

Production of Polyhydroxybutyrate by Genetically Modified Pseudomonas sp. phDV1: A Comparative Study of Utilizing Wine Industry Waste as a Carbon Source
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Medium, and Oligonucleotides
2.2. Construction of the ΔphaZ and ΔphaR Strains
2.3. Cultivation of Pseudomonas Strains
2.4. Analytical Methods
2.5. Analysis of the Polyphenolic Profile of the Culture
2.6. Nile Red Staining
2.7. Quantification of PHB with HPLC
2.8. Isolation of PHB Granules
2.9. NMR Spectroscopy
2.10. Data Analysis
3. Results and Discussion
3.1. Construction of ΔPhaZ and ΔPhaR Mutants of Pseudomonas sp. phDV1
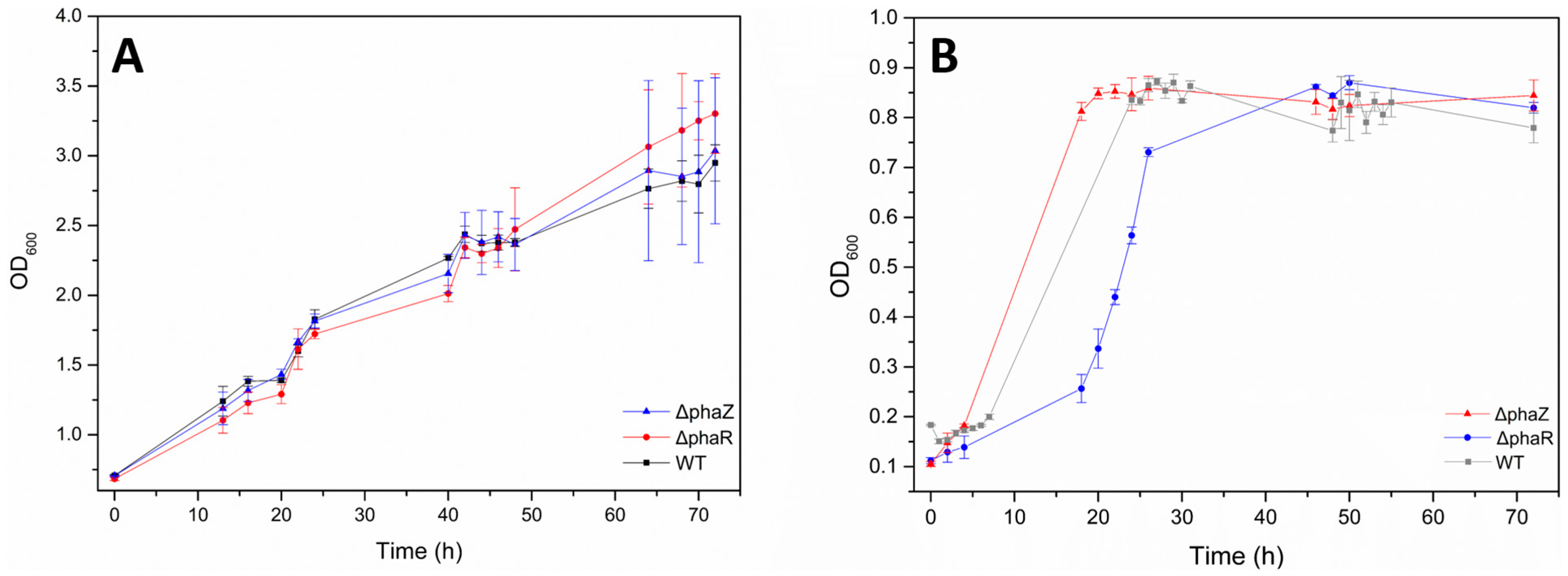
3.2. Pseudomonas sp. phDV1 Cell Growth in Grape Pomace Extract
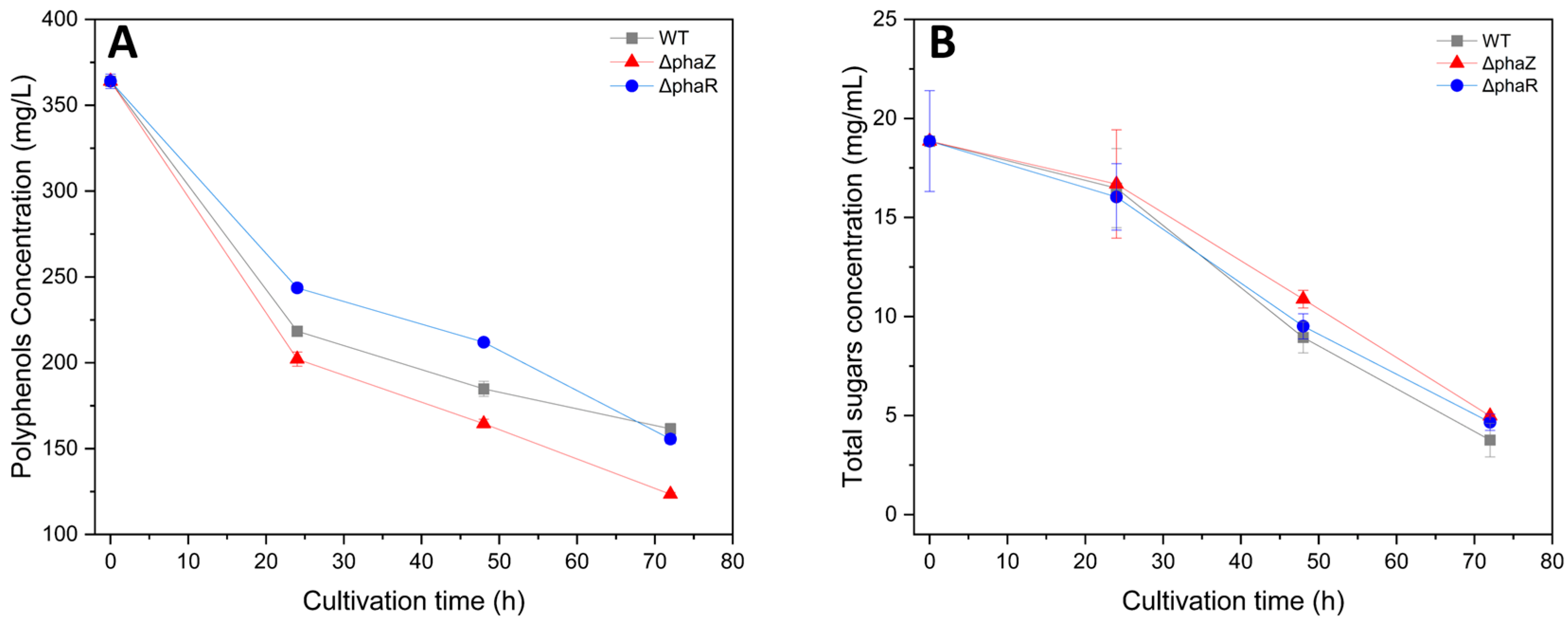
3.3. Biodegradation of Sugars and Polyphenols
3.4. PHB Production from Grape
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandez-Castillo, R.; Rodriguez-Valera, F.; Gonzalez-Ramos, J.; Ruiz-Berraquero, F. Accumulation of Poly (Beta-Hydroxybutyrate) by Halobacteria. Appl. Environ. Microbiol. 1986, 51, 214–216. [Google Scholar] [CrossRef] [Green Version]
- Steinbüchel, A.; Füchtenbusch, B. Bacterial and Other Biological Systems for Polyester Production. Trends Biotechnol. 1998, 16, 419–427. [Google Scholar] [CrossRef]
- Anderson, A.J.; Dawes, E.A. Occurrence, Metabolism, Metabolic Role, and Industrial Uses of Bacterial Polyhydroxyalkanoates. Microbiol. Rev. 1990, 54, 450–472. [Google Scholar] [CrossRef]
- de Smet, M.J.; Eggink, G.; Witholt, B.; Kingma, J.; Wynberg, H. Characterization of Intracellular Inclusions Formed by Pseudomonas Oleovorans during Growth on Octane. J. Bacteriol. 1983, 154, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Griebel, R.; Smith, Z.; Merrick, J.M. Metabolism of Poly-Beta-Hydroxybutyrate. I. Purification, Composition, and Properties of Native Poly-Beta-Hydroxybutyrate Granules from Bacillus Megaterium. Biochemistry 1968, 7, 3676–3681. [Google Scholar] [CrossRef]
- Jendrossek, D.; Pfeiffer, D. New Insights in the Formation of Polyhydroxyalkanoate Granules (Carbonosomes) and Novel Functions of Poly(3-Hydroxybutyrate). Environ. Microbiol. 2014, 16, 2357–2373. [Google Scholar] [CrossRef]
- Maehara, A.; Taguchi, S.; Nishiyama, T.; Yamane, T.; Doi, Y. A Repressor Protein, PhaR, Regulates Polyhydroxyalkanoate (PHA) Synthesis via Its Direct Interaction with PHA. J. Bacteriol. 2002, 184, 3992–4002. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Yamashita, K.; Wakuda, A.; Ichimura, K.; Maehara, A.; Maeda, M.; Taguchi, S. Autoregulator Protein PhaR for Biosynthesis of Polyhydroxybutyrate [P(3HB)] Possibly Has Two Separate Domains That Bind to the Target DNA and P(3HB): Functional Mapping of Amino Acid Residues Responsible for DNA Binding. J. Bacteriol. 2007, 189, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Valsamidis, G.; Mathioudaki, E.; Tsiotis, G. Complete Genome Sequence of Pseudomonas Sp. Strain PhDV1, an Isolate Capable of Efficient Degradation of Aromatic Hydrocarbons. Microbiol. Resour. Announc. 2019, 8, e01171-18. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete Genome Sequence of Pseudomonas aeruginosa PA01, an Opportunistic Pathogen. Nature 2000, 406, 959–964. [Google Scholar]
- Klinke, S.; de Roo, G.; Witholt, B.; Kessler, B. Role of PhaD in Accumulation of Medium-Chain-Length Poly(3-Hydroxyalkanoates) in Pseudomonas Oleovorans. Appl. Environ. Microbiol. 2000, 66, 3705–3710. [Google Scholar] [CrossRef] [Green Version]
- Prieto, M.A.; de Eugenio, L.I.; Galàn, B.; Luengo, J.M.; Witholt, B. Synthesis and Degradation of Polyhydroxyalkanoates. In Pseudomonas; Ramos, J.-L., Filloux, A., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 397–428. ISBN 978-1-4020-6096-0. [Google Scholar]
- Prieto, A.; Escapa, I.F.; Martínez, V.; Dinjaski, N.; Herencias, C.; de la Peña, F.; Tarazona, N.; Revelles, O. A Holistic View of Polyhydroxyalkanoate Metabolism in Pseudomonas Putida. Environ. Microbiol. 2016, 18, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-Q.; Jiang, X.-R. Engineering Microorganisms for Improving Polyhydroxyalkanoate Biosynthesis. Curr. Opin. Biotechnol. 2018, 53, 20–25. [Google Scholar] [CrossRef]
- Huisman, G.W.; Wonink, E.; Meima, R.; Kazemier, B.; Terpstra, P.; Witholt, B. Metabolism of Poly(3-Hydroxyalkanoates) (PHAs) by Pseudomonas Oleovorans. Identification and Sequences of Genes and Function of the Encoded Proteins in the Synthesis and Degradation of PHA. J. Biol. Chem. 1991, 266, 2191–2198. [Google Scholar] [CrossRef]
- Cai, L.; Yuan, M.-Q.; Liu, F.; Jian, J.; Chen, G.-Q. Enhanced Production of Medium-Chain-Length Polyhydroxyalkanoates (PHA) by PHA Depolymerase Knockout Mutant of Pseudomonas Putida KT2442. Bioresour. Technol. 2009, 100, 2265–2270. [Google Scholar] [CrossRef]
- Salvachúa, D.; Rydzak, T.; Auwae, R.; de Capite, A.; Black, B.A.; Bouvier, J.T.; Cleveland, N.S.; Elmore, J.R.; Furches, A.; Huenemann, J.D.; et al. Metabolic Engineering of Pseudomonas putida for Increased Polyhydroxyalkanoate Production from Lignin. Microb. Biotechnol. 2020, 13, 813. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S.; Srivastava, A.K. Recent Advances in Microbial Polyhydroxyalkanoates. Process Biochem. 2005, 40, 607–619. [Google Scholar] [CrossRef]
- Chen, G.Q. A Microbial Polyhydroxyalkanoates (PHA) Based Bio- and Materials Industry. Chem. Soc. Rev. 2009, 38, 2434–2446. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Kim, J.H.; Kim, M.S.; Kim, J.; Hong, J.W.; Hong, Y.G.; Kim, H.J.; Jeon, J.M.; Kim, S.H.; Ahn, J.; et al. Production of (3-Hydroxybutyrate-Co-3-Hydroxyhexanoate) Copolymer from Coffee Waste Oil Using Engineered Ralstonia Eutropha. Bioprocess Biosyst. Eng. 2018, 41, 229–235. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.K.; Ghodake, G.S.; Shin, H.S.; Saratale, G.D.; Park, Y.; Lee, H.S.; Bharagava, R.N.; Kim, D.S. Utilization of Noxious Weed Water Hyacinth Biomass as a Potential Feedstock for Biopolymers Production: A Novel Approach. Polymers 2020, 12, 1704. [Google Scholar] [CrossRef]
- Amadu, A.A.; Qiu, S.; Ge, S.; Addico, G.N.D.; Ameka, G.K.; Yu, Z.; Xia, W.; Abbew, A.W.; Shao, D.; Champagne, P.; et al. A Review of Biopolymer (Poly-β-Hydroxybutyrate) Synthesis in Microbes Cultivated on Wastewater. Sci. Total Environ. 2021, 756, 143729. [Google Scholar] [CrossRef]
- Jung, H.J.; Kim, S.H.; Cho, D.H.; Kim, B.C.; Bhatia, S.K.; Lee, J.; Jeon, J.M.; Yoon, J.J.; Yang, Y.H. Finding of Novel Galactose Utilizing Halomonas Sp. YK44 for Polyhydroxybutyrate (PHB) Production. Polymers 2022, 14, 5407. [Google Scholar] [CrossRef]
- Andler, R.; Pino, V.; Moya, F.; Soto, E.; Valdés, C.; Andreeßen, C. Synthesis of Poly-3-Hydroxybutyrate (PHB) by Bacillus cereus Using Grape Residues as Sole Carbon Source. Int. J. Biobased Plast. 2021, 3, 98–111. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, S.G.; Cho, D.H.; Bhatia, S.K.; Gurav, R.; Yang, S.Y.; Yang, J.; Jeon, J.M.; Yoon, J.J.; Choi, K.Y.; et al. Finding of Novel Lactate Utilizing Bacillus Sp. YHY22 and Its Evaluation for Polyhydroxybutyrate (PHB) Production. Int. J. Biol. Macromol. 2022, 201, 653–661. [Google Scholar] [CrossRef]
- Mozejko-Ciesielska, J.; Szacherska, K.; Marciniak, P. Pseudomonas Species as Producers of Eco-Friendly Polyhydroxyalkanoates. J. Polym. Environ. 2019, 27, 1151–1166. [Google Scholar] [CrossRef] [Green Version]
- Kourmentza, C.; Ntaikou, I.; Lyberatos, G.; Kornaros, M. Polyhydroxyalkanoates from Pseudomonas Sp. Using Synthetic and Olive Mill Wastewater under Limiting Conditions. Int. J. Biol. Macromol. 2015, 74, 202–210. [Google Scholar] [CrossRef]
- Poblete-Castro, I.; Rodriguez, A.L.; Lam, C.M.C.; Kessler, W. Improved Production of Medium-Chain-Length Polyhydroxyalkanoates in Glucose-Based Fed-Batch Cultivations of Metabolically Engineered Pseudomonas Putida Strains. J. Microbiol. Biotechnol. 2014, 24, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Lyratzakis, A.; Valsamidis, G.; Kanavaki, I.; Nikolaki, A.; Rupprecht, F.; Langer, J.D.; Tsiotis, G. Proteomic Characterization of the Pseudomonas Sp. Strain PhDV1 Response to Monocyclic Aromatic Compounds. Proteomics 2020, 21, e2000003. [Google Scholar] [CrossRef]
- Kanavaki, I.; Drakonaki, A.; Geladas, E.D.; Spyros, A.; Xie, H.; Tsiotis, G. Polyhydroxyalkanoate (PHA) Production in Pseudomonas Sp. PhDV1 Strain Grown on Phenol as Carbon Sources. Microorganisms 2021, 9, 1636. [Google Scholar] [CrossRef]
- Kourilova, X.; Pernicova, I.; Vidlakova, M.; Krejcirik, R.; Mrazova, K.; Hrubanova, K.; Krzyzanek, V.; Nebesarova, J.; Obruca, S. Biotechnological Conversion of Grape Pomace to Poly(3-Hydroxybutyrate) by Moderately Thermophilic Bacterium Tepidimonas taiwanensis. Bioengineering 2021, 8, 141. [Google Scholar] [CrossRef]
- Eleutheria, N.; Maria, I.; Vasiliki, T.; Alexandros, E.; Alexandros, A.; Vasileios, D. Energy Recovery and Treatment of Winery Wastes by a Compact Anaerobic Digester. Waste Biomass Valorization 2016, 7, 799–805. [Google Scholar] [CrossRef]
- Dwyer, K.; Hosseinian, F.; Rod, M. The Market Potential of Grape Waste Alternatives. J. Food Res. 2014, 3, 91. [Google Scholar] [CrossRef]
- Ntaikou, I.; Kourmentza, C.; Koutrouli, E.C.; Stamatelatou, K.; Zampraka, A.; Kornaros, M.; Lyberatos, G. Exploitation of Olive Oil Mill Wastewater for Combined Biohydrogen and Biopolymers Production. Bioresour. Technol. 2009, 100, 3724–3730. [Google Scholar] [CrossRef]
- Gumel, A.M.; Annuar, M.S.M.; Heidelberg, T. Biosynthesis and Characterization of Polyhydroxyalkanoates Copolymers Produced by Pseudomonas putida Bet001 Isolated from Palm Oil Mill Effluent. PLoS ONE 2012, 7, e45214. [Google Scholar] [CrossRef] [Green Version]
- Polymenakou, P.N.; Stephanou, E.G. Effect of Temperature and Additional Carbon Sources on Phenol Degradation by an Indigenous Soil Pseudomonad. Biodegradation 2005, 16, 403–413. [Google Scholar] [CrossRef]
- Tsirogianni, I.; Aivaliotis, M.; Karas, M.; Tsiotis, G. Mass Spectrometric Mapping of the Enzymes Involved in the Phenol Degradation of an Indigenous Soil Pseudomonad. Biochim. Biophys. Acta 2004, 1700, 117–123. [Google Scholar] [CrossRef]
- Shulman, M.J.; Hallick, L.M.; Echols, H.; Signer, E.R. Properties of Recombination-Deficient Mutants of Bacteriophage Lambda. J. Mol. Biol. 1970, 52, 501–520. [Google Scholar] [CrossRef]
- Lesic, B.; Rahme, L.G. Use of the Lambda Red Recombinase System to Rapidly Generate Mutants in Pseudomonas aeruginosa. BMC Mol. Biol. 2008, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Kohlstaedt, M.; Buschmann, S.; Xie, H.; Resemann, A.; Warkentin, E.; Langer, J.D.; Michel, H. Identification and Characterization of the Novel Subunit CcoM in the Cbb₃-Cytochrome c Oxidase from Pseudomonas Stutzeri ZoBell. mBio 2016, 7, e01921-15. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-Min Method for Preparation of Highly Electrocompetent Pseudomonas aeruginosa Cells: Application for DNA Fragment Transfer between Chromosomes and Plasmid Transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Belenioti, M.; Mathioudaki, E.; Spyridaki, E.; Ghanotakis, D.; Chaniotakis, N. Biodegradation of Phenolic Compounds from Grape Pomace of Vitis vinifera Asyrtiko by Chlamydomonas reinhardtii. J. Chem. Technol. Biotechnol. 2023; Early View. [Google Scholar] [CrossRef]
- Gröger, W.K.L. Determination of Sugars in Biological Media with Thymol in Sulphuric Acid. Clin. Chim. Acta 1961, 6, 866–873. [Google Scholar] [CrossRef]
- Schulze, C.; Wetzel, M.; Reinhardt, J.; Schmidt, M.; Felten, L.; Mundt, S. Screening of Microalgae for Primary Metabolites Including β-Glucans and the Influence of Nitrate Starvation and Irradiance on β-Glucan Production. J. Appl. Phycol. 2016, 28, 2719–2725. [Google Scholar] [CrossRef]
- Follonier, S. Pilot-Scale Production of Functionalized Mcl-PHA from Grape Pomace Supplemented with Fatty Acids. Chem. Biochem. Eng. Q. 2015, 29, 113–121. [Google Scholar] [CrossRef]
- Zhu, M.; Dai, X. Stringent Response Ensures the Timely Adaptation of Bacterial Growth to Nutrient Downshift. Nat. Commun. 2023, 14, 467. [Google Scholar] [CrossRef]
- Jin, Q.; O’Hair, J.; Stewart, A.C.; O’Keefe, S.F.; Neilson, A.P.; Kim, Y.T.; McGuire, M.; Lee, A.; Wilder, G.; Huang, H. Compositional Characterization of Different Industrial White and Red Grape Pomaces in Virginia and the Potential Valorization of the Major Components. Foods 2019, 8, 667. [Google Scholar] [CrossRef] [Green Version]
- Filippi, K.; Georgaka, N.; Alexandri, M.; Papapostolou, H.; Koutinas, A. Valorisation of Grape Stalks and Pomace for the Production of Bio-Based Succinic Acid by Actinobacillus succinogenes. Ind. Crops Prod. 2021, 168, 113578. [Google Scholar] [CrossRef]
- Tsirogianni, E.; Aivaliotis, M.; Papasotiriou, D.G.; Karas, M.; Tsiotis, G. Identification of Inducible Protein Complexes in the Phenol Degrader Pseudomonas sp. Strain PhDV1 by Blue Native Gel Electrophoresis and Mass Spectrometry. Amino Acids 2006, 30, 63–72. [Google Scholar] [CrossRef]
- Nikolaki, A.; Papadioti, A.; Arvaniti, K.; Kassotaki, E.; Langer, J.D.; Tsiotis, G. The Membrane Complexome of a New Pseudomonas Strain during Growth on Lysogeny Broth Medium and Medium Containing Glucose or Phenol. EuPA Open Proteom. 2014, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Almajano, M.P.; Carbó, R.; Jiménez, J.A.L.; Gordon, M.H. Antioxidant and Antimicrobial Activities of Tea Infusions. Food Chem. 2008, 108, 55–63. [Google Scholar] [CrossRef]
- Vaquero, M.J.R.; Alberto, M.R.; de Nadra, M.C.M. Antibacterial Effect of Phenolic Compounds from Different Wines. Food Control 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Caponio, G.R.; Noviello, M.; Calabrese, F.M.; Gambacorta, G.; Giannelli, G.; De Angelis, M. Effects of Grape Pomace Polyphenols and In Vitro Gastrointestinal Digestion on Antimicrobial Activity: Recovery of Bioactive Compounds. Antioxidants 2022, 11, 567. [Google Scholar] [CrossRef]
- Hervert-Hernández, D.; Pintado, C.; Rotger, R.; Goñi, I. Stimulatory Role of Grape Pomace Polyphenols on Lactobacillus acidophilus Growth. Int. J. Food Microbiol. 2009, 136, 119–122. [Google Scholar] [CrossRef]
- Papazi, A.; Kotzabasis, K. Bioenergetic Strategy of Microalgae for the Biodegradation of Phenolic Compounds—Exogenously Supplied Energy and Carbon Sources Adjust the Level of Biodegradation. J. Biotechnol. 2007, 129, 706–716. [Google Scholar] [CrossRef]
- Papazi, A.; Assimakopoulos, K.; Kotzabasis, K. Bioenergetic Strategy for the Biodegradation of P-Cresol by the Unicellular Green Alga Scenedesmus obliquus. PLoS ONE 2012, 7, e51852. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drakonaki, A.; Mathioudaki, E.; Geladas, E.D.; Konsolaki, E.; Vitsaxakis, N.; Chaniotakis, N.; Xie, H.; Tsiotis, G. Production of Polyhydroxybutyrate by Genetically Modified Pseudomonas sp. phDV1: A Comparative Study of Utilizing Wine Industry Waste as a Carbon Source. Microorganisms 2023, 11, 1592. https://doi.org/10.3390/microorganisms11061592
Drakonaki A, Mathioudaki E, Geladas ED, Konsolaki E, Vitsaxakis N, Chaniotakis N, Xie H, Tsiotis G. Production of Polyhydroxybutyrate by Genetically Modified Pseudomonas sp. phDV1: A Comparative Study of Utilizing Wine Industry Waste as a Carbon Source. Microorganisms. 2023; 11(6):1592. https://doi.org/10.3390/microorganisms11061592
Chicago/Turabian StyleDrakonaki, Athina, Eirini Mathioudaki, Ermis Dionysios Geladas, Eleni Konsolaki, Nikolaos Vitsaxakis, Nikos Chaniotakis, Hao Xie, and Georgios Tsiotis. 2023. "Production of Polyhydroxybutyrate by Genetically Modified Pseudomonas sp. phDV1: A Comparative Study of Utilizing Wine Industry Waste as a Carbon Source" Microorganisms 11, no. 6: 1592. https://doi.org/10.3390/microorganisms11061592