
Molecular Investigation Confirms Myotis Genus Bats as Common Hosts of Polychromophilus in Brazil

 , ,
, ,  ,
,  ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
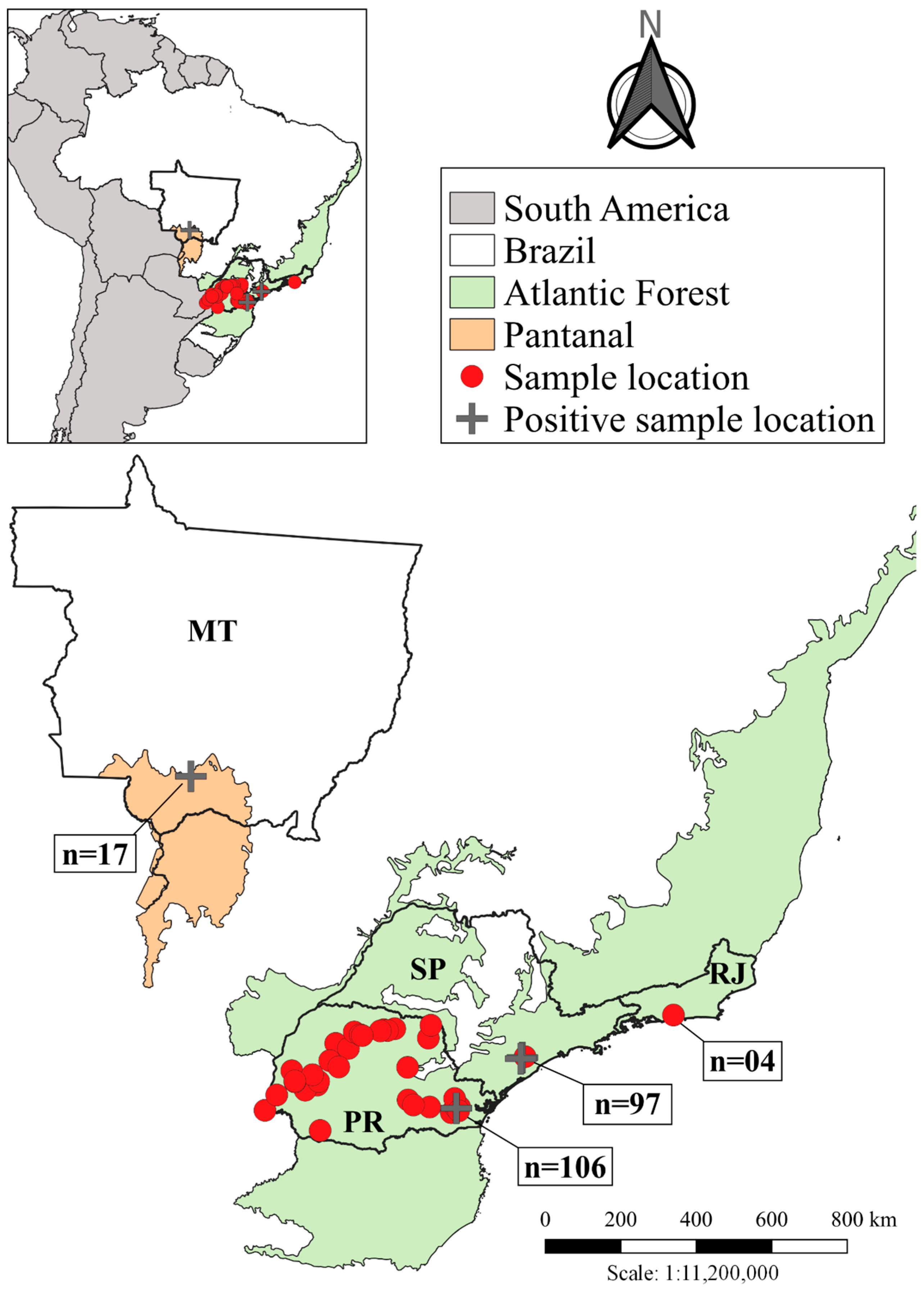
2.1. Sampling
2.1.1. Paraná/PR
2.1.2. Mato Grosso/MT and Rio de Janeiro/RJ
2.1.3. São Paulo/SP
2.2. Ethics Statement
2.3. Optical Microscopy Diagnosis
2.4. DNA Extraction
2.5. Molecular Detection of Haemosporidian Parasites
2.6. Phylogenetic Analysis
2.7. Host Species Identification
3. Results
3.1. Host Species Identification
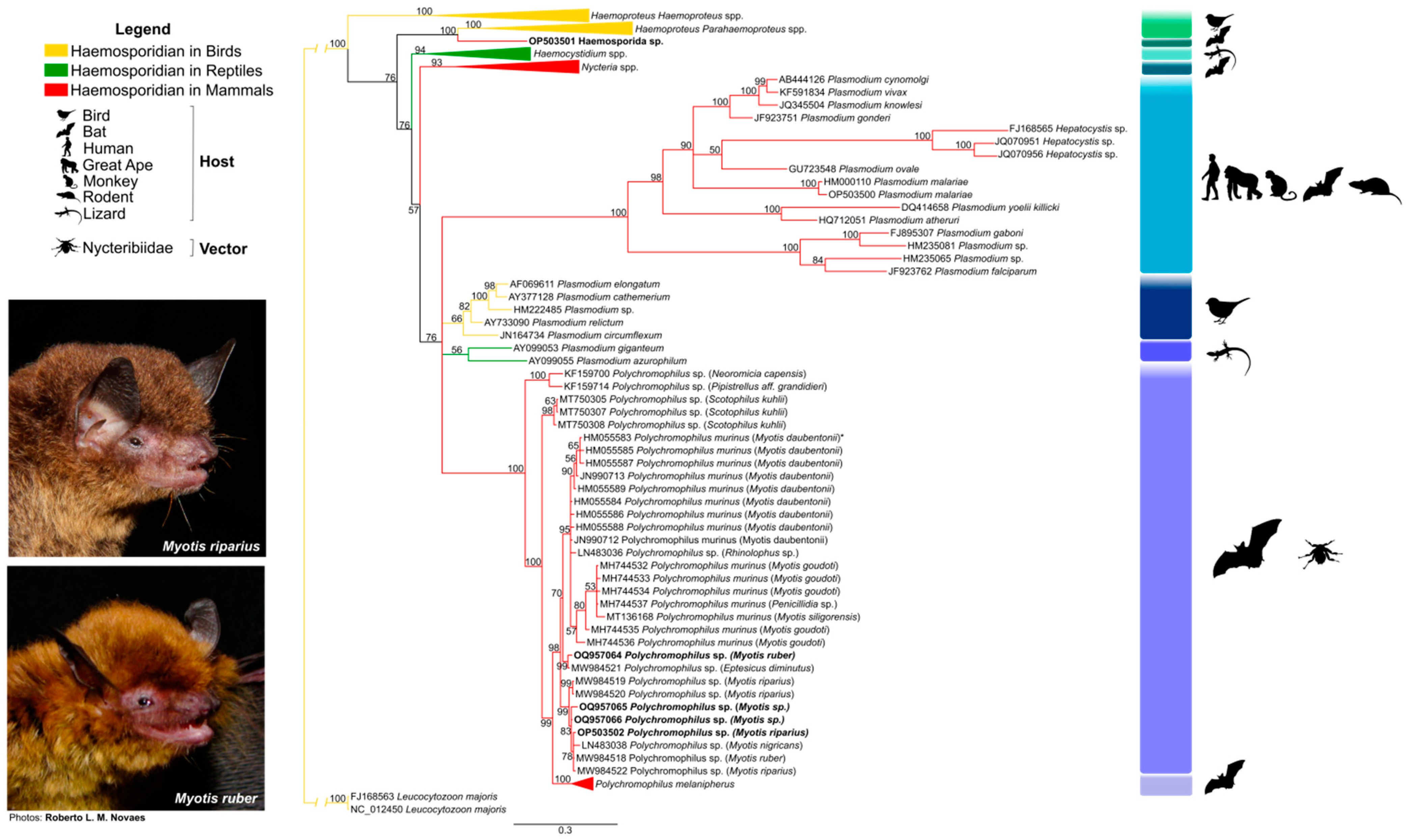
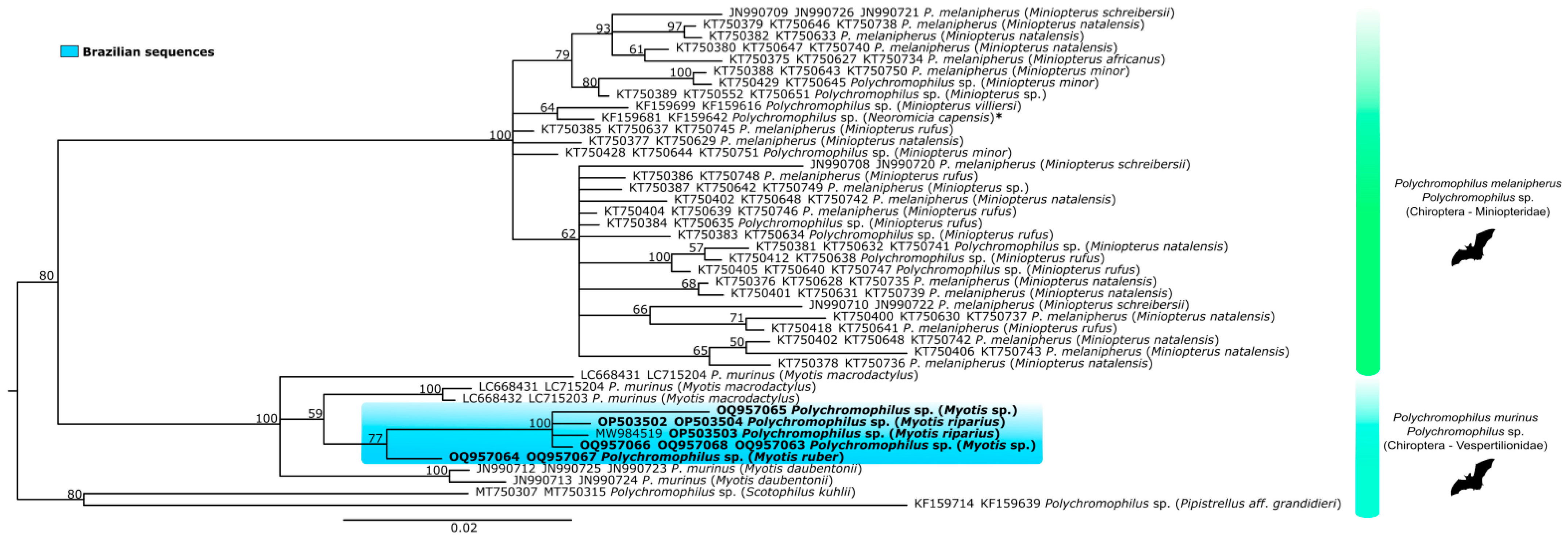
3.2. Haemosporidian Parasites and Phylogeny

{kind=link}
{kind=link}
{kind=link}
| Host Species | Locality | Year of Collection | Bats Examined | Bats Infected with Haemosporidians (%) |
|---|---|---|---|---|
| Vespertilionidae Gray, 1821 | ||||
| Myotis nigricans | Rio Bom, Toledo-PR | 2019–2020 | 2 | 0 |
| Myotis riparius | São José dos Pinhais, Curitiba-PR | 2019–2020 | 2 | 2 (100%) |
| Eptesicus furinalis | Assis Chateaubriand, Curitiba, Foz do Iguaçu, Maringá, Ramilândia-PR | 2019–2020 | 12 | 0 |
| Eptesicus sp. | Curitiba-PR | 2019–2020 | 2 | 0 |
| Myotis sp. | Curitiba-PR | 2019–2020 | 1 | 0 |
| Myotis cf. nigricans | Poconé-MT | 2019 | 4 | 0 |
| Eptesicus sp. | Miracatu-SP | 2018 | 1 | 0 |
| Myotis nigricans | Tapiraí, Miracatu-SP | 2018–2019 | 8 | 0 |
| Myotis riparius | Miracatu-SP | 2019 | 1 | 0 |
| Myotis ruber | Tapiraí-SP | 2019 | 1 | 1 (100%) |
| Myotis sp. | Miracatu-SP | 2019 | 4 | 2 (50%) |
| Molossidae Gervais, 1856 | ||||
| Molossus sp. | Curitiba, Araruna-PR | 2019–2020 | 4 | 0 |
| Molossus sp. | Curitiba-PR | 2015 | 2 | 0 |
| Eumops sp. | Foz do Iguaçu-PR | 2019–2020 | 1 | 0 |
| Molossus rufus | Braganey, Cascavel, Céu Azul, Francisco Beltrão, Jacarezinho, Londrina, Mamborê, Maringá, Maripá, Sarandi, Telêmaco Borba, Vera Cruz do Oeste-PR | 2019–2020 | 23 | 0 |
| Molossus molossus | Araucária, Assis Chateaubriand, Cascavel, Curitiba, Foz do Iguaçu, Guapirama, Guaratuba, Mandaguaçu, Maringá, Paulo Frontin, Ramilândia, Sarandi-PR | 2019–2020 | 29 | 0 |
| Molossus molossus | Poconé-MT | 2019 | 5 | 0 |
| Tadarida brasiliensis | Curitiba, Imbituva, Mamborê-PR | 2019–2020 | 5 | 0 |
| Promops nasutus | Cascavel, União da Vitória-PR | 2019–2020 | 3 | 0 |
| Eumops glaucinus | Assis Chateaubriand, Foz do Iguaçu, Maringá-PR | 2019–2020 | 3 | 0 |
| Molossops temminckii | Foz do Iguaçu-PR | 2019–2020 | 1 | 0 |
| Molossops neglectus | Salto do Lontra-PR | 2019–2020 | 1 | 0 |
| Molossus ater | Tapiraí-SP | 2018 | 1 | 0 |
| Nyctinomops sp. | Tapiraí-SP | 2019–2020 | 4 | 0 |
| Phyllostomidae Gray, 1825 | ||||
| Sturnira sp. | Curitiba-PR | 2019–2020 | 1 | 0 |
| Artibeus sp. | Foz do Iguaçu-PR | 2019–2020 | 1 | 0 |
| Artibeus lituratus | Rio de Janeiro-RJ | 2020 | 4 | 0 |
| Desmodus rotundus | Rio Branco do Sul-PR | 2015 | 6 | 0 |
| Diphylla ecaudata | Rio Branco do Sul-PR | 2015 | 2 | 0 |
| Glossophaga soricina | Poconé-MT | 2019 | 5 | 0 |
| Anoura caudifer | Tapiraí-SP | 2018 | 7 | 0 |
| Artibeus cinereus | Tapiraí-SP | 2018 | 2 | 0 |
| Artibeus fimbriatus | Tapiraí, Miracatu-SP | 2018 | 3 | 0 |
| Artibeus gnomus | Tapiraí-SP | 2018 | 1 | 0 |
| Artibeus lituratus | Tapiraí-SP | 2018 | 9 | 0 |
| Artibeus obscurus | Tapiraí-SP | 2018 | 9 | 0 |
| Artibeus planirostris | Tapiraí-SP | 2018 | 3 | 0 |
| Artibeus sp. | Tapiraí-SP | 2018 | 1 | 0 |
| Carollia perspicillata | Tapiraí, Miracatu-SP | 2018 | 18 | 0 |
| Chrotopterus auritus | Tapiraí-SP | 2018 | 1 | 0 |
| Desmodus rotundus | Tapiraí-SP | 2018 | 1 | 0 |
| Ectophylla sp. | Tapiraí-SP | 2018 | 1 | 0 |
| Lonchorhina aurita | Miracatu-SP | 2018 | 1 | 0 |
| Platyrrhinus lineatus | Tapiraí-SP | 2020 | 6 | 0 |
| Platyrrhinus sp. | Tapiraí-SP | 2020 | 1 | 0 |
| Rhinophylla pumilio | Tapiraí-SP | 2020 | 1 | 0 |
| Sturnira lilium | Tapiraí-SP | 2021 | 7 | 0 |
| Thrachops cirrhosus | Tapiraí-SP | 2021 | 1 | 0 |
| Uroderma bilobatum | Miracatu-SP | 2021 | 1 | 0 |
| Emballonuridae Gervais, 1856 | ||||
| Rhynchonycteris naso | Poconé-MT | 2019 | 2 | 0 |
| Pteropteryx sp. | Tapiraí-SP | 2020 | 3 | 0 |
| Noctilionidae Gray, 1821 | ||||
| Noctilio albiventris | Poconé-MT | 2019 | 1 | 1 (100%) |
| unknown | Curitiba, Paulo Frontin, Rolândia, Salto do Lontra-PR | 2019–2020 | 5 | 0 |
| TOTAL | 224 | 6 (2.67%) |
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| GenBank® Accession Number | Parasite Species | Host Species | Country of Source |
|---|---|---|---|
| MN316537, MN316538 | Haemocystidium cf. chelodinae | Myuchelys georgesi | Australia |
| MK976708-MK976710 | Haemocystidium pacayae | Podocnemis vogli | Colombia |
| MH177855 | Haemocystidium ptyodactylii | Squamate * | unknown |
| KT364883 | Haemocystidium sp. | Hemidactylus luqueorum | Oman |
| KT364884 | Haemocystidium sp. | Ptyodactylus hasselquistii | Oman |
| KX148083-KX148085 | Haemocystidium sp. | Kinixys erosa | Gabon |
| KX148088-KX148090 | Haemocystidium sp. | Kinixys erosa | Gabon |
| KX148086, KX148087 | Haemocystidium sp. | Pelusios castaneus | Gabon |
| MT684458 | Haemocystidium sp. | Podocnemis vogli | Colombia |
| MT684459 | Haemocystidium sp. | Trachylepis spilogaster | Angola |
| MT684460 | Haemocystidium sp. | Rhacodactylus auriculatus | New Caledonia |
| DQ630007 | Haemoproteus balmorali | Luscinia luscina | Lithuania |
| DQ630014 | Haemoproteus balmorali | Muscicapa striata | Lithuania |
| DQ630006 | Haemoproteus belopolskyi | Hippolais icterina | Sweden |
| MK843310 | Haemoproteus belopolskyi | Hippolais icterina | Lithuania |
| FJ168562 | Haemoproteus columbae | Columba livia | USA |
| MK843311 | Haemoproteus hirundinis | Delichon urbicum | Lithuania |
| KY653778 | Haemoproteus iwa | Fregata magnificens | Ecuador |
| KY653760 | Haemoproteus jenniae | Creagrus furcatus | Ecuador |
| DQ630010 | Haemoproteus lanii | Lanius collurio | Russia |
| MK843313 | Haemoproteus lanii | Lanius collurio | Lithuania |
| AY099045 | Haemoproteus majoris | Parus caeruleus | Sweden |
| JN164727, JN164728 | Haemoproteus majoris | Sylvia atricapilla | Spain |
| KU160476 | Haemoproteus minchini | Corythaeola cristata | Singapore |
| DQ630013 | Haemoproteus minutus | Turdus merula | Lithuania |
| KY653756 | Haemoproteus multipigmentatus | Zenaida galapagoensis | Ecuador |
| MK843312 | Haemoproteus nucleocondensus | Acrocephalus arundinaceus | Lithuania |
| JN164720 | Haemoproteus pallidulus | Sylvia atricapilla | Spain |
| DQ630004, DQ630005 | Haemoproteus pallidus | Ficedula hypoleuca | Sweden, Russia |
| JN164718, JN164719, JN164722 | Haemoproteus parabelopolskyi | Sylvia atricapilla | Spain |
| DQ630009 | Haemoproteus payevsky | Acrocephalus scipaceus | Lithuania |
| AY099040 | Haemoproteus sylvae | Acrocephalus arundinaceus | Sweden |
| OP503501 | Haemosporida sp. | Noctilio albiventris (ID Bat17) | Brazil |
| FJ168565 | Hepatocystis sp. | Pteropus hypomelanus | USA |
| JQ070951, JQ070956 | Hepatocystis sp. | Cercopithecus nictitans | Cameroon |
| FJ168563 | Leucocytozoon majoris | Zonotrichia leucophrys oriantha | USA |
| NC_012450 | Leucocytozoon majoris | Zonotrichia leucophrys oriantha | USA |
| KF159690 | Nycteria sp. | Rhinolophus landeri | Guinea |
| KF159720 | Nycteria sp. | Rhinolophus alcyone | Côte d’Ivoire |
| MK098843-MK098847 | Nycteria sp. | Rhinolophus sp., R. landeri | Gabon |
| FJ168561 | Parahaemoproteus vireonis | Vireo gilvus | USA |
| NC_012447 | Parahaemoproteus vireonis | Vireo gilvus | USA |
| HQ712051 | Plasmodium atheruri | Atherurus africanus | Madagascar |
| AY099055 | Plasmodium azurophilum | Anolis oculatus | Dominica |
| AY377128 | Plasmodium cathemerium | Serinus canaria | Germany |
| JN164734 | Plasmodium circumflexum | Sylvia atricapilla | Spain |
| AB444126 | Plasmodium cynomolgi | Monkey * | Japan |
| AF069611 | Plasmodium elongatum | Passer domesticus | North America |
| JF923762 | Plasmodium falciparum | Cercopithecus nictitans | Gabon |
| FJ895307 | Plasmodium gaboni | Pan sp. | Gabon |
| AY099053 | Plasmodium giganteum | Agama agama | Ghana |
| JF923751 | Plasmodium gonderi | Mandrillus sphinx | Gabon |
| JQ345504 | Plasmodium knowlesi | Homo sapiens | Malaysia |
| HM000110 | Plasmodium malariae | Pan troglodytes ellioti | Cameroon |
| OP503500 | Plasmodium malariae | Homo sapiens (ID I11) | Brazil |
| GU723548 | Plasmodium ovale | Homo sapiens | England |
| AY733090 | Plasmodium relictum | Hemignathus virens | USA |
| HM222485 | Plasmodium sp. | Icteria virens | USA |
| HM235065 | Plasmodium sp. | Gorilla sp. | Cameroon |
| HM235081 | Plasmodium sp. | Gorilla sp. | Cameroon |
| KF591834 | Plasmodium vivax | Homo sapiens | Congo |
| DQ414658 | Plasmodium yoelii killicki | Thamnomys rutilans | Congo |
| JN990708-JN990711 | Polychromophilus melanipherus | Miniopterus schreibersii | Switzerland |
| KJ131270-KJ131275 | Polychromophilus melanipherus | Miniopterus schreibersii | Europa |
| KU182361-KU182367 | Polychromophilus melanipherus | Nycteribia schmidlii scotti | Gabon |
| KU182368 | Polychromophilus melanipherus | Penicillidia fulvida | Gabon |
| MH744504, MH744505 | Polychromophilus melanipherus | Miniopterus mahafaliensis | Madagascar |
| MH744506, MH744519 | Polychromophilus melanipherus | Miniopterus griffithsi | Madagascar |
| MH744508 | Polychromophilus melanipherus | Miniopterus griveaudi | Madagascar |
| MH744522-MH744525 | Polychromophilus melanipherus | Miniopterus griveaudi | Madagascar |
| MH744509-MH744511 | Polychromophilus melanipherus | Miniopterus gleni | Madagascar |
| MH744518, MH744521 | Polychromophilus melanipherus | Miniopterus gleni | Madagascar |
| MH744512, MH744526 | Polychromophilus melanipherus | Miniopterus manavi | Madagascar |
| MH744514-MH744516 | Polychromophilus melanipherus | Miniopterus griveaudi | Madagascar |
| MH744520 | Polychromophilus melanipherus | Paratriaenops furculus | Madagascar |
| MH744527 | Polychromophilus melanipherus | Nycteribia stylidiopsis | Madagascar |
| MH744528-MH744531 | Polychromophilus melanipherus | Penicillidia leptothrinax | Madagascar |
| MK088162-MK088164, MK088168 | Polychromophilus melanipherus | Miniopterus orianae | Australia |
| MT136167 | Polychromophilus melanipherus | Taphozous melanopogon | Thailand |
| MW007671-MW007674 | Polychromophilus melanipherus | Nycteribia schmidlii scotti | South Africa |
| MW007676 | Polychromophilus melanipherus | Nycteribia schmidlii scotti | South Africa |
| MW007677 | Polychromophilus melanipherus | Miniopterus natalensis | South Africa |
| MW007680-MW007682 | Polychromophilus melanipherus | Nycteribia schmidlii | Hungary |
| MW007685 | Polychromophilus melanipherus | Nycteribia schmidlii | Spain |
| MW007689 | Polychromophilus melanipherus | Miniopterus schreibersii | Spain |
| HM055583 | Polychromophilus murinus | Myotis daubentonii | Switzerland |
| HM055583 | Polychromophilus murinus | Eptesicus serotinus | Switzerland |
| HM055583 | Polychromophilus murinus | Nyctalus noctula | Switzerland |
| HM055583 | Polychromophilus murinus | Myotis myotis | Switzerland |
| HM055584-HM055589 | Polychromophilus murinus | Myotis daubentonii | Switzerland |
| JN990712, JN990713 | Polychromophilus murinus | Myotis daubentonii | Switzerland |
| MH744532-MH744536 | Polychromophilus murinus | Myotis goudoti | Madagascar |
| MH744537 | Polychromophilus murinus | Penicillidia sp. | Madagascar |
| MT136168 | Polychromophilus murinus | Myotis siligorensis | Thailand |
| KF159675, KF159681 | Polychromophilus sp. | Miniopterus villiersi | Guinea |
| KF159699 | Polychromophilus sp. | Miniopterus villiersi | Guinea |
| KF159700 | Polychromophilus sp. | Neoromicia capensis | Guinea |
| LN483036 | Polychromophilus sp. | Rhinolophus sp. | Bulgaria |
| LN483038 | Polychromophilus sp. | Myotis nigricans | Panama |
| MK098848, MK098849 | Polychromophilus sp. | Miniopterus minor | Gabon |
| OP503502 | Polychromophilus sp. | Myotis riparius (ID 607) | Brazil |
| JQ995284-JQ995288 | Polychromophilus sp. | Miniopterus inflatus | Gabon |
| KF159714 | Polychromophilus sp. | Pipistrellus aff. grandidieri | Guinea |
| MT750305, MT750307, MT750308 | Polychromophilus sp. | Scotophilus kuhlii | Thailand |
| MW984518 | Polychromophilus sp. | Myotis ruber | Brazil |
| MW984519, MW984520 | Polychromophilus sp. | Myotis riparius | Brazil |
| MW984522 | Polychromophilus sp. | Myotis riparius | Brazil |
| MW984521 | Polychromophilus sp. | Eptesicus diminutus | Brazil |
| OQ957064 | Polychromophilus sp. | Myotis ruber (ID 125) | Brazil |
| OQ957065 | Polychromophilus sp. | Myotis sp. (ID 138) | Brazil |
| OQ957066 | Polychromophilus sp. | Myotis sp. (ID 141) | Brazil |
| Host Species | Parasite Species | cytb | asl | clpc | Country of Source |
|---|---|---|---|---|---|
| Miniopterus schreibersii | Polychromophilus melanipherus | JN990708 | - | JN990720 | Switzerland |
| Miniopterus schreibersii | Polychromophilus melanipherus | JN990709 | JN990726 | JN990721 | Switzerland |
| Miniopterus schreibersii | Polychromophilus melanipherus | JN990710 | - | JN990722 | Switzerland |
| Myotis daubentonii | Polychromophilus murinus | JN990712 | JN990725 | JN990723 | Switzerland |
| Myotis daubentonii | Polychromophilus murinus | JN990713 | - | JN990724 | Switzerland |
| Miniopterus villiersi | Polychromophilus sp. | KF159699 | - | KF159616 | Guinea |
| Neoromicia capensis | Polychromophilus sp. | KF159681 | - | KF159642 | Guinea |
| Pipistrellus aff. grandidieri | Polychromophilus sp. | KF159714 | - | KF159639 | Guinea |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750379 | KT750646 | KT750738 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750382 | KT750633 | - | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750380 | KT750647 | KT750740 | Kenya |
| Miniopterus rufus | Polychromophilus melanipherus | KT750385 | KT750637 | KT750745 | Kenya |
| Miniopterus rufus | Polychromophilus melanipherus | KT750386 | - | KT750748 | Kenya |
| Miniopterus sp. | Polychromophilus melanipherus | KT750387 | KT750642 | KT750749 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750377 | KT750629 | - | Kenya |
| Miniopterus africanus | Polychromophilus melanipherus | KT750375 | KT750627 | KT750734 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750400 | KT750630 | KT750737 | Kenya |
| Miniopterus rufus | Polychromophilus melanipherus | KT750403 | KT750636 | KT750744 | Kenya |
| Miniopterus rufus | Polychromophilus melanipherus | KT750404 | KT750639 | KT750746 | Kenya |
| Miniopterus rufus | Polychromophilus melanipherus | KT750418 | KT750641 | - | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750376 | KT750628 | KT750735 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750401 | KT750631 | KT750739 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750402 | KT750648 | KT750742 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750406 | - | KT750743 | Kenya |
| Miniopterus natalensis | Polychromophilus melanipherus | KT750378 | - | KT750736 | Kenya |
| Miniopterus minor | Polychromophilus sp. | KT750388 | KT750643 | KT750750 | Tanzania |
| Miniopterus minor | Polychromophilus sp. | KT750428 | KT750644 | KT750751 | Tanzania |
| Miniopterus minor | Polychromophilus sp. | KT750429 | KT750645 | - | Tanzania |
| Miniopterus sp. | Polychromophilus sp. | KT750389 | KT750552 | KT750651 | Mozambique |
| Miniopterus rufus | Polychromophilus sp. | KT750384 | KT750635 | - | Kenya |
| Miniopterus rufus | Polychromophilus sp. | KT750383 | KT750634 | - | Kenya |
| Miniopterus natalensis | Polychromophilus sp. | KT750381 | KT750632 | KT750741 | Kenya |
| Miniopterus rufus | Polychromophilus sp. | KT750412 | KT750638 | - | Kenya |
| Miniopterus rufus | Polychromophilus sp. | KT750405 | KT750640 | KT750747 | Kenya |
| Scotophilus kuhlii | Polychromophilus sp. | MT750307 | - | MT750315 | Thailand |
| Myotis macrodactylus | Polychromophilus murinus | LC668431 | - | LC715204 | Japan |
| Myotis macrodactylus | Polychromophilus murinus | LC668432 | - | LC715203 | Japan |
| Myotis macrodactylus | Polychromophilus murinus | LC668433 | - | LC715205 | Japan |
| Myotis riparius (ID 198) | Polychromophilus sp. | MW984519 | - | OP503503 | Brazil |
| Myotis riparius (ID 607) | Polychromophilus sp. | OP503502 | - | OP503504 | Brazil |
| Myotis ruber (ID 125) | Polychromophilus sp. | OQ957064 | OQ957067 | - | Brazil |
| Myotis sp. (ID 138) | Polychromophilus sp. | OQ957065 | - | - | Brazil |
| Myotis sp. (ID 141) | Polychromophilus sp. | OQ957066 | OQ957068 | OQ957063 | Brazil |
References
- Beltz, L.A. Bats and Human Health: Ebola, SARS, Rabies and Beyond; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Pacheco, M.A.; Escalante, A.A. Origin and diversity of malaria parasites and other Haemosporida. Trends Parasitol. 2023, 16, S1471-4922(23)00096-X. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-0-415-30097-1. [Google Scholar]
- Perkins, S.L.; Austin, C.C. Four New Species of Plasmodium from New Guinea Lizards: Integrating Morphology and Molecules. J. Parasitol. 2009, 95, 424–433. [Google Scholar] [CrossRef]
- Votýpka, J.; Modrý, D.; Oborník, M.; Šlapeta, J.; Lukeš, J. Apicomplexa. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 567–624. ISBN 978-3-319-28147-6. [Google Scholar]
- Perkins, S.L.; Schaer, J. A Modern Menagerie of Mammalian Malaria. Trends Parasitol. 2016, 32, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Carreno, R.A.; Kissinger, J.C.; McCutchan, T.F.; Barta, J.R. Phylogenetic Analysis of Haemosporinid Parasites (Apicomplexa: Haemosporina) and Their Coevolution with Vectors and Intermediate Hosts. Arch. Für Protistenkd. 1997, 148, 245–252. [Google Scholar] [CrossRef]
- Martinsen, E.S.; Perkins, S.L.; Schall, J.J. A Three-Genome Phylogeny of Malaria Parasites (Plasmodium and Closely Related Genera): Evolution of Life-History Traits and Host Switches. Mol. Phylogenet. Evol. 2008, 47, 261–273. [Google Scholar] [CrossRef]
- Schaer, J.; Perkins, S.L.; Decher, J.; Leendertz, F.H.; Fahr, J.; Weber, N.; Matuschewski, K. High Diversity of West African Bat Malaria Parasites and a Tight Link with Rodent Plasmodium Taxa. Proc. Natl. Acad. Sci. USA 2013, 110, 17415–17419. [Google Scholar] [CrossRef] [Green Version]
- Landau, I.; Rosin, G.; Miltgen, F.; Hugot, J.-P.; Leger, N.; Beveridge, I.; Baccam, D. Sur Le Genre Polycbromophilus: (Haemoproteidae, Parasite de Microchiroptères)1. Ann. Parasitol. Hum. Comp. 1980, 55, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Landau, I.; Baccam, D.; Ratanaworabhan, N.; Yenbutra, S.; Boulard, Y.; Chabaud, A.G. [New Haemoproteidae parasites of Chiroptera in Thailand]. Ann. Parasitol. Hum. Comp. 1984, 59, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Landau, I.; Chavatte, J.M.; Beveridge, I. Johnsprentia Copemani Gen. Nov., Sp. Nov. (Haemoproteidae), a Parasite of the Flying-Fox, Pteropus Alecto (Pteropidae). Mem. Qld. Mus. 2012, 56, 61–66, ISSN 0079-8835. [Google Scholar]
- Landau, I.; Chavatte, J.M.; Karadjian, G.; Chabaud, A.; Beveridge, I. The Haemosporidian Parasites of Bats with Description of Sprattiella Alecto Gen. Nov., Sp. Nov. Parasite 2012, 19, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Garnham, P.C.C.; Lainson, R.; Shaw, J.J. A Contribution to the Study of the Haematozoon Parasites of Bats. A New Mammalian Haemoproteid, Polychromophilus Deanei n. Sp. Mem. Inst. Oswaldo Cruz 1971, 69, 119–125. [Google Scholar] [CrossRef]
- Borner, J.; Pick, C.; Thiede, J.; Kolawole, O.M.; Kingsley, M.T.; Schulze, J.; Cottontail, V.M.; Wellinghausen, N.; Schmidt-Chanasit, J.; Bruchhaus, I.; et al. Phylogeny of Haemosporidian Blood Parasites Revealed by a Multi-Gene Approach. Mol. Phylogenet. Evol. 2016, 94, 221–231. [Google Scholar] [CrossRef]
- Minozzo, G.A.; da Silva Mathias, B.; Riediger, I.N.; de Oliveira Guimarães, L.; dos Anjos, C.C.; Monteiro, E.F.; dos Santos, A.P.; Biondo, A.W.; Kirchgatter, K. First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species. Microorganisms 2021, 9, 1240. [Google Scholar] [CrossRef]
- Correia dos Santos, L.; Vidotto, O.; dos Santos, N.J.R.; Ribeiro, J.; Pellizzaro, M.; dos Santos, A.P.; Haisi, A.; Wischral Jayme Vieira, T.S.; de Barros Filho, I.R.; Cubilla, M.P.; et al. Hemotropic Mycoplasmas (Hemoplasmas) in Free-Ranging Bats from Southern Brazil. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101416. [Google Scholar] [CrossRef]
- Perkins, S.L.; Schall, J.J. A molecular phylogeny of malarial parasites recovered from cytochrome b gene sequences. J. Parasitol. 2002, 88, 972–978. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut FigTree: Tree Figure Drawing Tool; Version 1.4.0; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2010.
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ruiz, F.; Linton, Y.-M.; Ponsonby, D.J.; Conn, J.E.; Herrera, M.; Quiñones, M.L.; Vélez, I.D.; Wilkerson, R.C. Molecular Comparison of Topotypic Specimens Confirms Anopheles (Nyssorhynchus) Dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem. Inst. Oswaldo Cruz 2010, 105, 899–903. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Matta, N.E.; Valkiūnas, G.; Parker, P.G.; Mello, B.; Stanley, C.E.; Lentino, M.; Garcia-Amado, M.A.; Cranfield, M.; Kosakovsky Pond, S.L.; et al. Mode and Rate of Evolution of Haemosporidian Mitochondrial Genomes: Timing the Radiation of Avian Parasites. Mol. Biol. Evol. 2018, 35, 383–403. [Google Scholar] [CrossRef] [Green Version]
- Rosyadi, I.; Shimoda, H.; Takano, A.; Yanagida, T.; Sato, H. Isolation and molecular characterization of Polychromophilus spp. (Haemosporida: Plasmodiidae) from the Asian long-fingered bat (Miniopterus fuliginosus) and Japanese large-footed bat (Myotis macrodactylus) in Japan. Parasitol. Res. 2022, 121, 2547–2559. [Google Scholar] [CrossRef]
- Simmons, N.B. Order Chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 312–529. ISBN 978-0-8018-8221-0. [Google Scholar]
- Abreu, E.F.; Casali, D.; Costa-Araújo, R.; Garbino, G.S.T.; Libardi, G.S.; Loretto, D.; Loss, A.C.; Marmontel, M.; Moras, L.M.; Nascimento, M.C.; et al. Lista de Mamíferos do Brasil (Version 2022-1) [Data Set]. Available online: https://zenodo.org/record/7469767 (accessed on 19 March 2022).
- Nogueira, M.R.; de Lima, I.P.; Moratelli, R.; da Cunha Tavares, V.; Gregorin, R.; Peracchi, A.L. Checklist of Brazilian Bats, with Comments on Original Records. Check List 2014, 10, 808–821. [Google Scholar] [CrossRef] [Green Version]
- Paglia, A.P.; Fonseca, G.A.B.; Rylands, A.B.; Herrmann, G.; Aguiar, L.M.S.; Chiarello, A.G.; Leite, Y.L.R.; Costa, L.P.; Siciliano, S.; Kierulff, A.M.; et al. Lista Anotada Dos Mamíferos Do Brasil. In Occasional Papers in Conservation Biology; Conservação Internacional: Belo Horizonte, MG, Brazil, 2012. [Google Scholar]
- Dias, D.; Esbérard, C.E.L.; Moratelli, R. A New Species of Lonchophylla (Chiroptera, Phyllostomidae) from the Atlantic Forest of Southeastern Brazil, with Comments on L. Bokermanni. Zootaxa 2013, 3722, 347. [Google Scholar] [CrossRef] [Green Version]
- Reis, N.R.; Fregonezi, M.N.; Peracchi, A.L.; Shibatta, O.A. Morcegos Do Brasil: Guia de Campo, 1st ed.; Série Manuais & guias TB; Technical Books Editora: Rio de Janeiro, Brazil, 2013; ISBN 978-85-61368-31-9. [Google Scholar]
- Reis, N.R.; Peracchi, A.L.; Batista, C.B.; de Lima, I.P.; Pereira, A.D. História Natural Dos Morcegos Brasileiros: Chave de Identificação de Espécies, 1st ed.; Technical Books Editora: Rio de Janeiro, Brazil, 2017; ISBN 978-85-61368-55-5. [Google Scholar]



| Sample ID | Blood Sample | Spleen Sample | Liver Sample |
|---|---|---|---|
| 125 | NA | Positive | Negative |
| 138 | Positive | Positive | Negative |
| 141 | Positive | NA | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathias, B.d.S.; Minozzo, G.A.; Biondo, A.W.; Costa, J.d.O.J.; Soares, H.S.; Marcili, A.; Guimarães, L.d.O.; Anjos, C.C.d.; Santos, A.P.D.; Riediger, I.N.; et al. Molecular Investigation Confirms Myotis Genus Bats as Common Hosts of Polychromophilus in Brazil. Microorganisms 2023, 11, 1531. https://doi.org/10.3390/microorganisms11061531
Mathias BdS, Minozzo GA, Biondo AW, Costa JdOJ, Soares HS, Marcili A, Guimarães LdO, Anjos CCd, Santos APD, Riediger IN, et al. Molecular Investigation Confirms Myotis Genus Bats as Common Hosts of Polychromophilus in Brazil. Microorganisms. 2023; 11(6):1531. https://doi.org/10.3390/microorganisms11061531
Chicago/Turabian StyleMathias, Bruno da Silva, Guilherme Augusto Minozzo, Alexander Welker Biondo, Jaciara de Oliveira Jorge Costa, Herbert Sousa Soares, Arlei Marcili, Lilian de Oliveira Guimarães, Carolina Clares dos Anjos, Andrea Pires Dos Santos, Irina Nastassja Riediger, and et al. 2023. "Molecular Investigation Confirms Myotis Genus Bats as Common Hosts of Polychromophilus in Brazil" Microorganisms 11, no. 6: 1531. https://doi.org/10.3390/microorganisms11061531