

Diversity of Endophytic Microbes in Taxus yunnanensis and Their Potential for Plant Growth Promotion and Taxane Accumulation
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sampling
2.2. Illumina-Based Analysis of Endophytic Microbes
2.3. Isolation and Identification of Endophytes
2.4. Growth-Promoting Characteristics of Endophytic Bacteria
2.5. Plant Experiment
2.6. Effects of Endophytic Elicitor on the Accumulation of Taxanes in Stem Cells of T. yunnanensis
2.7. Statistical Analysis
3. Results
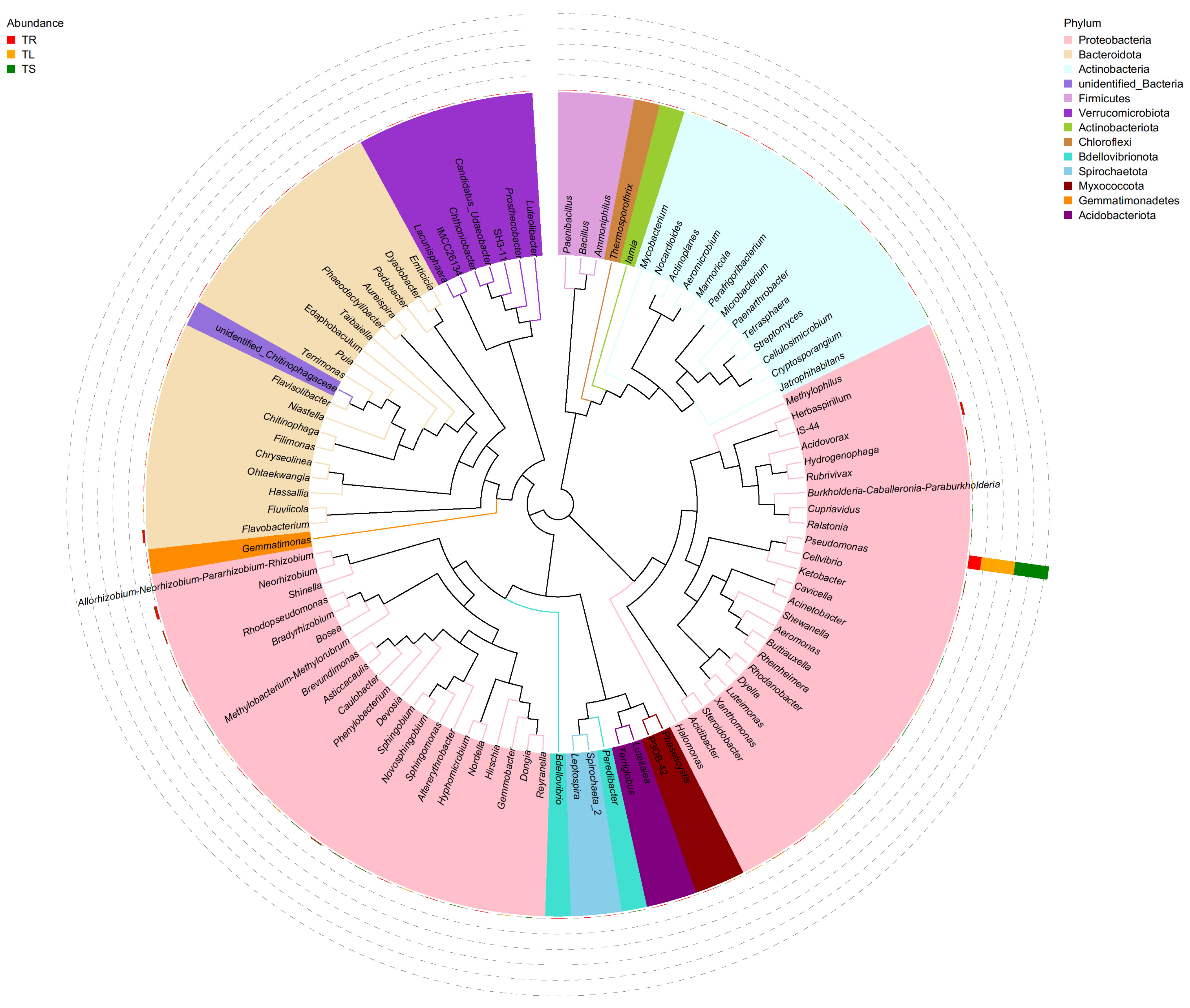
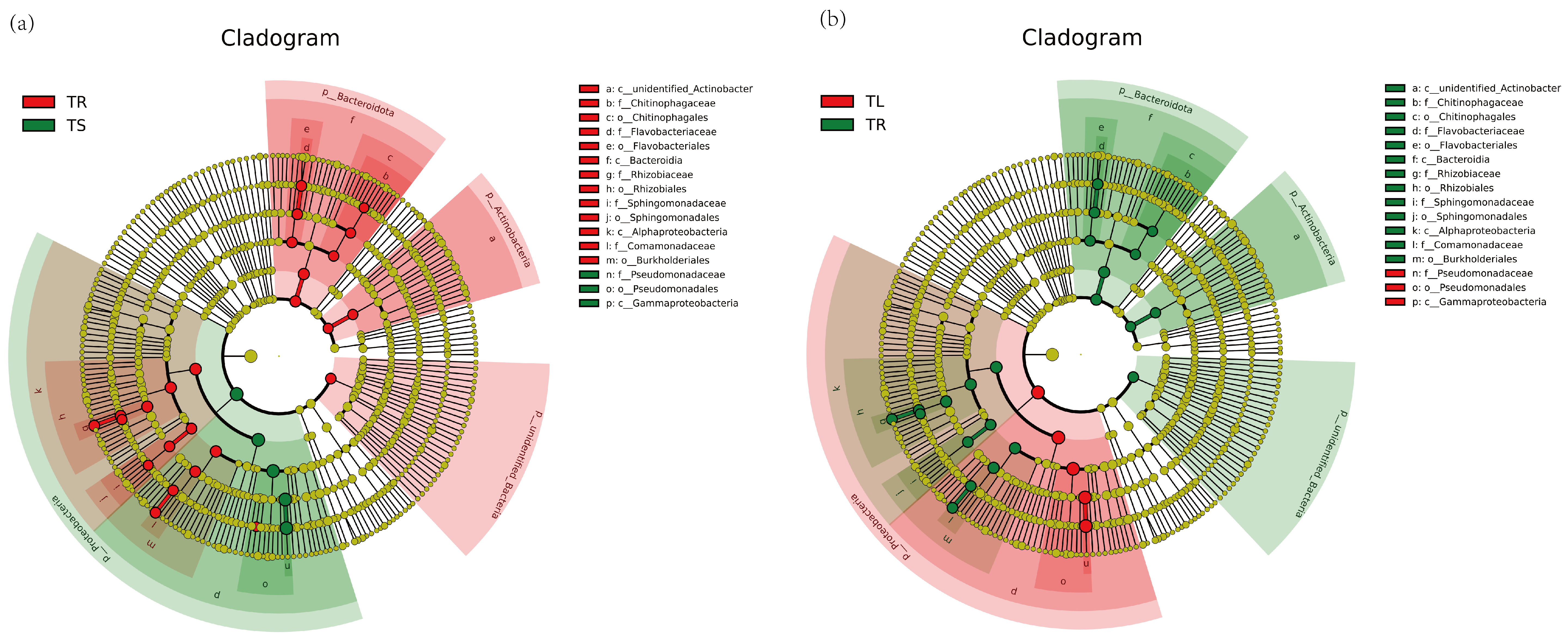
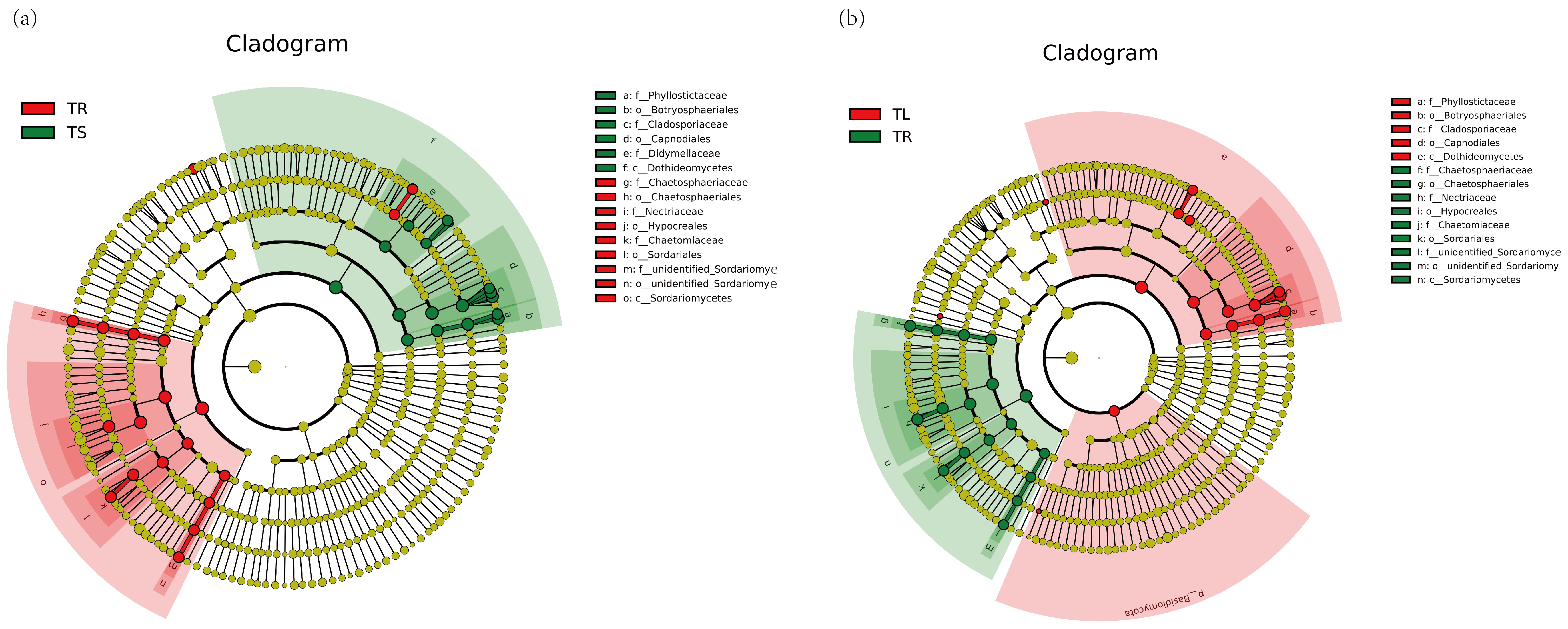
3.1. Endophyte Community Diversity Analysis
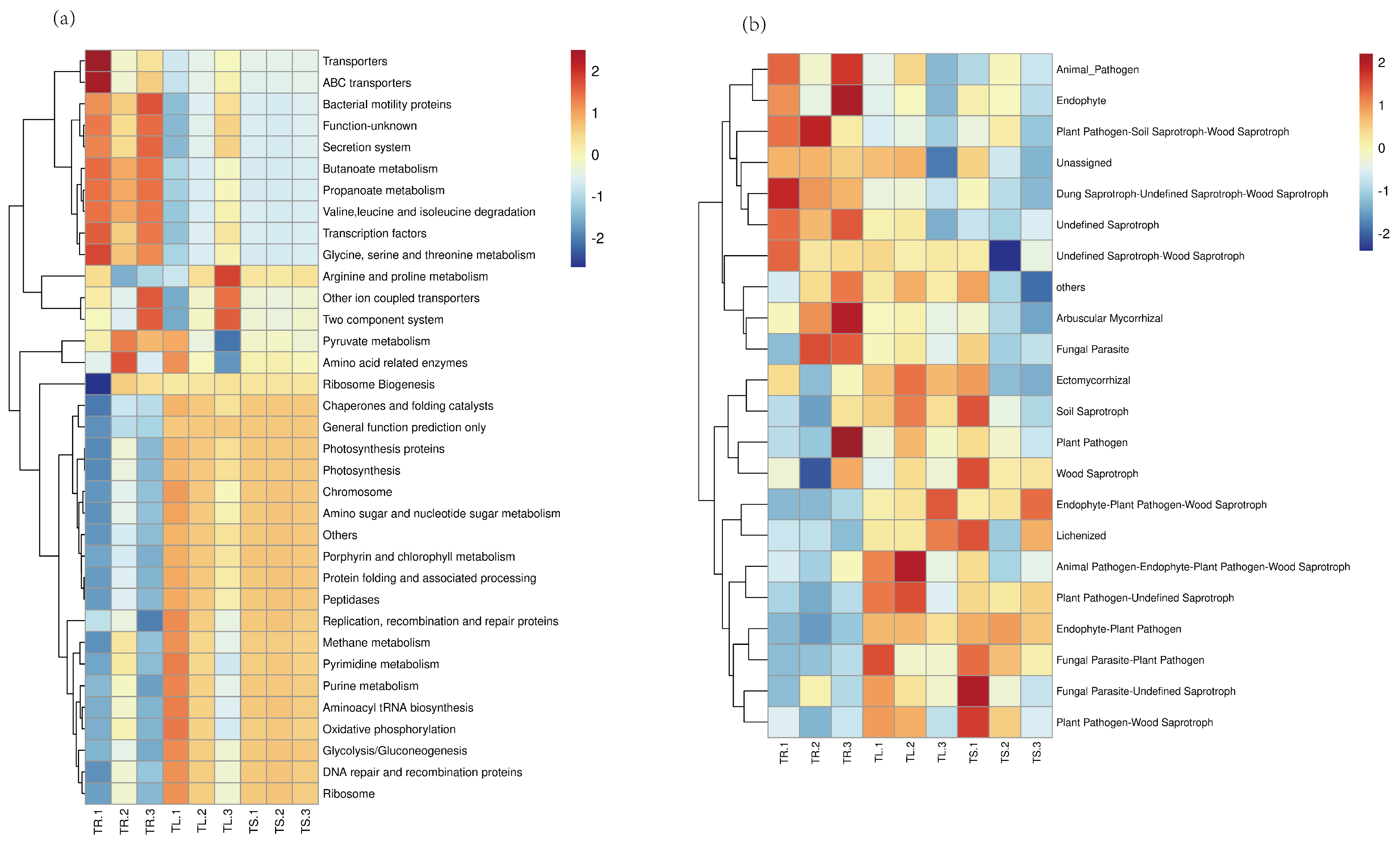
3.2. Function Prediction of Endophytes
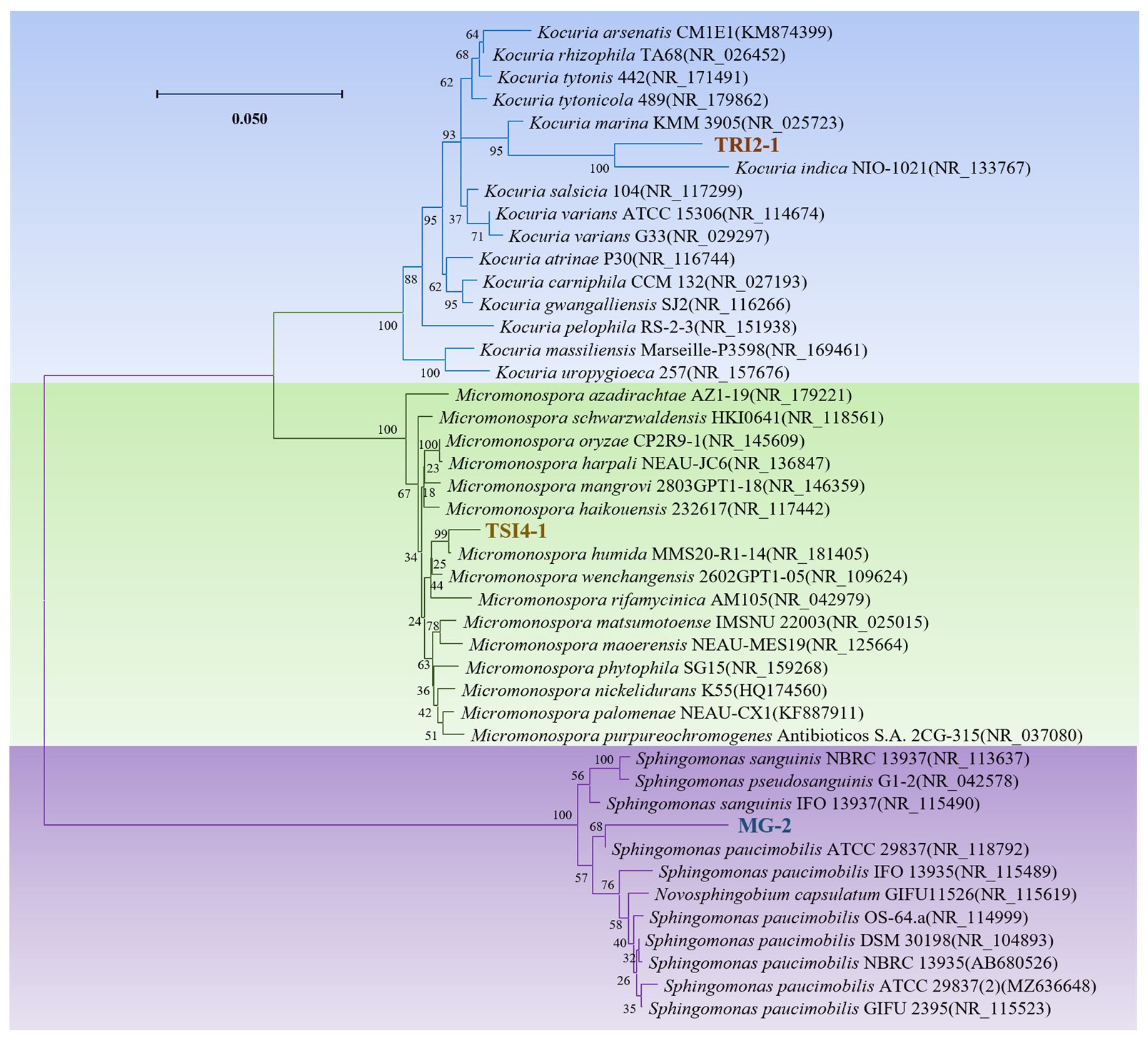
3.3. Isolation of Culturable Endophytes
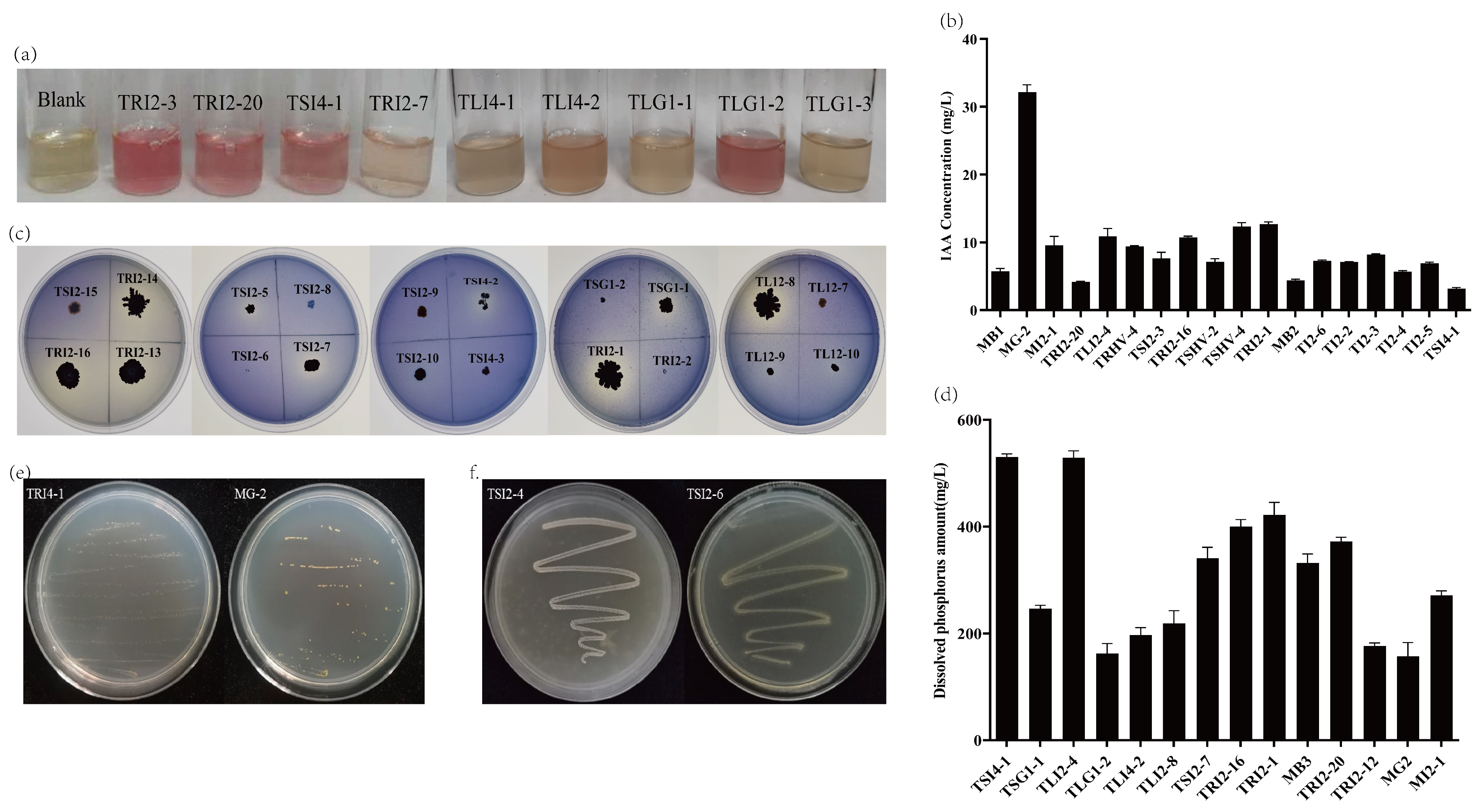
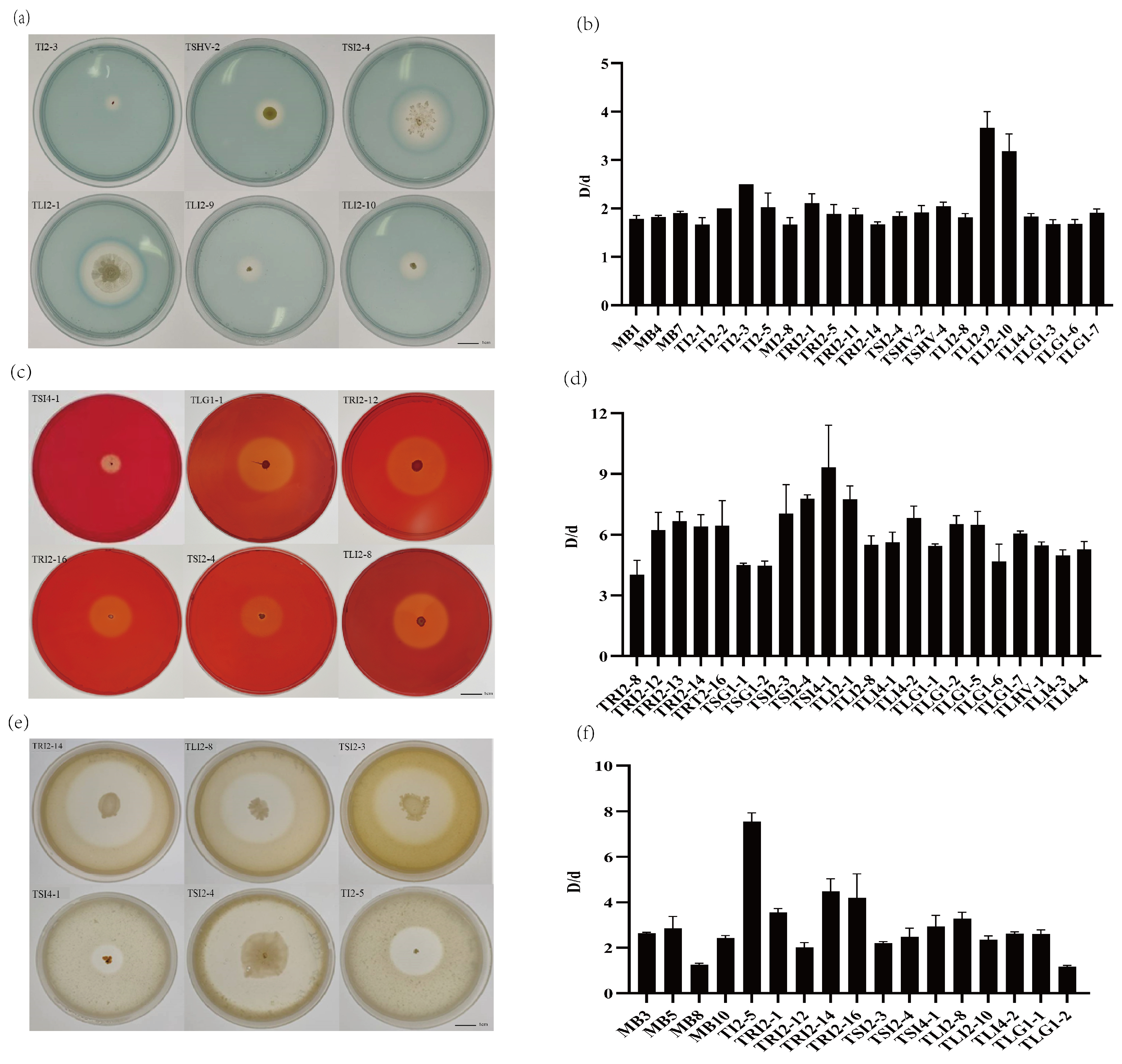
3.4. Growth-Promoting Characteristics of Endophytic Bacteria
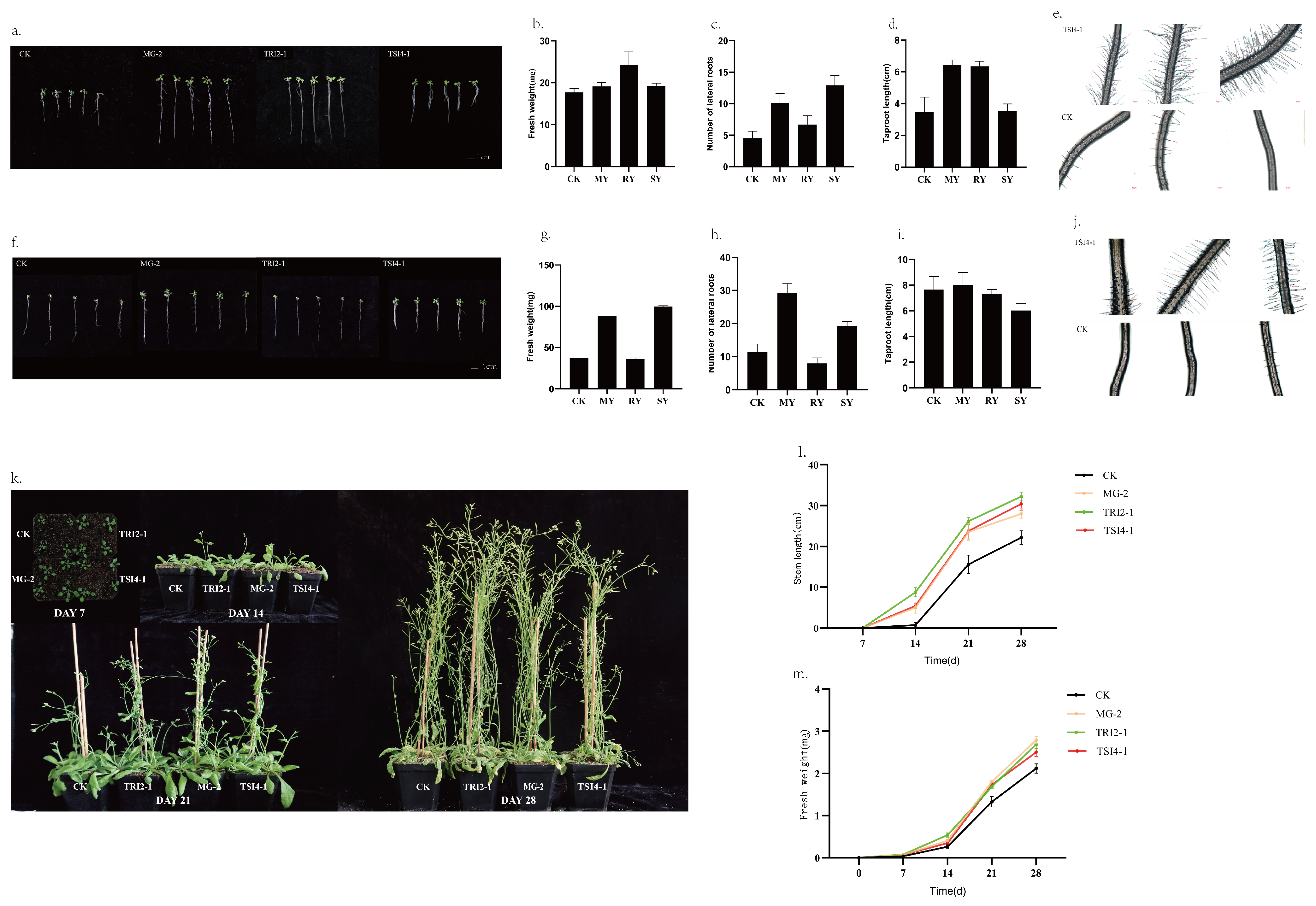
3.5. Effects of MG-2, TRI2-1, and TSI4-1 on the Growth of Arabidopsis
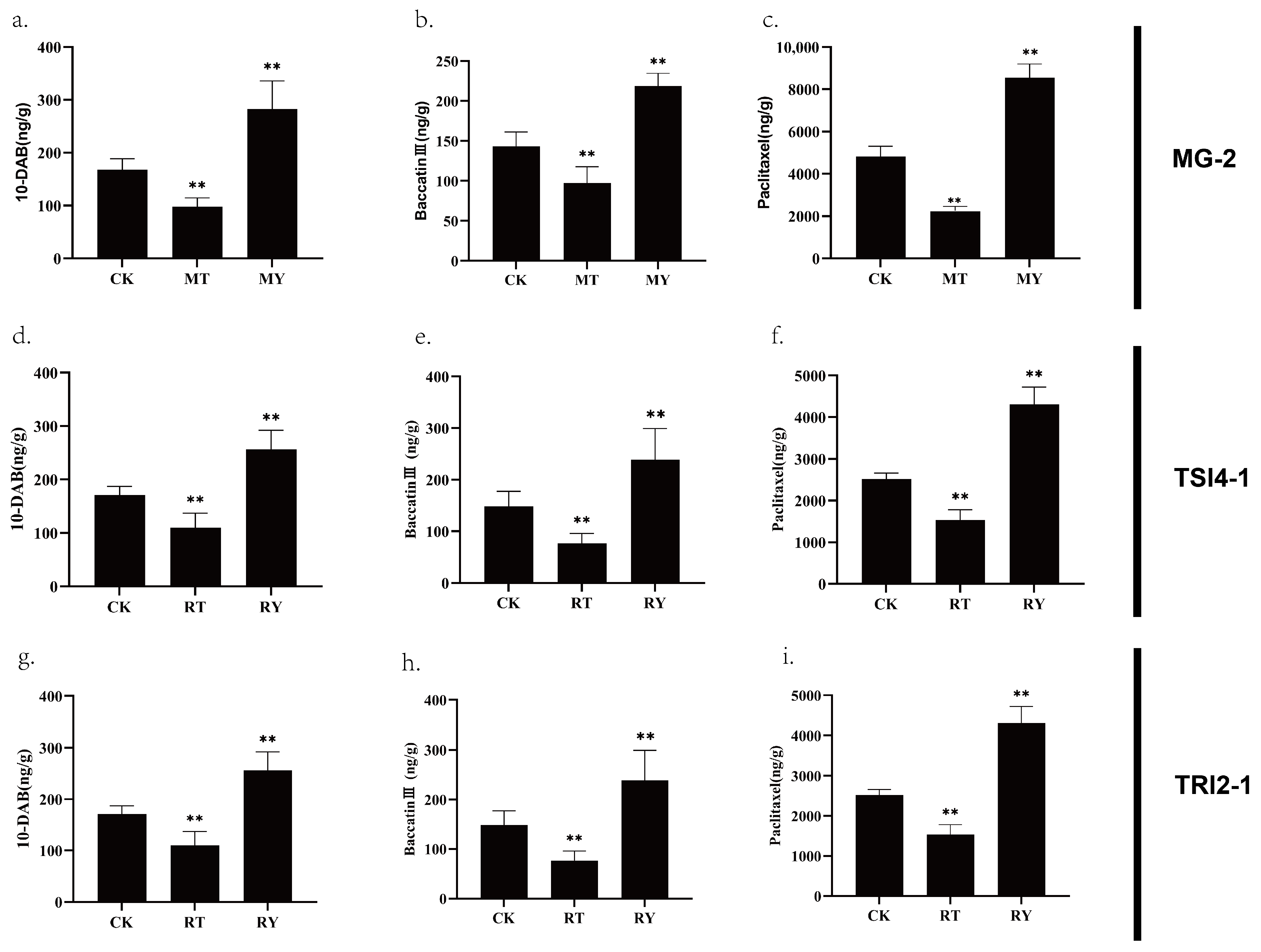
3.6. The Effects of MG-2, TRI2-1, and TSI4-1 on the Accumulation of Taxanes in T. yunnanensis Stem Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, X.-L.; Zhan, X.-R. Research Progress on the Dormancy Mechanism of Taxus Seeds. Chin. Med. J. Res. Prac. 2018, 32, 78–81. [Google Scholar] [CrossRef]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of Natural Products on Developing New Anti-Cancer Agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Rustamova, N.; Bozorov, K.; Efferth, T.; Yili, A. Novel Secondary Metabolites from Endophytic Fungi: Synthesis and Biological Properties. Phytochem. Rev. 2020, 19, 425–448. [Google Scholar] [CrossRef]
- Manish, L.; Mangesh, B.; Sabale, V. Taxus—The Panacea for Cancer Treatment. Anc. Sci. Life 2005, 24, 152–159. [Google Scholar] [PubMed]
- Nicolaou, K.C.; Yang, Z.; Liu, J.J.; Ueno, H.; Nantermet, P.G.; Guy, R.K.; Claiborne, C.F.; Renaud, J.; Couladouros, E.A.; Paulvannan, K.; et al. Total Synthesis of Taxol. Nature 1994, 367, 630–634. [Google Scholar] [CrossRef]
- Kanda, Y.; Nakamura, H.; Umemiya, S.; Puthukanoori, R.K.; Murthy Appala, V.R.; Gaddamanugu, G.K.; Paraselli, B.R.; Baran, P.S. Two-Phase Synthesis of Taxol. J. Am. Chem. Soc. 2020, 142, 10526–10533. [Google Scholar] [CrossRef]
- Ketchum, R.E.B.; Gibson, D.M.; Croteau, R.B.; Shuler, M.L. The Kinetics of Taxoid Accumulation in Cell Suspension Cultures of Taxus Following Elicitation with Methyl Jasmonate. Biotechnol. Bioeng. 1999, 62, 97–105. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Mao, J.-W.; Tan, X.-L. Research Progress on the Source, Production, and Anti-Cancer Mechanisms of Paclitaxel. Chin. J. Nat. Med. 2020, 18, 890–897. [Google Scholar] [CrossRef]
- Liu, W.C.; Gong, T.; Zhu, P. Advances in Exploring Alternative Taxol Sources. RSC Adv. 2016, 6, 48800–48809. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.L.; Zhang, Q.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K.P. Review: Endophytic Microbes and Their Potential Applications in Crop Management. Pest. Manag. Sci. 2019, 75, 2558–2565. [Google Scholar] [CrossRef]
- Singh, S.; Pandey, A. Plant-Associated Microbial Endophytes: Promising Source for Bioprospecting. In Mining of Microbial Wealth and MetaGenomics; Kalia, V.C., Shouche, Y., Purohit, H.J., Rahi, P., Eds.; Springer: Singapore, 2017; pp. 249–265. ISBN 978-981-10-5708-3. [Google Scholar]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. Isolation and Characterization of Plant Growth-Promoting Endophytic Bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. Biomed Res. Int. 2020, 2020, 8650957. [Google Scholar] [CrossRef] [Green Version]
- Abedinzadeh, M.; Etesami, H.; Alikhani, H.A. Characterization of Rhizosphere and Endophytic Bacteria from Roots of Maize (Zea mays L.) Plant Irrigated with Wastewater with Biotechnological Potential in Agriculture. Biotechnol. Rep. 2019, 21, e00305. [Google Scholar] [CrossRef]
- Weilharter, A.; Mitter, B.; Shin, M.V.; Chain, P.S.G.; Nowak, J.; Sessitsch, A. Complete Genome Sequence of the Plant Growth-Promoting Endophyte Burkholderia Phytofirmans Strain PsJN. J. Bacteriol. 2011, 193, 3383–3384. [Google Scholar] [CrossRef] [Green Version]
- Borah, A.; Das, R.; Mazumdar, R.; Thakur, D. Culturable Endophytic Bacteria of Camellia Species Endowed with Plant Growth Promoting Characteristics. J. Appl. Microbiol. 2019, 127, 825–844. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, P.; Sharma, A.; Guo, D.-J.; Upadhyay, S.K.; Song, Q.-Q.; Verma, K.K.; Li, D.-P.; Malviya, M.K.; Song, X.-P.; et al. Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth. Int. J. Mol. Sci. 2022, 23, 6242. [Google Scholar] [CrossRef]
- Dubey, A.; Malla, M.A.; Kumar, A.; Dayanandan, S.; Khan, M.L. Plants Endophytes: Unveiling Hidden Agenda for Bioprospecting toward Sustainable Agriculture. Crit. Rev. Biotechnol. 2020, 40, 1210–1231. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic Potential of Endophytic Bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Zhang, S.; Zhang, J.; Ma, P.; Duan, J.; Liang, Z. Effect and Mechanism of Endophytic Bacteria on Growth and Secondary Metabolite Synthesis in Salvia miltiorrhiza Hairy Roots. Acta Physiol. Plant. 2014, 36, 1095–1105. [Google Scholar] [CrossRef]
- Lòpez-Fernàndez, S.; Compant, S.; Vrhovsek, U.; Bianchedi, P.L.; Sessitsch, A.; Pertot, I.; Campisano, A. Grapevine Colonization by Endophytic Bacteria Shifts Secondary Metabolism and Suggests Activation of Defense Pathways. Plant Soil 2016, 405, 155–175. [Google Scholar] [CrossRef]
- Kilam, D.; Saifi, M.; Abdin, M.Z.; Agnihotri, A.; Varma, A. Endophytic Root Fungus Piriformospora indica Affects Transcription of Steviol Biosynthesis Genes and Enhances Production of Steviol Glycosides in Stevia rebaudiana. Physiol. Mol. Plant Pathol. 2017, 97, 40–48. [Google Scholar] [CrossRef]
- Zheng, L.P.; Tian, H.; Yuan, Y.F.; Wang, J.W. The Influence of Endophytic Penicillium oxalicum B4 on Growth and Artemisinin Biosynthesis of in Vitro Propagated Plantlets of Artemisia annua L. Plant Growth Regul. 2016, 80, 93–102. [Google Scholar] [CrossRef]
- Wang, C.; Wu, J.; Mei, X. Enhancement of Taxol Production and Excretion in Taxus Chinensis Cell Culture by Fungal Elicitation and Medium Renewal. Appl. Microbiol. Biotechnol. 2001, 55, 404–410. [Google Scholar] [CrossRef]
- Li, Y.-C.; Tao, W.-Y.; Cheng, L. Paclitaxel Production Using Co-Culture of Taxus Suspension Cells and Paclitaxel-Producing Endophytic Fungi in a Co-Bioreactor. Appl. Microbiol. Biotechnol. 2009, 83, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xu, L.; Wang, J.; Dong, M.; Xu, C.; Kai, G.; Wan, W.; Jiang, J. Endophytic Fungus Pseudodidymocyrtis lobariellae KL27 Promotes Taxol Biosynthesis and Accumulation in Taxus chinensis. BMC Plant Biol. 2022, 22, 12. [Google Scholar] [CrossRef] [PubMed]
- Gallego, A.; Imseng, N.; Bonfill, M.; Cusido, R.M.; Palazon, J.; Eibl, R.; Moyano, E. Development of a Hazel Cell Culture-Based Paclitaxel and Baccatin III Production Process on a Benchtop Scale. J. Biotechnol. 2015, 195, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Wanke, U.; Draeger, S.; Aust, H.-J. Endophytes from Herbaceous Plants and Shrubs: Effectiveness of Surface Sterilization Methods. Mycol. Res. 1993, 97, 1447–1450. [Google Scholar] [CrossRef]
- Kim, M.; Morrison, M.; Yu, Z. Evaluation of Different Partial 16S RRNA Gene Sequence Regions for Phylogenetic Analysis of Microbiomes. J. Microbiol. Methods 2011, 84, 81–87. [Google Scholar] [CrossRef]
- Parfitt, D.; Hunt, J.; Dockrell, D.; Rogers, H.J.; Boddy, L. Do All Trees Carry the Seeds of Their Own Destruction? PCR Reveals Numerous Wood Decay Fungi Latently Present in Sapwood of a Wide Range of Angiosperm Trees. Fungal Ecol. 2010, 3, 338–346. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors that Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Sheng, X.-F.; Xia, J.-J.; Jiang, C.-Y.; He, L.-Y.; Qian, M. Characterization of Heavy Metal-Resistant Endophytic Bacteria from Rape (Brassica napus) Roots and Their Potential in Promoting the Growth and Lead Accumulation of Rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Mehta, S.; Nautiyal, C.S. An Efficient Method for Qualitative Screening of Phosphate-Solubilizing Bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B.R. Methods for Isolating and Characterizing ACC Deaminase-Containing Plant Growth-Promoting Rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Roy, N. Screening, Purification and Characterization of Cellulase from Cellulase Producing Bacteria in Molasses. BMC Res. Notes 2018, 11, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajpai, A.; Johri, B.N. Endophytic Pseudomonads and Their Metabolites. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Switzerland, 2019; pp. 33–59. ISBN 978-3-319-90484-9. [Google Scholar]
- Niem, J.M.; Billones-Baaijens, R.; Stodart, B.; Savocchia, S. Diversity Profiling of Grapevine Microbial Endosphere and Antagonistic Potential of Endophytic Pseudomonas Against Grapevine Trunk Diseases. Front. Microbiol. 2020, 11, 477. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, H.; Zhang, T.; Sun, J.; Lou, K. Illumina-Based Analysis of Endophytic Bacterial Diversity and Space-Time Dynamics in Sugar Beet on the North Slope of Tianshan Mountain. Appl. Microbiol. Biotechnol. 2014, 98, 6375–6385. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, Z.; Li, X.; Liu, S.; Song, F.; Liu, F. Relationship between Endophytic Microbial Diversity and Grain Quality in Wheat Exposed to Multi-Generational CO2 Elevation. Sci. Total Environ. 2021, 776, 146029. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Wang, S.-S.; Liu, J.-M.; Sun, J.; Sun, Y.-F.; Liu, J.-N.; Jia, N.; Fan, B.; Dai, X.-F. Diversity of Culture-Independent Bacteria and Antimicrobial Activity of Culturable Endophytic Bacteria Isolated from Different Dendrobium Stems. Sci. Rep. 2019, 9, 10389. [Google Scholar] [CrossRef] [Green Version]
- Rashmi, M. A Worldwide List of Endophytic Fungi with Notes on Ecology and Diversity. Mycosphere 2019, 10, 798–1079. [Google Scholar] [CrossRef]
- Riva, V.; Mapelli, F.; Bagnasco, A.; Mengoni, A.; Borin, S. A Meta-Analysis Approach to Defining the Culturable Core of Plant Endophytic Bacterial Communities. Appl. Environ. Microbiol. 2022, 88, e02537-21. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Javed, M.A.; Afridi, M.S.; Abbasi, H.A.; Qayyum, A.; Batool, T.; Ullah, A.; Marc, R.A.; Jaouni, S.K.A.; et al. Role of Endophytic Bacteria in Salinity Stress Amelioration by Physiological and Molecular Mechanisms of Defense: A Comprehensive Review. S. Afr. J. Bot. 2022, 151, 33–46. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas aeruginosa L10: A Hydrocarbon-Degrading, Biosurfactant-Producing, and Plant-Growth-Promoting Endophytic Bacterium Isolated from a Reed (Phragmites australis). Front. Microbiol. 2018, 9, 1087. [Google Scholar] [CrossRef] [Green Version]
- Guttenberger, N.; Blankenfeldt, W.; Breinbauer, R. Recent Developments in the Isolation, Biological Function, Biosynthesis, and Synthesis of Phenazine Natural Products. Bioorganic Med. Chem. 2017, 25, 6149–6166. [Google Scholar] [CrossRef]
- Reis, R.S.; Pereira, A.G.; Neves, B.C.; Freire, D.M.G. Gene Regulation of Rhamnolipid Production in Pseudomonas aeruginosa—A Review. Bioresour. Technol. 2011, 102, 6377–6384. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Munder, A.; Aravind, R.; Eapen, S.J.; Tümmler, B.; Raaijmakers, J.M. Friend or Foe: Genetic and Functional Characterization of Plant Endophytic Pseudomonas aeruginosa. Environ. Microbiol. 2013, 15, 764–779. [Google Scholar] [CrossRef]
- Kusam Lata, R.; Divjot, K.; Tanvir, K.; Rubee, D.; Ashok, Y.; Ajar Nath, Y. Bioprospecting of Endophytic Bacteria from the Indian Himalayas and Their Role in Plant Growth Promotion of Maize (Zea mays L.). J. App. Biol. Biotech. 2021, 9, 41–50. [Google Scholar] [CrossRef]
- Liu, J.-Q.; Chen, S.-M.; Zhang, C.-M.; Xu, M.-J.; Xing, K.; Li, C.-G.; Li, K.; Zhang, Y.-Q.; Qin, S. Abundant and Diverse Endophytic Bacteria Associated with Medicinal Plant Arctium lappa L. and Their Potential for Host Plant Growth Promoting. Antonie Leeuwenhoek 2022, 115, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Ashajyothi, M.; Kumar, A.; Sheoran, N.; Ganesan, P.; Gogoi, R.; Subbaiyan, G.K.; Bhattacharya, R. Black Pepper (Piper nigrum L.) Associated Endophytic Pseudomonas Putida BP25 Alters Root Phenotype and Induces Defense in Rice (Oryza sativa L.) against Blast Disease Incited by Magnaporthe Oryzae. Biol. Control. 2020, 143, 104181. [Google Scholar] [CrossRef]
- Shan, W.; Zhou, Y.; Liu, H.; Yu, X. Endophytic Actinomycetes from Tea Plants (Camellia sinensis): Isolation, Abundance, Antimicrobial, and Plant-Growth-Promoting Activities. BioMed Res. Int. 2018, 2018, e1470305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanan, Z.; Glick, B.R. Secondary Metabolites Produced by Plant Growth-Promoting Bacterial Endophytes. Microorganisms 2022, 10, 2008. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and Interactions in Plant Growth Promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [Green Version]
- Dahmani, M.A.; Desrut, A.; Moumen, B.; Verdon, J.; Mermouri, L.; Kacem, M.; Coutos-Thévenot, P.; Kaid-Harche, M.; Bergès, T.; Vriet, C. Unearthing the Plant Growth-Promoting Traits of Bacillus Megaterium RmBm31, an Endophytic Bacterium Isolated from Root Nodules of Retama Monosperma. Front. Plant Sci. 2020, 11, 124. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Xu, M.; Guo, J.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Xing, K.; Qin, S. Antifungal Effect of Volatile Organic Compounds Produced by Pseudomonas chlororaphis subsp. Aureofaciens SPS-41 on Oxidative Stress and Mitochondrial Dysfunction of Ceratocystis fimbriata. Pestic. Biochem. Physiol. 2021, 173, 104777. [Google Scholar] [CrossRef] [PubMed]
- Khosroushahi, A.Y.; Valizadeh, M.; Ghasempour, A.; Khosrowshahli, M.; Naghdibadi, H.; Dadpour, M.R.; Omidi, Y. Improved Taxol Production by Combination of Inducing Factors in Suspension Cell Culture of Taxus baccata. Cell Biol. Int. 2006, 30, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Sun, K.; Chen, F.; Yuan, J.; Li, X.; Dai, C. Endophytic Pseudomonas Induces Metabolic Flux Changes That Enhance Medicinal Sesquiterpenoid Accumulation in Atractylodes lancea. Plant Physiol. Biochem. 2018, 130, 473–481. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Observed_Species | Shannon | Simpson | Chao 1 | ACE | Goods_Coverage | PD_Whole_Tree |
|---|---|---|---|---|---|---|---|
| Endophytic bacteria | |||||||
| TR.1 | 442 | 4.432 | 0.827 | 569.317 | 591.599 | 0.988 | 33.877 |
| TR.2 | 577 | 5.22 | 0.893 | 825.061 | 852.577 | 0.981 | 45.092 |
| TR.3 | 535 | 4.749 | 0.805 | 679.222 | 713.189 | 0.985 | 42.331 |
| TL.1 | 28 | 0.157 | 0.029 | 31 | 33.452 | 0.999 | 3.373 |
| TL.2 | 31 | 0.128 | 0.025 | 146.5 | 103.173 | 0.998 | 4.162 |
| TL.3 | 83 | 0.257 | 0.043 | 170.5 | 186.929 | 0.996 | 8.284 |
| TS.1 | 47 | 0.202 | 0.034 | 68.375 | 71.53 | 0.998 | 4.637 |
| TS.2 | 24 | 0.111 | 0.019 | 33 | 40.664 | 0.999 | 2.488 |
| TS.3 | 39 | 0.191 | 0.033 | 47.667 | 52.891 | 0.999 | 4.261 |
| Endophytic fungi | |||||||
| TR.1 | 497 | 3.877 | 0.867 | 559.129 | 570.758 | 0.999 | 328.138 |
| TR.2 | 315 | 2.629 | 0.681 | 336 | 353.881 | 0.999 | 196.471 |
| TR.3 | 702 | 4.183 | 0.866 | 780.705 | 790.063 | 0.998 | 421.522 |
| TL.1 | 761 | 4.437 | 0.9 | 833.02 | 851.161 | 0.998 | 360.4 |
| TL.2 | 1017 | 5.169 | 0.92 | 1328.675 | 1236.798 | 0.996 | 534.092 |
| TL.3 | 457 | 2.712 | 0.686 | 512.152 | 530.888 | 0.999 | 236.541 |
| TS.1 | 937 | 4.622 | 0.871 | 1021.037 | 1035.52 | 0.998 | 450.165 |
| TS.2 | 326 | 3.261 | 0.785 | 368.981 | 373.437 | 0.999 | 194.626 |
| TS.3 | 394 | 3.15 | 0.803 | 445.891 | 448.959 | 0.999 | 210.703 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Li, L.; Chen, Y.; Wang, S.; Xue, L.; Meng, W.; Jiang, J.; Cao, X. Diversity of Endophytic Microbes in Taxus yunnanensis and Their Potential for Plant Growth Promotion and Taxane Accumulation. Microorganisms 2023, 11, 1645. https://doi.org/10.3390/microorganisms11071645
Liu Q, Li L, Chen Y, Wang S, Xue L, Meng W, Jiang J, Cao X. Diversity of Endophytic Microbes in Taxus yunnanensis and Their Potential for Plant Growth Promotion and Taxane Accumulation. Microorganisms. 2023; 11(7):1645. https://doi.org/10.3390/microorganisms11071645
Chicago/Turabian StyleLiu, Qiao, Ludan Li, Yujie Chen, Sai Wang, Lina Xue, Weiying Meng, Jihong Jiang, and Xiaoying Cao. 2023. "Diversity of Endophytic Microbes in Taxus yunnanensis and Their Potential for Plant Growth Promotion and Taxane Accumulation" Microorganisms 11, no. 7: 1645. https://doi.org/10.3390/microorganisms11071645