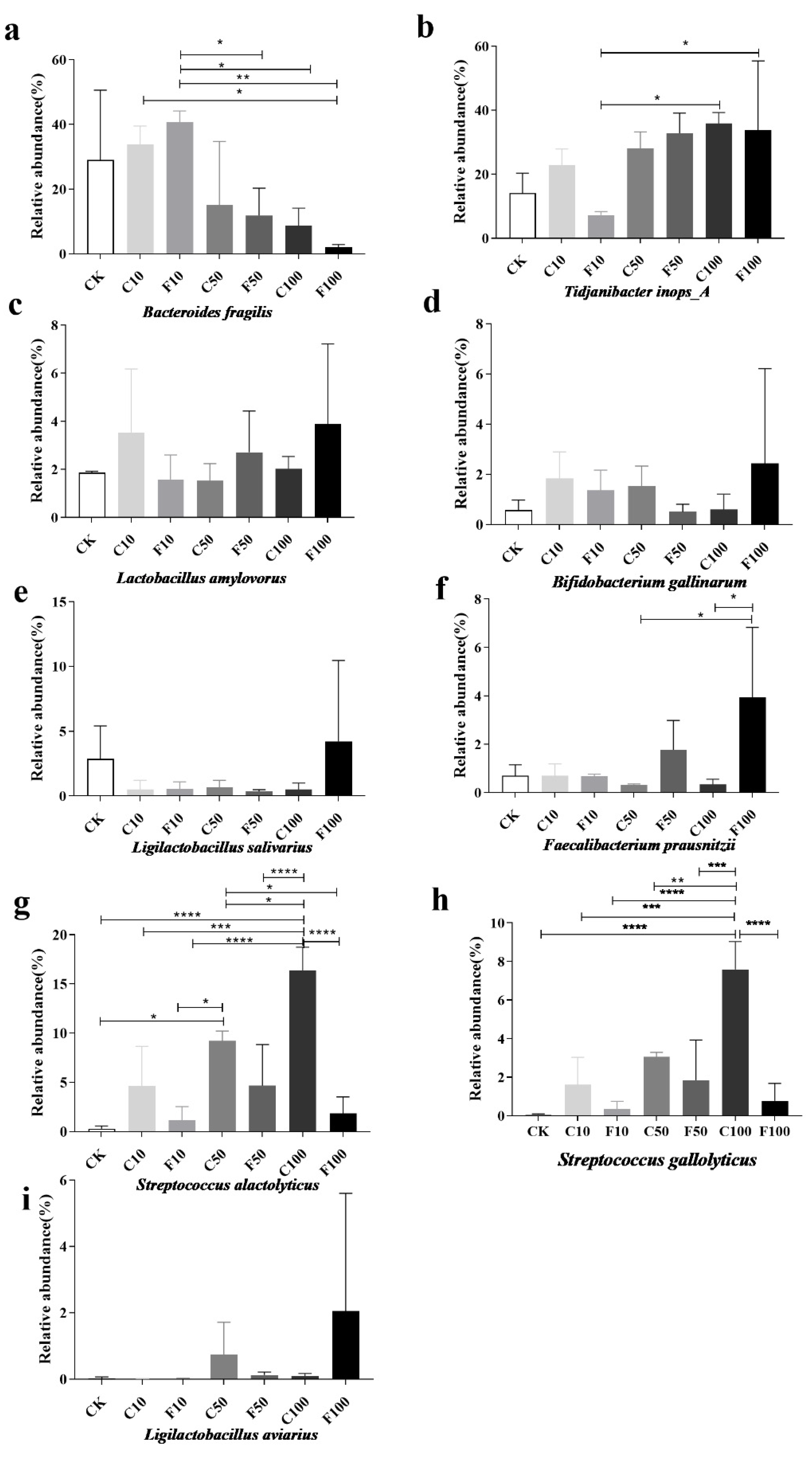
3.4. Effect of B. amyloliquefaciens D1 Fermentation of Soy Milk on the Structural Composition of Microbiota in Broiler Cecum Contents
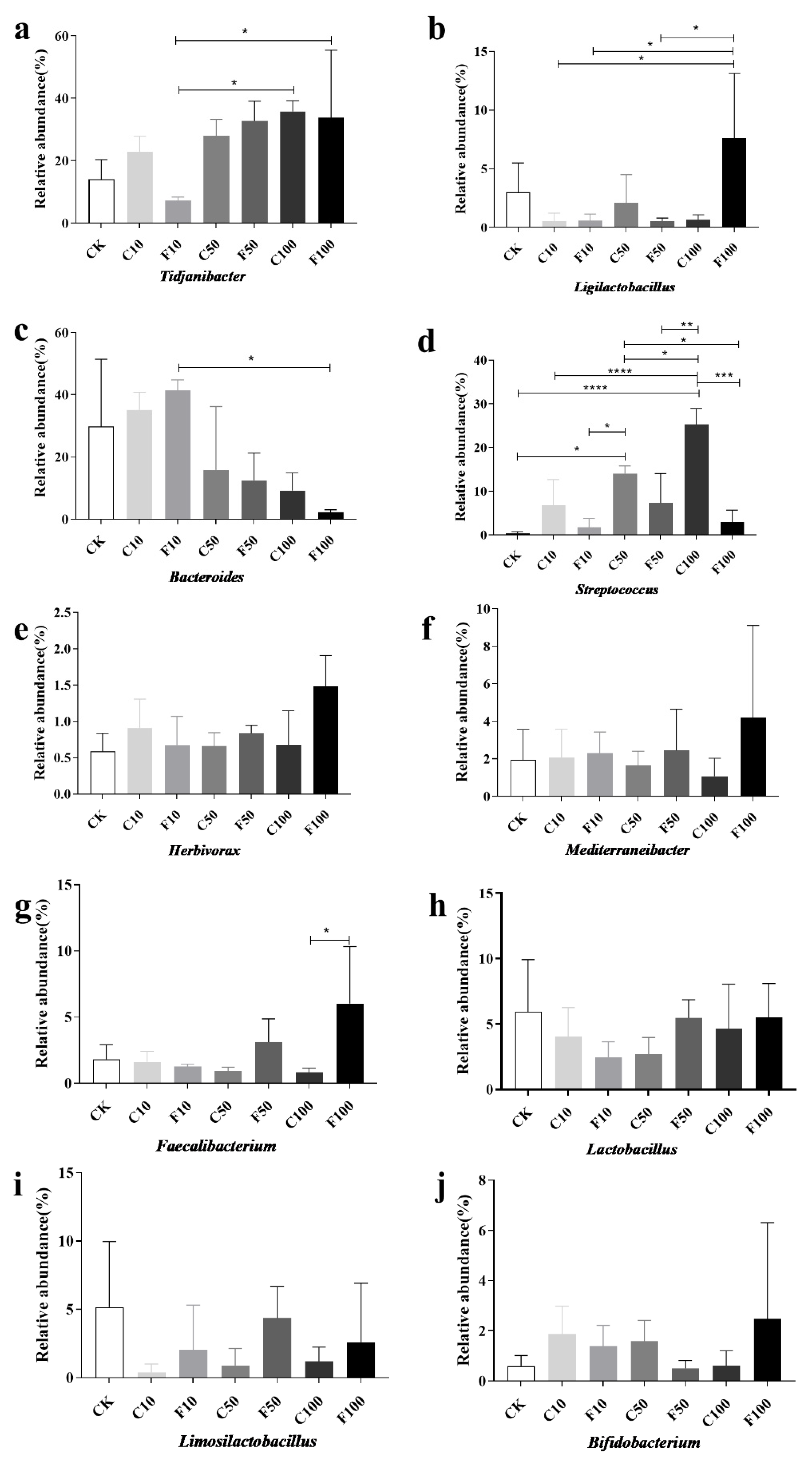
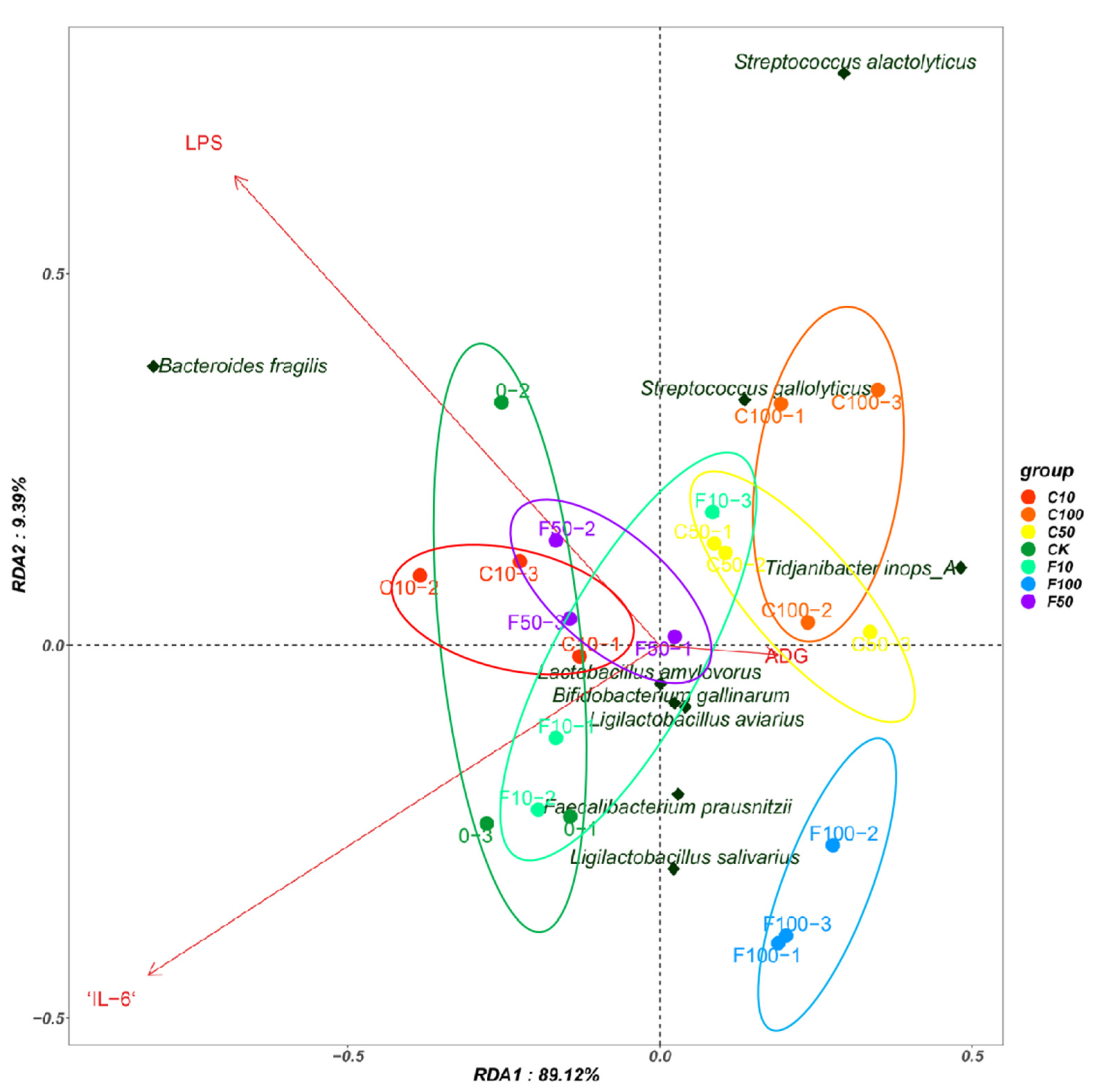
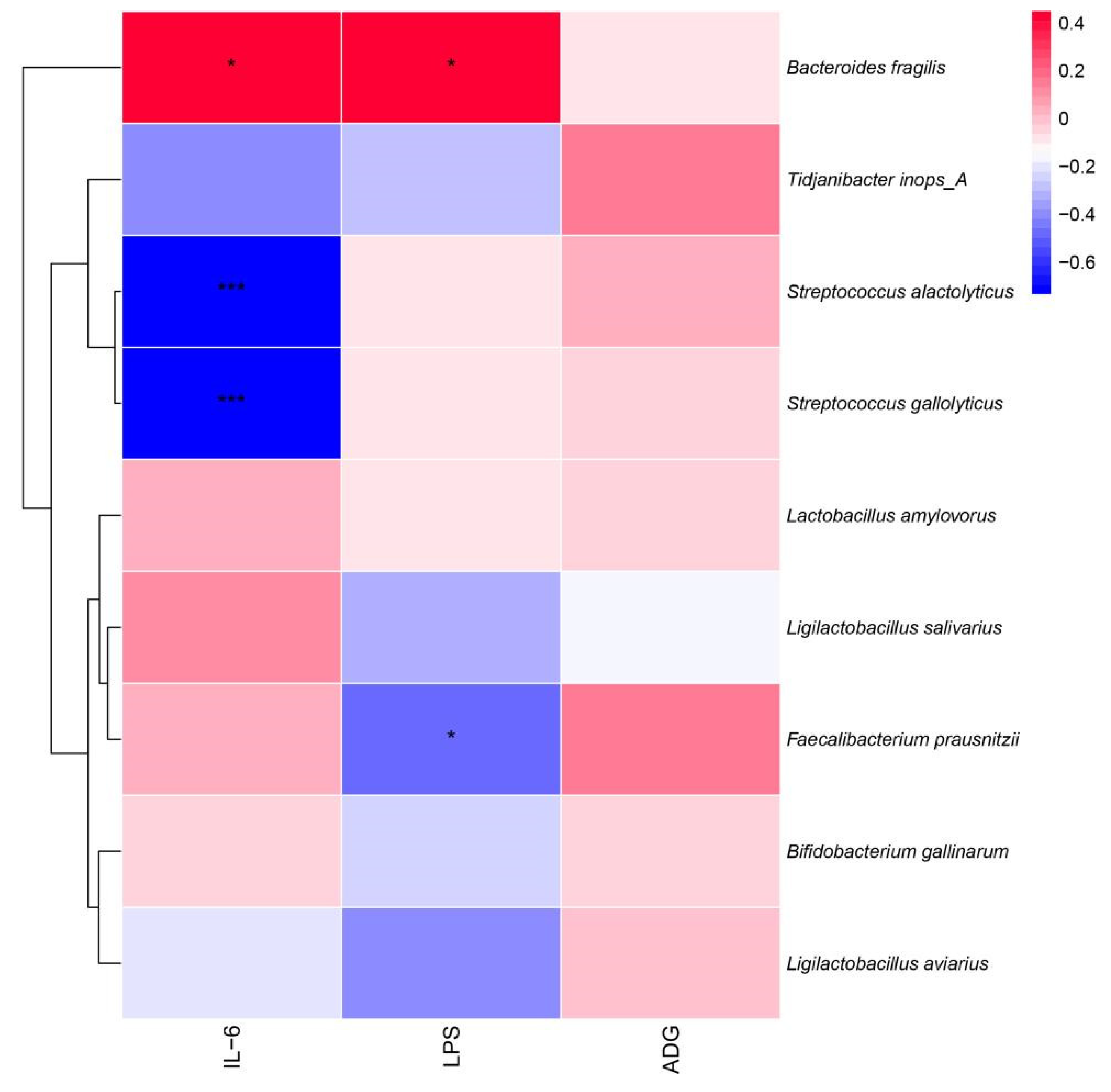
3.5. Correlation Analysis of Strains of Broiler Intestinal Microbiota Species Level with Immune Factors and ADG
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, G.T.; Zhan, X.A.; Zhang, L.L.; Zeng, X.F.; Chen, A.G.; Yang, C.M. Effects of dietary Bacillus amyloliquefaciens on mucosal immunity, cecal volatile fatty acids and microbial diversity in broiler chickens. Indian J. Anim. Res. 2018, 53, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Kim, S.Y.; Chang, H.I.; Park, E.; Paik, H.D. Immunomodulatory and Antigenotoxic Properties of Bacillus amyloliquefaciens KU801. Korean J. Microbiol. Biotechnol. 2013, 41, 249–252. [Google Scholar] [CrossRef]
- Hong, Y.; Cheng, Y.; Guan, L.; Zhou, Z.; Li, X.; Shi, D.; Xiao, Y. Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance. Microorganisms 2021, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Lei, X.; Wang, J.; Wang, L.; Zhong, Q.; Fang, X.; Li, P.; Du, B.; Wang, Y.; Liao, Z. Lipopolysaccharides derived from gram-negative bacterial pool of human gut microbiota promote inflammation and obesity development. Int. Rev. Immunol. 2022, 41, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Ghanima, M.; Kasrawy, N.; Aziz, A. Impact of Soymilk on Productive and Biochemical performances as well as Economic indices of Two Commercial Broiler Breeds. Alex. J. Vet. Sci. 2019, 60, 94. [Google Scholar] [CrossRef]
- Jayachandran, M.; Xu, B. An insight into the health benefits of fermented soy products. Food Chem. 2019, 271, 362–371. [Google Scholar] [CrossRef]
- Li, J.; Zhou, R.; Ren, Z.; Fan, Y.; Hu, S.; Zhuo, C.; Deng, Z. Improvement of protein quality and degradation of allergen in soybean meal fermented by Neurospora crassa. LWT 2019, 101, 220–228. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, B.; Wu, Z.; Wang, W.; Li, C.; Liu, G.; Cai, H. Effects of Fermented Soybean Meal Supplementation on the Growth Performance and Cecal Microbiota Community of Broiler Chickens. Animals 2020, 10, 1098. [Google Scholar] [CrossRef]
- Soumeh, E.A.; Mohebodini, H.; Toghyani, M.; Shabani, A.; Ashayerizadeh, A.; Jazi, V. Synergistic effects of fermented soybean meal and mannan-oligosaccharide on growth performance, digestive functions, and hepatic gene expression in broiler chickens. Poult. Sci. 2019, 98, 6797–6807. [Google Scholar] [CrossRef]
- Yuan, Y.; Peng, D.; Gu, X.; Gong, Y.; Sheng, Z.; Hu, X. Polygenic Basis and Variable Genetic Architectures Contribute to the Complex Nature of Body Weight—A Genome-Wide Study in Four Chinese Indigenous Chicken Breeds. Front. Genet. 2018, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Jiang, S.; Cheng, Z.; Ding, F.; Fan, Q.; Lin, X.; Wang, Y.; Gou, Z. Feed Restriction Improves Lipid Metabolism by Changing the Structure of the Cecal Microbial Community and Enhances the Meat Quality and Flavor of Bearded Chickens. Animals 2022, 12, 970. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Huang, Q.; Zhang, J.; He, J.; Zhang, L.; Wang, T. Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. Sci. 2017, 96, 74–82. [Google Scholar] [CrossRef]
- Ahmat, M.; Cheng, J.; Abbas, Z.; Cheng, Q.; Fan, Z.; Ahmad, B.; Hou, M.; Osman, G.; Guo, H.; Wang, J.; et al. Effects of Bacillus amyloliquefaciens LFB112 on Growth Performance, Carcass Traits, Immune, and Serum Biochemical Response in Broiler Chickens. Antibiotics 2021, 10, 1427. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.W.; Fang, C.L.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Molecular analysis of intestinal bacterial microbiota of broiler chickens fed diets containing fermented cottonseed meal. Poult. Sci. 2013, 92, 392–401. [Google Scholar] [CrossRef]
- Dinani, O.P.; Tyagi, P.K.; Tyagi, J.S.; Bhanja, S.K.; Rokade, J.J. Effect of feeding rice gluten meal with and without enzymes on hematobiochemical profile of broiler chickens. Vet. World 2020, 13, 2062–2069. [Google Scholar] [CrossRef]
- Saleh, A.A.; Shukry, M.; Farrag, F.; Soliman, M.M.; Abdel-Moneim, A.E. Effect of Feeding Wet Feed or Wet Feed Fermented by Bacillus licheniformis on Growth Performance, Histopathology and Growth and Lipid Metabolism Marker Genes in Broiler Chickens. Animals 2021, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Wei, Y.; Wang, X.; Ba, C.; Shi, W. Protective effect of Salvia miltiorrhiza polysaccharides on liver injury in chickens. Poult. Sci. 2019, 98, 3496–3503. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Cassilly, C.D.; Liu, X.; Park, S.M.; Tusi, B.K.; Chen, X.; Kwon, J.; Filipcik, P.; Bolze, A.S.; Liu, Z.; et al. Akkermansia muciniphila phospholipid induces homeostatic immune responses. Nature 2022, 608, 168–173. [Google Scholar] [CrossRef]
- Mazanko, M.S.; Popov, I.V.; Prazdnova, E.V.; Refeld, A.G.; Bren, A.B.; Zelenkova, G.A.; Chistyakov, V.A.; Algburi, A.; Weeks, R.M.; Ermakov, A.M.; et al. Beneficial Effects of Spore-Forming Bacillus Probiotic Bacteria Isolated From Poultry Microbiota on Broilers’ Health, Growth Performance, and Immune System. Front. Vet. Sci. 2022, 9, 877360. [Google Scholar] [CrossRef]
- Yitbarek, A.; Echeverry, H.; Munyaka, P.; Rodriguez-Lecompte, J.C. Innate immune response of pullets fed diets supplemented with prebiotics and synbiotics. Poult. Sci. 2015, 94, 1802–1811. [Google Scholar] [CrossRef]
- Gadde, U.; Oh, S.T.; Lee, Y.S.; Davis, E.; Zimmerman, N.; Rehberger, T.; Lillehoj, H.S. The Effects of Direct-fed Microbial Supplementation, as an Alternative to Antibiotics, on Growth Performance, Intestinal Immune Status, and Epithelial Barrier Gene Expression in Broiler Chickens. Probiotics Antimicrob. Proteins 2017, 9, 397–405. [Google Scholar] [CrossRef]
- Guo, M.; Li, M.; Zhang, C.; Zhang, X.; Wu, Y. Dietary Administration of the Bacillus subtilis Enhances Immune Responses and Disease Resistance in Chickens. Front. Microbiol. 2020, 11, 1768. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Y.P.; Yang, M.X.; Zhang, L.L.; Lu, Z.X.; Zhou, Y.M.; Wang, T. Bacillus amyloliquefaciens supplementation alleviates immunological stress and intestinal damage in lipopolysaccharide-challenged broilers. Anim. Feed Sci. Tech. 2015, 208, 119–131. [Google Scholar] [CrossRef]
- Torok, V.A.; Hughes, R.J.; Mikkelsen, L.L.; Perez-Maldonado, R.; Balding, K.; MacAlpine, R.; Percy, N.J.; Ophel-Keller, K. Identification and Characterization of Potential Performance-Related Gut Microbiotas in Broiler Chickens across Various Feeding Trials. Appl. Environ. Microb. 2011, 77, 5868–5878. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Wu, Z.; Li, T.; Liu, C.; Han, D.; Tao, S.; Pi, Y.; Li, N.; Wang, J. Perturbation of the lipid metabolism and intestinal inflammation in growing pigs with low birth weight is associated with the alterations of gut microbiota. Sci. Total Environ. 2020, 719, 137382. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Xie, Z.; Bai, Y.; Chen, G.; Rui, Y.; Chen, D.; Sun, Y.; Zeng, X.; Liu, Z. Modulation of gut homeostasis by exopolysaccharides from Aspergillus cristatus (MK346334), a strain of fungus isolated from Fuzhuan brick tea, contributes to immunomodulatory activity in cyclophosphamide-treated mice. Food Funct. 2020, 11, 10397–10412. [Google Scholar] [CrossRef]
- Miquel, S.; Martin, R.; Bridonneau, C.; Robert, V.; Sokol, H.; Bermudez-Humaran, L.G.; Thomas, M.; Langella, P. Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacterium prausnitzii. Gut Microbes 2014, 5, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Sun, L.; Liu, Y.; Ren, H.; Shen, Y.; Bi, F.; Zhang, T.; Wang, X. Alter between gut bacteria and blood metabolites and the anti-tumor effects of Faecalibacterium prausnitzii in breast cancer. BMC Microbiol. 2020, 20, 82. [Google Scholar] [CrossRef] [PubMed]
- Chaves, B.D.; Brashears, M.M.; Nightingale, K.K. Applications and safety considerations of Lactobacillus salivarius as a probiotic in animal and human health. J. Appl. Microbiol. 2017, 123, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Limaye, A.; Chang, H.; Liu, J. Screening of Lactic Acid Bacterial Strains with Antiviral Activity Against Porcine Epidemic Diarrhea. Probiotics Antimicrob. Proteins 2022, 14, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, W.K.A.; de Souza, G.T.; Brandão, L.R.; de Lima, R.C.; Garcia, E.F.; Dos Santos Lima, M.; de Souza, E.L.; Saarela, M.; Magnani, M. Exploiting antagonistic activity of fruit-derived Lactobacillus to control pathogenic bacteria in fresh cheese and chicken meat. Food Res. Int 2018, 108, 172–182. [Google Scholar] [CrossRef] [PubMed]
- García-Hernández, Y.; Pérez-Sánchez, T.; Boucourt, R.; Balcázar, J.L.; Nicoli, J.R.; Moreira-Silva, J.; Rodríguez, Z.; Fuertes, H.; Nuñez, O.; Albelo, N.; et al. Isolation, characterization and evaluation of probiotic lactic acid bacteria for potential use in animal production. Res. Vet. Sci 2016, 108, 125–132. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Zhao, Y.; Lin, Z.; Ji, L.; Ma, X. Tibetan Pig-Derived Probiotic Lactobacillus amylovorus SLZX20-1 Improved Intestinal Function via Producing Enzymes and Regulating Intestinal Microflora. Front. Nutr. 2022, 9, 846991. [Google Scholar] [CrossRef]
- Shankar KapnoorVeerappa, H.M. Production of α-Galactosidase by Aspergillus oryzae through Solid-state Fermentation and its Application in Soymilk Galactooligosaccharide Hydrolysis. Braz. Arch. Biol. Technol. 2010, 1, 211–218. [Google Scholar]
- An, K.; Gao, W.; Li, P.; Li, L.; Xia, Z. Dietary Lactobacillus plantarum improves the growth performance and intestinal health of Pekin ducks. Poult. Sci. 2022, 101, 101844. [Google Scholar] [CrossRef]
- Wang, B.; Gong, L.; Zhou, Y.; Tang, L.; Zeng, Z.; Wang, Q.; Zou, P.; Yu, D.; Li, W. Probiotic Paenibacillus polymyxa 10 and Lactobacillus plantarum 16 enhance growth performance of broilers by improving the intestinal health. Anim. Nutr. 2021, 7, 829–840. [Google Scholar] [CrossRef]
- Sekizaki, T.; Nishiya, H.; Nakajima, S.; Nishizono, M.; Kawano, M.; Okura, M.; Takamatsu, D.; Nishino, H.; Ishiji, T.; Osawa, R. Endocarditis in chickens caused by subclinical infection of Streptococcus gallolyticus subsp. gallolyticus. Avian Dis. 2008, 52, 183–186. [Google Scholar] [CrossRef]
- Zheng, L.; Li, D.; Li, Z.L.; Kang, L.N.; Jiang, Y.Y.; Liu, X.Y.; Chi, Y.P.; Li, Y.Q.; Wang, J.H. Effects of Bacillus fermentation on the protein microstructure and anti-nutritional factors of soybean meal. Lett. Appl. Microbiol. 2017, 65, 520–526. [Google Scholar] [CrossRef]
- Oehmcke-Hecht, S.; Mandl, V.; Naatz, L.T.; Dühring, L.; Köhler, J.; Kreikemeyer, B.; Maletzki, C. Streptococcus gallolyticus abrogates anti-carcinogenic properties of tannic acid on low-passage colorectal carcinomas. Sci. Rep. 2020, 10, 4714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokrani, M.; Charradi, K.; Limam, F.; Aouani, E.; Urdaci, M.C. Grape seed and skin extract, a potential prebiotic with anti-obesity effect through gut microbiota modulation. Gut Pathog. 2022, 14, 30. [Google Scholar] [CrossRef]
- Zhang, Y.; Weng, Y.; Gan, H.; Zhao, X.; Zhi, F. Streptococcus gallolyticus conspires myeloid cells to promote tumorigenesis of inflammatory bowel disease. Biochem. Biophys. Res. Commun. 2018, 506, 907–911. [Google Scholar] [CrossRef]
- Hanifi, A.; Culpepper, T.; Mai, V.; Anand, A.; Ford, A.L.; Ukhanova, M.; Christman, M.; Tompkins, T.A.; Dahl, W.J. Evaluation of Bacillus subtilis R0179 on gastrointestinal viability and general wellness: A randomised, double-blind, placebo-controlled trial in healthy adults. Benef. Microbes 2015, 6, 19–27. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Number | Intervention Method | Interventions | Dosage (mL/kg) | Intervention Period (d) |
|---|---|---|---|---|---|
| F10 | 6 | Mixing | Fermented soy milk | 100 | 35 |
| F50 | 6 | Mixing | Fermented soy milk | 500 | 35 |
| F100 | 6 | Mixing | Fermented soy milk | 1000 | 35 |
| C10 | 6 | Mixing | Unfermented soy milk | 100 | 35 |
| C50 | 6 | Mixing | Unfermented soy milk | 500 | 35 |
| C100 | 6 | Mixing | Unfermented soy milk | 1000 | 35 |
| CK | 6 | - | - | - | - |
| Projects | CK | F-10 | C-10 | F-50 | C-50 | F-100 | C-100 |
|---|---|---|---|---|---|---|---|
| Initial weight (g/bird) | 48.29 | 53.62 | 52.83 | 47.95 | 52.35 | 54.53 | 55.33 |
| Average daily feed intake (ADFI)(g/d/bird) | 38.72 | 49.84 | 51.99 | 48.69 | 49.25 | 44.86 | 45.73 |
| Average daily gain (ADG) (g/d/bird) | 16.18 | 20.28 | 19.13 | 20.53 | 20.15 | 19.39 | 18.32 |
| Average feed/Gain ratio (F/G) | 2.39 | 2.46 | 2.72 | 2.37 | 2.44 | 2.31 | 2.50 |
| Projects | CK | F-10 | C-10 | F-50 | C-50 | F-100 | C-100 |
|---|---|---|---|---|---|---|---|
| Total protein (TP) (g/L) | 33.33 | 32.00 | 34.33 | 32.67 | 29.67 | 32.67 | 27.67 |
| Albumin (ALB) (g/L) | 14.20 | 13.50 | 14.43 | 14.17 | 11.93 | 13.63 | 11.20 |
| Globulin (GLOB) (g/L) | 19.13 | 18.50 | 19.90 | 18.50 | 17.73 | 19.03 | 16.47 |
| Albumin/Globulin (A/G) | 0.74 | 0.73 | 0.73 | 0.77 | 0.67 | 0.72 | 0.68 |
| Alanine aminotransferase(ALT) (U/L) | - | - | - | - | - | - | - |
| Aspartate aminotransferase (AST) (U/L) | 208.33 | 182.00 | 195.67 | 202.33 | 180.00 | 232.00 | 192.67 |
| ALT/AST | |||||||
| Triglyceride (TG) (mmol/L) | 0.37 | 0.36 | 0.31 | 0.28 | 0.37 | 0.36 | 0.33 |
| Projects | CK | C10 | F10 | C50 | F50 | C100 |
|---|---|---|---|---|---|---|
| Chao1 Index | 9375.36 | 7116.31 | 6358.51 | 10,185.75 | 7144.04 | 7715.71 |
| Richness Index | 7937.67 | 3535.00 | 5182.33 | 8232.00 | 5006.67 | 4002.67 |
| Shannon Index | 9.83 | 7.35 | 8.38 | 9.49 | 8.76 | 7.07 |
| Simpson Index | 0.050 | 0.076 | 0.069 | 0.055 | 0.065 | 0.098 |
| ACE Index | 9837.41 | 8412.53 | 6901.73 | 10,976.78 | 8272.03 | 8926.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, L.; Chen, W.; Wang, J.; Huang, L.; Zheng, Q.; Chen, J.; Wang, L.; Cai, C.; Zhang, X.; Wang, L.; et al. Beneficial Effects of Bacillus amyloliquefaciens D1 Soy Milk Supplementation on Serum Biochemical Indexes and Intestinal Health of Bearded Chickens. Microorganisms 2023, 11, 1660. https://doi.org/10.3390/microorganisms11071660
Du L, Chen W, Wang J, Huang L, Zheng Q, Chen J, Wang L, Cai C, Zhang X, Wang L, et al. Beneficial Effects of Bacillus amyloliquefaciens D1 Soy Milk Supplementation on Serum Biochemical Indexes and Intestinal Health of Bearded Chickens. Microorganisms. 2023; 11(7):1660. https://doi.org/10.3390/microorganisms11071660
Chicago/Turabian StyleDu, Liyu, Weizhe Chen, Jie Wang, Lingzhu Huang, Qikai Zheng, Junjie Chen, Linhao Wang, Changyu Cai, Xiangbin Zhang, Li Wang, and et al. 2023. "Beneficial Effects of Bacillus amyloliquefaciens D1 Soy Milk Supplementation on Serum Biochemical Indexes and Intestinal Health of Bearded Chickens" Microorganisms 11, no. 7: 1660. https://doi.org/10.3390/microorganisms11071660