
Involvement of AoMdr1 in the Regulation of the Fluconazole Resistance, Mycelial Fusion, Conidiation, and Trap Formation of Arthrobotrys oligospora


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Growth Conditions
2.2. Sequence and Cluster Analysis of AoMdr1
2.3. Deletion and Verification of Aomdr1
2.4. Comparison of Mycelial Growth and Cell Nuclei
2.5. Observation of Mycelial Fusion and Lipid Droplet (LD) Accumulation
2.6. Stress Response Assays
2.7. Comparison of Conidial Production, Trap Induction, and Bioassay
2.8. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) Assays
2.9. Metabolomics Profile Analysis
2.10. Statistical Analysis
3. Results
3.1. Sequence and Phylogenetic Analyses of AoMdr1
3.2. AoMdr1 Plays a Vital Role in Hyphal Septa and the Number of Cell Nuclei
3.3. AoMdr1 Regulates LD Accumulation and Hyphal Fusion under Nutrient-Deprived Conditions
3.4. AoMdr1 Afects the Stress Responses of Fluconazole and Chemical Reagents
3.5. AoMdr1 Impairs Sporulation and Spore Morphology
3.6. AoMdr1 Regulates the Number of Traps and Trap Morphology
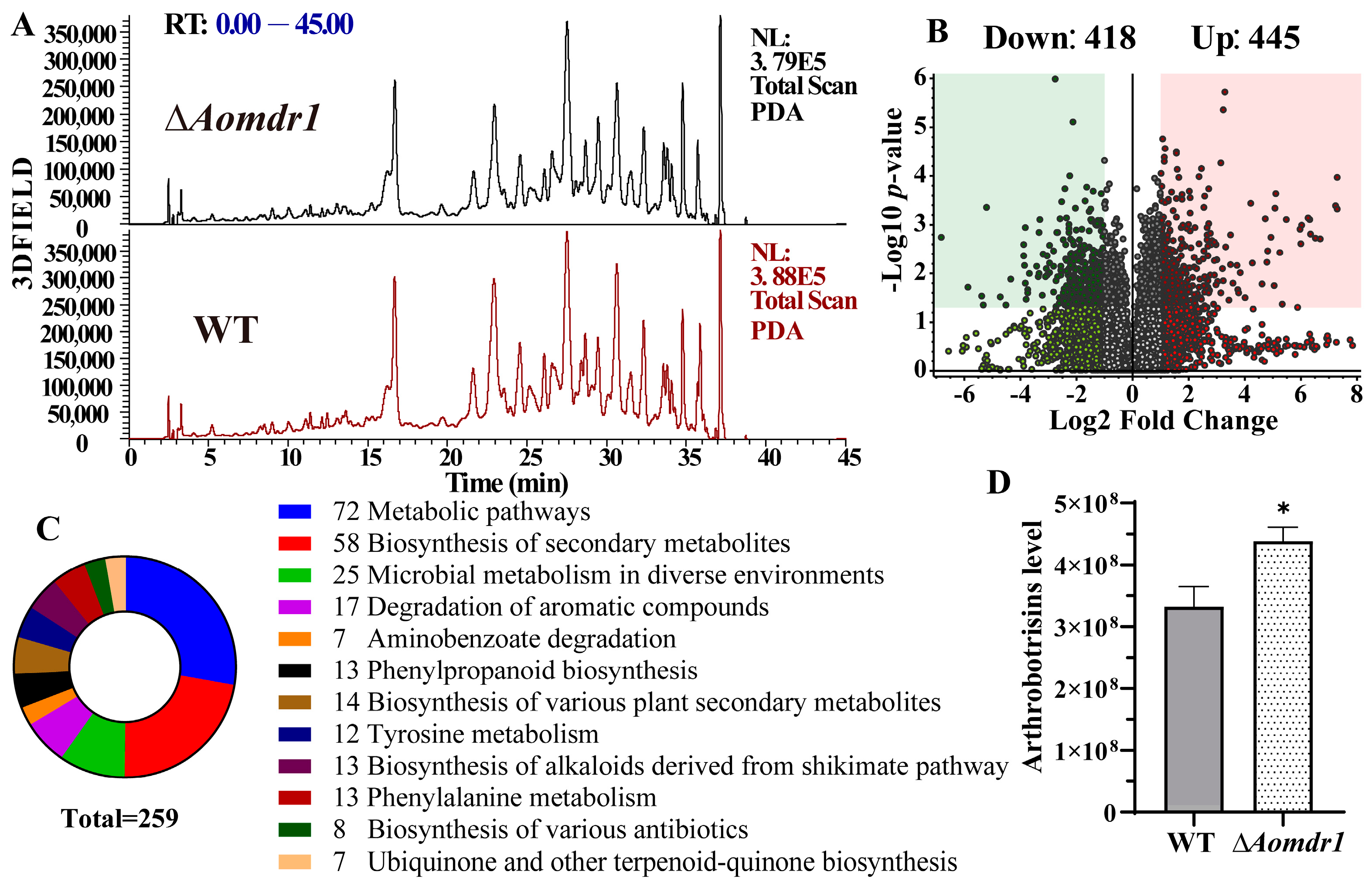
3.7. AoMdr1 Impairs Secondary Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nazarov, P.A. MDR pumps as crossroads of resistance: Antibiotics and bacteriophages. Antibiotics 2022, 11, 734. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Pais, P.; Galocha, M.; Teixeira, M.C. Host-pathogen interactions mediated by MDR transporters in fungi: As pleiotropic as it gets! Genes 2018, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Wegner, M.S.; Gruber, L.; Mattjus, P.; Geisslinger, G.; Grosch, S. The UDP-glucose ceramide glycosyltransferase (UGCG) and the link to multidrug resistance protein 1 (MDR1). BMC Cancer 2018, 18, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.L.; Myers, L.C. Candida albicans Swi/Snf and mediator complexes differentially regulate Mrr1-induced MDR1 expression and fluconazole resistance. Antimicrob. Agents Chemother. 2017, 61, e01344-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.Y.; Cao, Z.R.; Wang, Q.; Wang, Y.C.; Wang, X.J.; Chen, H.B.; Wang, H. MDR1 overexpression combined with ERG11 mutations induce high-level fluconazole resistance in Candida tropicalis clinical isolates. BMC Infect. Dis. 2018, 18, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.L.; Zhai, P.F.; Wang, T.L.; Bromley, M.J.; Zhang, Y.W.; Lu, L. The C2H2 transcription factor SltA contributes to azole resistance by coregulating the expression of the drug target Erg11A and the drug efflux pump Mdr1 in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2021, 65, e01839-20. [Google Scholar] [CrossRef]
- Fattahi, A.; Zaini, F.; Kordbacheh, P.; Rezaie, S.; Safara, M.; Fateh, R.; Farahyar, S.; Kanani, A.; Heidari, M. Evaluation of mRNA expression Levels of cyp51A and mdr1, candidate genes for voriconazole resistance in Aspergillus flavus. Jundishapur. J. Microbiol. 2015, 8, e26990. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.L.; Uhrig, J.; Vu, K.; Singapuri, A.; Dennis, M.; Gelli, A.; Thompson, G.R. Fluconazole susceptibility in Cryptococcus gattii is dependent on the ABC transporter Pdr11. Antimicrob. Agents Chemother. 2016, 60, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.Z.; Xiang, M.C.; Liu, X.Z. Nematode-trapping fungi. Microbiol. Spectr. 2017, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Zhao, Y.; Zhou, J.; Feng, H.H.; Jiang, D.W.; Zhang, K.Q.; Yang, J.K. Trapping devices of nematode-trapping fungi: Formation, evolution, and genomic perspectives. Biol. Rev. Camb. Philos. Soc. 2017, 92, 357–368. [Google Scholar] [CrossRef]
- Zhu, M.C.; Li, X.M.; Zhao, N.; Yang, L.; Zhang, K.Q.; Yang, J.K. Regulatory mechanism of trap formation in the nematode-trapping fungi. J. Fungi 2022, 8, 406. [Google Scholar] [CrossRef]
- Yang, E.C.; Xu, L.L.; Yang, Y.; Zhang, X.Y.; Xiang, M.C.; Wang, C.S.; An, Z.Q.; Liu, X.Z. Origin and evolution of carnivorism in the Ascomycota (fungi). Proc. Natl. Acad. Sci. USA 2012, 109, 10960–10965. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, G.H.; Zou, C.G.; Ji, X.L.; Liu, T.; Zhao, P.J.; Liang, L.M.; Xu, J.P.; An, Z.Q.; Zheng, X.; et al. Bacteria can mobilize nematode-trapping fungi to kill nematodes. Nat. Commun. 2014, 5, 5776. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.K.; Wang, L.; Ji, X.L.; Feng, Y.; Li, X.M.; Zou, C.G.; Xu, J.P.; Ren, Y.; Mi, Q.L.; Wu, J.L.; et al. Genomic and proteomic analyses of the fungus Arthrobotrys oligospora provide insights into nematode-trap formation. PLoS Pathog. 2011, 7, e1002179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.X.; Zhu, Y.M.; Bai, N.; Yang, L.; Xie, M.H.; Yang, J.L.; Zhu, M.C.; Zhang, K.Q.; Yang, J.K. AoATG5 plays pleiotropic roles in vegetative growth, cell nucleus development, conidiation, and virulence in the nematode-trapping fungus Arthrobotrys oligospora. Sci. China Life Sci. 2022, 65, 412–425. [Google Scholar] [CrossRef]
- Chen, Y.L.; Gao, Y.; Zhang, K.Q.; Zou, C.G. Autophagy is required for trap formation in the nematode-trapping fungus Arthrobotrys oligospora. Environ. Microbiol. Rep. 2013, 5, 511–517. [Google Scholar] [CrossRef]
- Li, X.M.; Zhu, M.C.; Liu, Y.K.; Yang, L.; Yang, J.K. Aoatg11 and Aoatg33 are indispensable for mitophagy, and contribute to conidiation, the stress response, and pathogenicity in the nematode-trapping fungus Arthrobotrys oligospora. Microbiol. Res. 2023, 266, 127252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.X.; Zhu, Y.M.; Bai, N.; Xie, M.H.; Zhang, K.Q.; Yang, J.K. Aolatg1 and Aolatg13 regulate autophagy and play different roles in conidiation, trap formation, and pathogenicity in the nematode-trapping fungus Arthrobotrys oligospora. Front. Cell. Infect. Microbiol. 2022, 11, 824407. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Li, D.N.; Bai, N.; Zhu, Y.M.; Yang, J.K. Peroxin Pex14/17 is required for trap formation, and plays pleiotropic roles in mycelial development, stress response, and secondary metabolism in Arthrobotrys oligospora. mSphere 2023, 8, e0001223. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Li, D.N.; Jiang, K.X.; Zhang, K.Q.; Yang, J.K. AoPEX1 and AoPEX6 are required for mycelial growth, conidiation, stress response, fatty acid utilization, and trap formation in Arthrobotrys oligospora. Microbiol. Spectr. 2022, 10, e0027522. [Google Scholar] [CrossRef]
- Bai, N.; Zhang, G.S.; Wang, W.J.; Feng, H.H.; Yang, X.W.; Zheng, Y.Q.; Yang, L.; Xie, M.H.; Zhang, K.Q.; Yang, J.K. Ric8 acts as a regulator of G-protein signalling required for nematode-trapping lifecycle of Arthrobotrys oligospora. Environ. Microbiol. 2022, 24, 1714–1730. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Zhao, Y.N.; Wang, Y.C.; Yang, L.; Li, D.N.; Yang, J.L.; Jiang, K.X.; Zhang, K.Q.; Yang, J.K. Functional analysis of seven regulators of G protein signaling (RGSs) in the nematode-trapping fungus Arthrobotrys oligospora. Virulence 2021, 12, 1825–1840. [Google Scholar] [CrossRef]
- Wang, W.J.; Zhao, Y.N.; Bai, N.; Zhang, K.Q.; Yang, J.K. AMPK is involved in regulating the utilization of carbon sources, conidiation, pathogenicity, and stress response of the nematode-trapping fungus Arthrobotrys oligospora. Microbiol. Spectr. 2022, 10, e0222522. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.A.; Lin, H.C.; Schroeder, F.C.; Hsueh, Y.P. Prey sensing and response in a nematode-trapping fungus is governed by the MAPK pheromone response pathway. Genetics 2021, 217, iyaa008. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.H.; Yang, J.L.; Jiang, K.X.; Bai, N.; Zhu, M.C.; Zhu, Y.M.; Zhang, K.Q.; Yang, J.K. AoBck1 and AoMkk1 are necessary to maintain cell wall integrity, vegetative growth, conidiation, stress resistance, and pathogenicity in the nematode-trapping fungus Arthrobotrys oligospora. Front. Microbiol. 2021, 12, 649582. [Google Scholar] [CrossRef]
- Zhen, Z.Y.; Xing, X.J.; Xie, M.H.; Yang, L.; Yang, X.W.; Zheng, Y.Q.; Chen, Y.L.; Ma, N.; Li, Q.; Zhang, K.Q.; et al. MAP kinase Slt2 orthologs play similar roles in conidiation, trap formation, and pathogenicity in two nematode-trapping fungi. Fungal Genet. Biol. 2018, 116, 42–50. [Google Scholar] [CrossRef]
- Christianson, T.W.; Sikorski, R.S.; Dante, M.; Shero, J.H.; Hieter, P. Multifunctional yeast high-copy-number shuttle vectors. Gene 1992, 110, 119–122. [Google Scholar] [CrossRef]
- Jiang, D.W.; Zhou, J.; Bai, G.Z.; Xing, X.J.; Tang, L.Y.; Yang, X.W.; Li, J.; Zhang, K.Q.; Yang, J.K. Random mutagenesis analysis and identification of a novel C(2)H(2)type transcription factor from the nematode-trapping fungus Arthrobotrys oligospora. Sci. Rep. 2017, 7, 5640. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.X.; Yang, X.W.; Xie, M.H.; Zhang, G.S.; Yang, L.; Bai, N.; Zhao, Y.N.; Li, D.N.; Zhang, K.Q.; Yang, J.K. The Arf-GAP AoGlo3 regulates conidiation, endocytosis, and pathogenicity in the nematode-trapping fungus Arthrobotrys oligospora. Fungal Genet. Biol. 2020, 138, 103352. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvinkova, L.; Weiss, R.L.; Borkovich, K.A.; Dunlap, J.C. A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhang, W.W.; Chen, Y.; Xiang, M.C.; Liu, X.Z. DdaSTE12 is involved in trap formation, ring inflation, conidiation, and vegetative growth in the nematode-trapping fungus Drechslerella dactyloides. Appl. Microbiol. Biotechnol. 2021, 105, 7379–7393. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, M.; Wang, W.; Li, X.; Bai, N.; Xie, M.; Yang, J. AoMae1 regulates hyphal fusion, lipid droplet accumulation, conidiation, and trap formation in Arthrobotrys oligospora. J. Fungi 2023, 9, 496. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.W.; Ma, N.; Yang, L.; Zheng, Y.Q.; Zhen, Z.Y.; Li, Q.; Xie, M.H.; Li, J.; Zhang, K.Q.; Yang, J.K. Two Rab GTPases play different roles in conidiation, trap formation, stress resistance, and virulence in the nematode-trapping fungus Arthrobotrys oligospora. Appl. Microbiol. Biotechnol. 2018, 102, 4601–4613. [Google Scholar] [CrossRef]
- Jiang, K.X.; Liu, Q.Q.; Bai, N.; Zhu, M.C.; Zhang, K.Q.; Yang, J.K. AoSsk1, a response regulator required for mycelial growth and development, stress responses, trap formation, and the secondary metabolism in Arthrobotrys oligospora. J. Fungi 2022, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.M.; Zhou, D.X.; Bai, N.; Liu, Q.Q.; Zhao, N.; Yang, J.K. SNARE protein AoSec22 orchestrates mycelial growth, vacuole assembly, trap formation, stress response, and secondary metabolism in Arthrobotrys oligospora. J. Fungi 2023, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.S.; Zheng, Y.Q.; Ma, Y.X.; Yang, L.; Xie, M.H.; Zhou, D.X.; Niu, X.M.; Zhang, K.Q.; Yang, J.K. The velvet proteins VosA and VelB play different roles in conidiation, trap formation, and pathogenicity in the nematode-trapping fungus Arthrobotrys oligospora. Front Microbiol. 2019, 10, 1917. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tong, S.M.; Qiu, L.; Ying, S.H.; Feng, M.G. Two histidine kinases can sense different stress cues for activation of the MAPK Hog1 in a fungal insect pathogen. Environ. Microbiol. 2017, 19, 4091–4102. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.K.; Sun, H.H.; Ying, S.H.; Feng, M.G. Characterization of the Hog1 MAPK pathway in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2017, 19, 1808–1821. [Google Scholar] [CrossRef]
- Yang, J.L.; Wang, W.J.; Liu, Y.K.; Xie, M.H.; Yang, J.K. The MADS-box transcription factor AoRlmA is involved in the regulation of mycelium development, conidiation, cell-wall integrity, stress response, and trap formation of Arthrobotrys oligospora. Microbiol. Res. 2023, 268, 127299. [Google Scholar] [CrossRef]
- Yang, L.; Li, X.M.; Xie, M.H.; Bai, N.; Yang, J.L.; Jiang, K.X.; Zhang, K.Q.; Yang, J.K. Pleiotropic roles of Ras GTPases in the nematode-trapping fungus Arthrobotrys oligospora identified through multi-omics analyses. Iscience 2021, 24, 102820. [Google Scholar] [CrossRef]
- Xie, M.H.; Ma, N.; Bai, N.; Yang, L.; Yang, X.W.; Zhang, K.Q.; Yang, J.K. PKC-SWI6 signaling regulates asexual development, cell wall integrity, stress response, and lifestyle transition in the nematode-trapping fungus Arthrobotrys oligospora. Sci. China Life Sci. 2022, 65, 2455–2471. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, Q.F.; Li, S.H.; Yan, J.X.; Wu, L.; Cheng, Q.Y.; He, Z.Q.; Yue, X.T.; Zhang, K.Q.; Zhang, L.L.; et al. The multifaceted Gene 275 embedded in the PKS-PTS gene cluster was involved in the regulation of arthrobotrisin biosynthesis, TCA cycle, and septa formation in nematode-trapping fungus Arthrobotrys oligospora. J. Fungi 2022, 8, 1261. [Google Scholar] [CrossRef]
- Xie, M.H.; Ma, N.; Bai, N.; Zhu, M.C.; Zhang, K.Q.; Yang, J.K. Phospholipase C (AoPLC2) regulates mycelial development, trap morphogenesis, and pathogenicity of the nematode-trapping fungus Arthrobotrys oligospora. J Appl. Microbiol. 2022, 132, 2144–2156. [Google Scholar] [CrossRef]
- Schubert, S.; Barker, K.S.; Znaidi, S.; Schneider, S.; Dierolf, F.; Dunkel, N.; Aid, M.; Boucher, G.; Rogers, P.D.; Raymond, M.; et al. Regulation of efflux pump expression and drug resistance by the transcription factors Mrr1, Upc2, and Cap1 in Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 2212–2223. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, K.; Guo, X.H.; Jiang, J.; Qian, L.; Xu, J.Q.; Che, Z.P.; Huang, X.B.; Liu, S.M. Transcriptomic profiling of Fusarium pseudograminearum in response to carbendazim, pyraclostrobin, tebuconazole, and phenamacril. J. Fungi 2023, 9, 334. [Google Scholar] [CrossRef] [PubMed]
- Song, T.T.; Zhao, J.; Ying, S.H.; Feng, M.G. Differential contributions of five ABC Transporters to mutidrug resistance, antioxidion and virulence of Beauveria bassiana, an entomopathogenic fungus. PLoS ONE 2013, 8, e62179. [Google Scholar] [CrossRef] [Green Version]
- Jonkers, W.; Fischer, M.S.; Do, H.P.; Starr, T.L.; Glass, N.L. Chemotropism and cell fusion in Neurospora crassa Relies on the formation of distinct protein complexes by HAM-5 and a novel protein HAM-14. Genetics 2016, 203, 319–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, S.; Schumann, M.R.; Fleissner, A. Cell fusion in Neurospora crassa. Curr. Opin. Microbiol. 2015, 28, 53–59. [Google Scholar] [CrossRef]
- Nordbringhertz, B.; Friman, E.; Veenhuis, M. Hyphal fusion during initial stages of trap formation in Arthrobotrys oligospora. Antonie Van Leeuwenhoek 1989, 55, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, N.; Xie, M.; Liu, Q.; Wang, W.; Liu, Y.; Yang, J. AoSte12 is required for mycelial development, conidiation, trap morphogenesis, and secondary metabolism by regulating hyphal fusion in nematode-trapping fungus Arthrobotrys oligospora. Microbiol. Spectr. 2023, 11, e0395722. [Google Scholar] [CrossRef]
- Suchodolski, J.; Krasowska, A. Fructose induces fluconazole resistance in Candida albicans through activation of Mdr1 and Cdr1 transporters. Int. J. Mol. Sci. 2021, 22, 2127. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Coste, A.T.; Bachmann, D.; Sanglard, D.; Lamoth, F. Deciphering the Mrr1/Mdr1 pathway in azole resistance of Candida auris. Antimicrob. Agents Chemother. 2022, 66, e0006722. [Google Scholar] [CrossRef]
- Basso, L.R.; Gast, C.E.; Bruzual, I.; Wong, B. Identification and properties of plasma membrane azole efflux pumps from the pathogenic fungi Cryptococcus gattii and Cryptococcus neoformans. J. Antimicrob. Chemother. 2015, 70, 1396–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef]
- Wei, L.X.; Zhang, H.X.; Tan, J.L.; Chu, Y.S.; Li, N.; Xue, H.X.; Wang, Y.L.; Niu, X.M.; Zhang, Y.; Zhang, K.Q. Arthrobotrisins A-C, oligosporons from the nematode-trapping fungus Arthrobotrys oligospora. J. Nat. Prod. 2011, 74, 1526–1530. [Google Scholar] [CrossRef]
- Yu, X.; Hu, X.; Pop, M.; Wernet, N.; Kirschhöfer, F.; Brenner-Weiß, G.; Keller, J.; Bunzel, M.; Fischer, R. Fatal attraction of Caenorhabditis elegans to predatory fungi through 6-methyl-salicylic acid. Nat. Commun. 2021, 12, 5462. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, X.; Zhu, M.; Bai, N.; Wang, W.; Yang, J. Involvement of AoMdr1 in the Regulation of the Fluconazole Resistance, Mycelial Fusion, Conidiation, and Trap Formation of Arthrobotrys oligospora. Microorganisms 2023, 11, 1612. https://doi.org/10.3390/microorganisms11061612
Liu Y, Yang X, Zhu M, Bai N, Wang W, Yang J. Involvement of AoMdr1 in the Regulation of the Fluconazole Resistance, Mycelial Fusion, Conidiation, and Trap Formation of Arthrobotrys oligospora. Microorganisms. 2023; 11(6):1612. https://doi.org/10.3390/microorganisms11061612
Chicago/Turabian StyleLiu, Yankun, Xuewei Yang, Meichen Zhu, Na Bai, Wenjie Wang, and Jinkui Yang. 2023. "Involvement of AoMdr1 in the Regulation of the Fluconazole Resistance, Mycelial Fusion, Conidiation, and Trap Formation of Arthrobotrys oligospora" Microorganisms 11, no. 6: 1612. https://doi.org/10.3390/microorganisms11061612