
Effect of Heavy-Metal-Resistant PGPR Inoculants on Growth, Rhizosphere Microbiome and Remediation Potential of Miscanthus × giganteus in Zinc-Contaminated Soil
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Preparation
2.2. Rhizobacterial Strains
2.3. Experimental Design
2.4. Plant Biomass Measurement
2.5. Measurement of Metal Content in Plant Biomass
2.6. 16S rRNA Gene-Based Metagenomic Analyses of Rhizosphere Soil and Bioinformatics
2.7. Statistics
3. Results
3.1. Characteristics of Bacterial Inoculants
3.2. Effect of Bacterial Inoculants and Zn on the Growth Performances of M. × giganteus
3.3. Effect of Bacterial Inoculants and Zn on Rhizosphere Microbial Communities of M. × giganteus
3.3.1. Diversity of Rhizosphere Microbial Communities
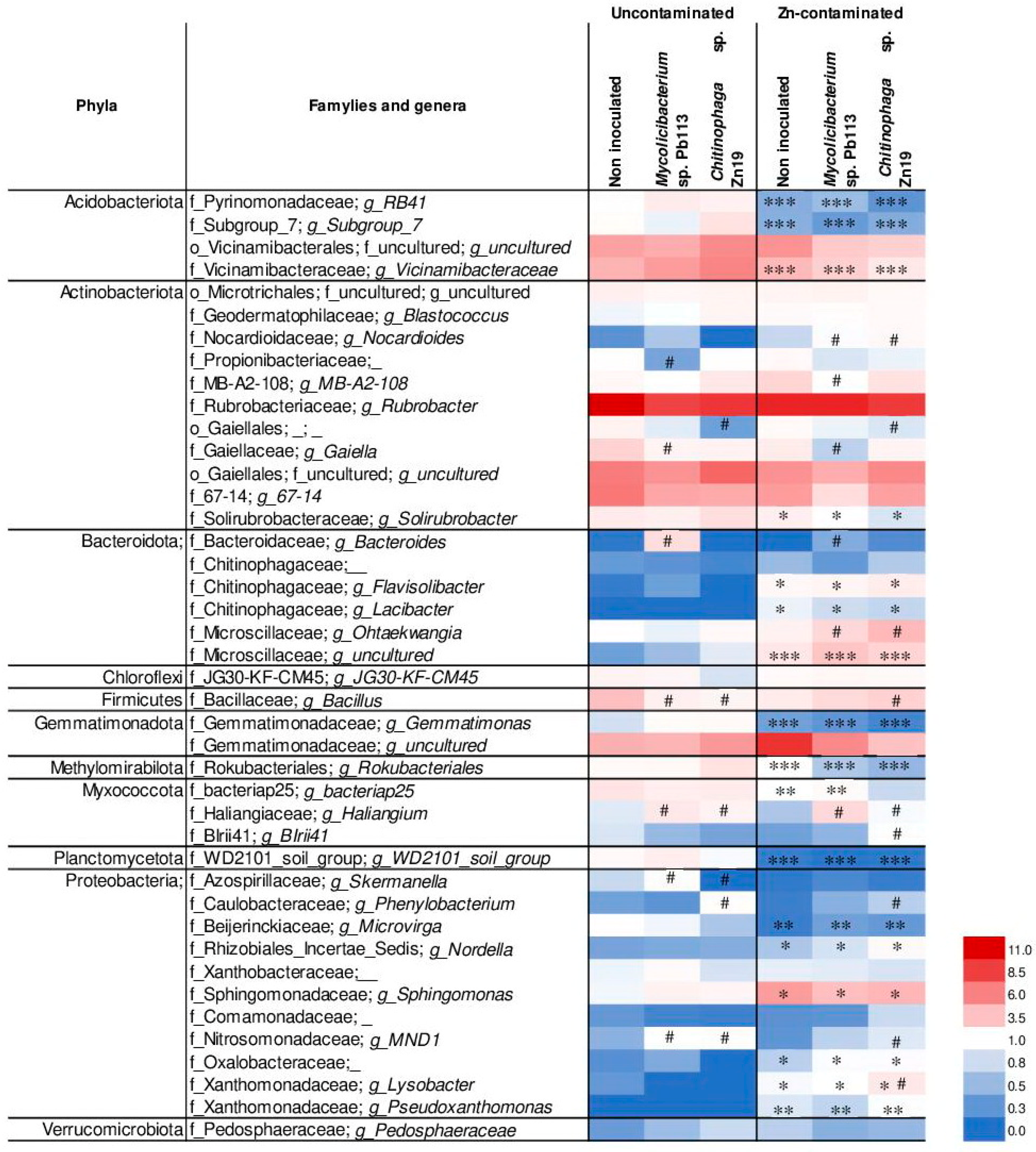
3.3.2. Taxonomic Structure of Rhizosphere Microbial Communities
3.4. Effect of Bacterial Inoculants on Zn Content in Plant Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Heaton, E.A.; Dohleman, F.G.; Miguez, A.F.; Juvik, J.A.; Lozovaya, V.; Widholm, J.; Zabotina, O.A.; McIsaac, G.F.; David, M.B.; Voigt, T.B.; et al. Chapter 3—Miscanthus: A promising biomass crop. In Advances in Botanical Research; Kader, J.-C., Delseny, M., Eds.; Academic Press: San Diego, CA, USA, 2010; Volume 56, pp. 75–137. [Google Scholar]
- Arnoult, S.; Brancourt-Hulmel, M.A. Review on Miscanthus biomass production and composition for bioenergy use: Genotypic and environmental variability and implications for breeding. BioEnergy Res. 2015, 8, 502–526. [Google Scholar] [CrossRef] [Green Version]
- Raikova, S.; Piccini, M.; Surman, M.K.; Allen, M.J.; Chuck, C.J. Making light work of heavy metal contamination: The potential for coupling bioremediation with bioenergy production. J. Chem. Technol. Biotechnol. 2019, 94, 3064–3072. [Google Scholar] [CrossRef]
- Wang, C.; Kong, Y.; Hu, R.; Zhou, G. Miscanthus: A fast-growing crop for environmental remediation and biofuel production. Glob. Chang. Biol. Bioenergy 2021, 13, 58–69. [Google Scholar] [CrossRef]
- Nurzhanova, A.; Pidlisnyuk, V.; Abit, K.; Nurzhanov, C.; Kenessov, B.; Stefanovska, T.; Erickson, L. Comparative assessment of using Miscanthus ×giganteus for remediation of soils contaminated by heavy metals: A case of military and mining sites. Environ. Sci. Pollut. Res. 2019, 26, 13320–13333. [Google Scholar] [CrossRef] [PubMed]
- Krzyżak, J.; Pogrzeba, M.; Rusinowski, S.; Clifton-Brown, J.; McCalmont, J.P.; Kiesel, A.; Mangold, A.; Mos, M. Heavy metal uptake by novel Miscanthus seed-based hybrids cultivated in heavy metal contaminated soil. Civ. Environ. Eng. Rep. 2017, 26, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Hussain, I.; Rasheed, R.; Iqbal, M.; Riaz, M.; Arif, M.S. Advances in microbe-assisted reclamation of heavy metal contaminated soils over the last decade: A review. J. Environ. Manag. 2017, 198, 132–143. [Google Scholar] [CrossRef]
- Pinto, A.P.; de Varennes, A.; Dias, C.M.B.; Lopes, M.E. Microbial-assisted phytoremediation: A convenient use of plant and microbes to clean up soils. In Phytoremediation. Management of Environmental Contaminants; Ansari, A., Gill, S., Gill, R., Lanza, G.R., Newman, L., Eds.; Springer: Cham, Switzerland, 2018; Volume 6, pp. 21–87. [Google Scholar]
- Hansda, A.; Kisku, P.C.; Kumar, V. Anshumali Plant-microbe association to improve phytoremediation of heavy metal. Chapter 6. In Advances in Microbe-Assisted Phytoremediation of Polluted Sites; Bauddh, K., Ma, Y., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2022; pp. 113–145. [Google Scholar]
- Kidd, P.S.; Álvarez-López, V.; Becerra-Castro, C.; Cabello-Conejo, M.; Prieto-Fernández, Á. Chapter 3—Potential role of plant-associated bacteria in plant metal uptake and implications in phytotechnologies. In Advances in Botanical Research; Cuypers, A., Vangronsveld, J., Eds.; Academic Press: San Diego, CA, USA, 2017; Volume 83, pp. 87–126. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Sheng, X.; He, L.; Wang, Q.; Ye, H.; Jiang, C. Effects of inoculation of biosurfactant-producing Bacillus sp. J119 on plant growth and cadmium uptake in a cadmium-amended soil. J. Hazard. Mater. 2008, 155, 17–22. [Google Scholar] [CrossRef]
- Pratush, A.; Kumar, A.; Hu, Z. Adverse effect of heavy metals (As, Pb, Hg, and Cr) on health and their bioremediation strategies: A review. Int. Microbiol. 2018, 21, 97–106. [Google Scholar] [CrossRef]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 109779. [Google Scholar] [CrossRef]
- Bourgeois, E.; Dequiedt, S.; Lelièvre, M.; van Oort, F.; Lamy, I.; Ranjard, L.; Maron, P.A. Miscanthus bioenergy crop stimulates nutrient-cycler bacteria and fungi in wastewater-contaminated agricultural soil. Environ. Chem. Lett. 2015, 13, 503–511. [Google Scholar] [CrossRef]
- Li, D.; Voigt, T.B.; Kent, A.D. Plant and soil effects on bacterial communities associated with Miscanthus × giganteus rhizosphere and rhizomes. GCB Bioenergy 2016, 8, 183–193. [Google Scholar] [CrossRef]
- Fan, D.; Schwinghamer, T.; Smith, D.L. Isolation and diversity of culturable rhizobacteria associated with economically important crops and uncultivated plants in Québec, Canada. Syst. Appl. Microbiol. 2018, 41, 629–640. [Google Scholar] [CrossRef]
- Pham, H.N.; Pham, P.A.; Nguyen, T.T.H.; Meiffren, G.; Brothier, E.; Lamy, I.; Michalet, S.; Dijoux-Franca, M.-G.; Nazaret, S. Influence of metal contamination in soil on metabolic profiles of Miscanthus × giganteus belowground parts and associated bacterial communities. Appl. Soil Ecol. 2018, 125, 240–249. [Google Scholar] [CrossRef]
- Zadel, U.; Nesme, J.; Michalke, B.; Vestergaard, G.; Płaza, G.A.; Schröder, P.; Radl, V.; Schloter, M. Changes induced by heavy metals in the plant-associated microbiome of Miscanthus × giganteus. Sci. Total Environ. 2020, 711, 134433. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, W.; Shao, Y.; Li, Y.-J.; Lin, L.-A.; Zhang, Y.-J.; Han, H.; Chen, Z.-J. Miscanthus cultivation shapes rhizosphere microbial community structure and function as assessed by Illumina MiSeq sequencing combined with PICRUSt and FUNGUIld analyses. Arch. Microbiol. 2020, 202, 1157–1171. [Google Scholar] [CrossRef]
- Nebeská, D.; Trögl, J.; Ševců, A.; Špánek, R.; Marková, K.; Davis, L.; Burdová, H.; Pidlisnyuk, V. Miscanthus × giganteus role in phytodegradation and changes in bacterial community of soil contaminated by petroleum industry. Ecotoxicol. Environ. Saf. 2021, 224, 112630. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Rocha, F.I.; Lee, J.; Choi, J.; Tejera, M.; Sooksa-Nguan, T.; Boersma, N.; Van Loocke, A.; Heaton, E.; Howe, A. The impact of stand age and fertilization on the soil microbiome of Miscanthus × giganteus. Phytobiomes J. 2021, 5, 51–59. [Google Scholar] [CrossRef]
- Babu, A.G.; Shea, P.J.; Sudhakar, D.; Jung, I.-B.; Oh, B.-T. Potential use of Pseudomonas koreensis AGB-1 in association with Miscanthus sinensis to remediate heavy metal(loid)-contaminated mining site soil. J. Environ. Manag. 2015, 151, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.S.; Mrnka, L.; Frantík, T.; Lovecká, P.; Vosátka, M. Plant growth promotion of Miscanthus × giganteus by endophytic bacteria and fungi on non-polluted and polluted soils. World J. Microbiol. Biotechnol. 2018, 34, 48. [Google Scholar] [CrossRef]
- Fei, H.; Crouse, M.; Papadopoulos, Y.A.; Vessey, J.K. Improving biomass yield of giant Miscanthus by application of beneficial soil microbes and a plant biostimulant. Can. J. Plant Sci. 2019, 100, 29–39. [Google Scholar] [CrossRef]
- Pranaw, K.; Pidlisnyuk, V.; Trögl, J.; Malinská, H. Bioprospecting of a novel plant growth-promoting bacterium Bacillus altitudinis KP-14 for enhancing Miscanthus × giganteus growth in metals contaminated soil. Biology 2020, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ludewig, U. Improved establishment of Miscanthus × giganteus stem propagation by Herbaspirillum inoculation. Ind. Crops Prod. 2020, 150, 112339. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.; Mamirova, A.; Pranaw, K.; Stadnik, V.; Kuráň, P.; Trögl, J.; Shapoval, P. Miscanthus giganteus phytoremediation of soil contaminated with trace elements as influenced by the presence of plant growth-promoting bacteria. Agronomy 2022, 12, 771. [Google Scholar] [CrossRef]
- Muratova, A.Y.; Sungurtseva, I.Y.; Nurzhanova, A.A.; Turkovskaya, O.V. Rhizosphere microbiome of the bioenergetic metal-resistant plant Miscanthus × giganteus and its remediation potential. In Smart and Innovative Farming for Sustainable Agriculture and Food Systems; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar]
- GN 2.1.7.2511-09; Orientirovochno Dopustimye Koncentracii (ODK) Himicheskih Veshchestv v Pochve: Gigienicheskie Normativy—M.: Federal’nyj Centr Gigieny i Epidemiologii Rospotrebnadzora. Approximate Permissible Concentrations (APC) of Chemical Substances in Soils. Federal Center for Hygiene and Epidemiology of Rospotrebnadzor: Moscow, Russia, 2009; 10p. Available online: https://files.stroyinf.ru/Data2/1/4293828/4293828439.pdf (accessed on 21 February 2023). (In Russian)
- Muratova, A.Y.; Lyubun, Y.V.; Sungurtseva, I.Y.; Nurzhanova, A.A.; Turkovskaya, O.V. Physiological and biochemical responses of Miscanthus × giganteus to soil contamination with heavy metals. Ecobiotech 2019, 2, 482–493, (In Russian, abstract in English). [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Verma, V.; Joshi, K.; Mazumdar, B. Study of siderophore formation in nodule-forming bacterial species. Res. J. Chem. Sci. 2012, 2, 26–29. [Google Scholar]
- Muromtsev, G.S. Some methods for studying the dissolution of calcium phosphates by microorganisms. Microbiologiya 1957, 26, 172–178. (In Russian) [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Golubev, S.N.; Muratova, A.Y.; Panchenko, L.V.; Shchyogolev, S.Y.; Turkovskaya, O.V. Mycolicibacterium sp. strain PAM1, an alfalfa rhizosphere dweller, catabolizes PAHs and promotes partner-plant growth. Microbiol. Res. 2021, 253, 126885. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muratova, A.; Gorelova, S.; Golubev, S.; Kamaldinova, D.; Gins, M. Rhizosphere microbiomes of Amaranthus spp. grown in soils with anthropogenic polyelemental anomalies. Agronomy 2023, 13, 759. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 62, 346–351. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kocoń, A.; Matyka, M. Phytoextractive potential of Miscanthus giganteus and Sida hermaphrodita growing under moderate pollution of soil with Zn and Pb. J. Food Agric. Environ. 2012, 10, 1253–1256. [Google Scholar]
- Zgorelec, Z.; Bilandzija, N.; Knez, K.; Galic, M.; Zuzul, S. Cadmium and Mercury phytostabilization from soil using Miscanthus ×giganteus. Sci. Rep. 2020, 10, 6685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, B.; Boléo, S.; Sidella, S.; Costa, J.; Duarte, M.P.; Mendes, B.; Cosentino, S.L.; Fernando, A.L. Phytoremediation of heavy metal-contaminated soils using the perennial energy crops Miscanthus spp. and Arundo donax L. Bioenergy Res. 2015, 8, 1500–1511. [Google Scholar] [CrossRef]
- Muratova, A.; Lyubun, Y.; Sungurtseva, I.; Turkovskaya, O.; Nurzhanova, A. Physiological and biochemical characteristic of Miscanthus × giganteus grown in heavy metal–oil sludge co-contaminated soil. J. Environ. Sci. 2022, 115, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef] [Green Version]
- Mirza, B.; Rodrigues, J. Development of a direct isolation procedure for free-living diazotrophs under controlled hypoxic conditions. Appl. Environ. Microbiol. 2012, 78, 5542–5549. [Google Scholar] [CrossRef] [Green Version]
- Moyes, A.; Kueppers, L.; Pett-Ridge, J.; Carper, D.; Vandehey, N.; O’Neil, J.; Frank, A.C. Evidence for foliar endophytic nitrogen fixation in a widely distributed subalpine conifer. New Phytol. 2016, 210, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Rilling, J.I.; Acuña, J.J.; Sadowsky, M.J.; Jorquera, M.A. Putative nitrogen-fixing bacteria associated with the rhizosphere and root endosphere of wheat plants grown in an andisol from Southern Chile. Front. Microbiol. 2018, 20, 2710. [Google Scholar] [CrossRef] [Green Version]
- Chanda, D.; Sharma, G.D.; Jha, D.K.; Hijri, M.; Al-Otaibi, F.A. Isolation and characterization of Chitinophaga sp. from paper mill polluted soil. Int. J. Sci. Res. 2014, 3, 513–516. [Google Scholar]
- Sanjaya, W.T.A.; Khoirunnisa, N.S.; Ismiani, S.; Hazra, F.; Santosa, D.A. Isolation and characterization of mercury-resistant microbes from gold mine area in Mount Pongkor, Bogor District, Indonesia. Biodiversitas 2021, 22, 2656–2666. [Google Scholar] [CrossRef]
- Augelletti, F.; Tremblay, J.; Agathos, S.N.; Jousset, A.; Stenuit, B. Draft whole-genome sequence of the anthracene-degrading strain Mycolicibacterium frederiksbergense LB501T, isolated from a polycyclic aromatic hydrocarbon-contaminated soil. Microbiol. Resour. Announc. 2020, 9, e00671-20. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Klimova, S.Y.; Shestakov, A.I.; Botina, S.G.; Netrusov, A.I. Orchid-associated bacteria produce indole-3-acetic acid, promote seed germination, and increase their microbial yield in response to exogenous auxin. Arch. Microbiol. 2007, 188, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Dell’Amico, E.; Cavalca, L.; Andreoni, V. Improvement of Brassica napus growth under cadmium stress by cadmium-resistant rhizobacteria. Soil Biol. Biochem. 2008, 40, 74–84. [Google Scholar] [CrossRef]
- Egamberdiyeva, D. Colonization of Mycobacterium phlei in the rhizosphere of wheat grown under saline conditions. Turk. J. Biol. 2012, 36, 487–492. [Google Scholar] [CrossRef]
- Rupaedah, B.; Anas, I.; Santosa, D.A.; Sumaryono, W.; Budi, S.W. Screening and characterization of rhizobacteria for enhancing growth and chlorophyll content of sweet sorghum (Sorghum bicolor L. Moench). J. Int. Soc. Southeast Asian Agric. 2014, 20, 86–97. [Google Scholar]
- Li, N.; Liu, R.; Chen, J.; Wang, J.; Hou, L.; Zhou, Y. Enhanced phytoremediation of PAHs and cadmium contaminated soils by a Mycobacterium. Sci. Total Environ. 2021, 754, 141198. [Google Scholar] [CrossRef]
- Yang, J.; Gu, Y.; Chen, Z.; Song, Y.; Sun, F.; Liu, J.; Waigi, M. Colonization and performance of a pyrene-degrading bacterium Mycolicibacterium sp. Pyr9 on root surfaces of white clover. Chemosphere 2021, 263, 127918. [Google Scholar] [CrossRef]
- Chimwamurombe, P.M.; Grönemeyer, J.L.; Reinhold-Hurek, B. Isolation and characterization of culturable seed-associated bacterial endophytes from gnotobiotically grown Marama bean seedlings. FEMS Microbiol. Ecol. 2016, 92, fiw083. [Google Scholar] [CrossRef] [Green Version]
- Shet, S.A.; Garg, S. Plant growth promotion of Vigna unguiculata in arid sandy soil using bacterial species from coastal sand dune. Agric. Res. 2022, 11, 716–727. [Google Scholar] [CrossRef]
- Hu, J.; Yang, T.; Friman, V.-P.; Kowalchuk, G.A.; Hautier, Y.; Li, M.; Wei, Z.; Xu, Y.; Shen, Q.; Jousset, A. Introduction of probiotic bacterial consortia promotes plant growth via impacts on the resident rhizosphere microbiome. Proc. R. Soc. B 2021, 288, 20211396. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Rafiq, M.T.; Khan, K.Y.; Pan, F.; Yang, X.; Feng, Y. Improvement of cadmium uptake and accumulation in Sedum alfredii by endophytic bacteria Sphingomonas SaMR12: Effects on plant growth and root exudates. Chemosphere 2014, 17, 367–373. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, N.; Guo, X.; Zhang, Y.; Ye, B. Comparative analysis of bacterial community structure in the rhizosphere of maize by high-throughput pyrosequencing. PLoS ONE 2017, 12, e0178425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Babu, A.G.; Kim, J.-D.; Oh, B.-T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard. Mater. 2013, 250–251, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-J.; Tian, W.; Li, Y.-J.; Sun, L.-N.; Chen, Y.; Zhang, H.; Li, Y.Y.; Han, H. Responses of rhizosphere bacterial communities, their functions and their network interactions to Cd stress under phytostabilization by Miscanthus spp. Environ. Pollut. 2021, 287, 117663. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Feng, Z.; Tomura, T.; Suzuki, A.; Miyano, S.; Tsuge, T.; Mori, H.; Suh, J.-W.; Iizuka, T.; Fudou, R.; et al. Heterologous production of the marine myxobacterial antibiotic haliangicin and its unnatural analogues generated by engineering of the biochemical pathway. Sci. Rep. 2016, 6, 22091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-K.; Chiang, M.-S.; Hseu, Z.-Y.; Kuo, C.-H.; Liu, C.-T. A photosynthetic bacterial inoculant exerts beneficial effects on the yield and quality of tomato and affects bacterial community structure in an organic field. Front. Microbiol. 2022, 13, 959080. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Romanyà, J.; Li, N.; Xiang, Y.; Yang, J.; Han, X. Biochar fertilization effects on soil bacterial community and soil phosphorus forms depends on the application rate. Sci. Total Environ. 2022, 843, 157022. [Google Scholar] [CrossRef]
- Ofek-Lalzar, M.; Sela, N.; Goldman-Voronov, M.; Green, S.J.; Hadar, Y.; Minz, D. Niche and host-associated functional signatures of the root surface microbiome. Nat. Commun. 2014, 5, 4950. [Google Scholar] [CrossRef] [Green Version]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Ferreira, A.; Nobre, M.; Moore, E.M.; Rainey, F.A.; Battista, J.R.; da Costa, M.S. Characterization and radiation resistance of new isolates of Rubrobacter radiotolerans and Rubrobacter xylanophilus. Extremophiles 1999, 3, 235–238. [Google Scholar] [CrossRef]
- Khudur, L.S.; Gleeson, D.B.; Ryan, M.H.; Shahsavari, E.; Haleyur, N.; Nugegoda, D.; Ball, A.S. Implications of co-contamination with aged heavy metals and total petroleum hydrocarbons on natural attenuation and ecotoxicity in Australian soils. Environ. Pollut. 2018, 243, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Czarny, J.; Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Juzwa, W.; Wolniewicz, A.; Marecik, R.; Ławniczak, Ł.; Chrzanowski, Ł. Acinetobacter sp. as the key player in diesel oil degrading community exposed to PAHs and heavy metals. J. Hazard. Mater. 2020, 383, 121168. [Google Scholar] [CrossRef]
- Caracciolo, A.B.; Terenzi, V. Rhizosphere microbial communities and heavy metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef] [PubMed]
- Bilal, S.; Khan, A.L.; Shahzad, R.; Kim, Y.-H.; Imran, M.; Khan, M.J.; Al-Harrasi, A.; Kim, T.H.; Lee, I.-J. Mechanisms of Cr(VI) resistance by endophytic Sphingomonas sp. LK11 and its Cr(VI) phytotoxic mitigating effects in soybean (Glycine max L.). Ecotoxicol. Environ. Saf. 2018, 164, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Mahbub, K.R.; Krishnan, K.; Naidu, R.; Megharaj, M. Mercury resistance and volatilization by Pseudoxanthomonas sp. SE1 isolated from soil. Environ. Technol. Innov. 2016, 6, 94–104. [Google Scholar] [CrossRef]

 —uncontaminated, non-inoculated;
—uncontaminated, non-inoculated;  —uncontaminated + Mycolicibacterium sp. Pb113;
—uncontaminated + Mycolicibacterium sp. Pb113;  —uncontaminated + Chitinophaga sp. Zn19;
—uncontaminated + Chitinophaga sp. Zn19;  —Zn-contaminated, non-inoculated;
—Zn-contaminated, non-inoculated;  —Zn-contaminated + Mycolicibacterium sp. Pb-113;
—Zn-contaminated + Mycolicibacterium sp. Pb-113;  —Zn-contaminated + Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—uncontaminated, non-inoculated; —uncontaminated + Mycolicibacterium sp. Pb113; —uncontaminated + Chitinophaga sp. Zn19; —Zn-contaminated, non-inoculated; —Zn-contaminated + Mycolicibacterium sp. Pb-113; —Zn-contaminated + Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—Zn-contaminated + Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—uncontaminated, non-inoculated; —uncontaminated + Mycolicibacterium sp. Pb113; —uncontaminated + Chitinophaga sp. Zn19; —Zn-contaminated, non-inoculated; —Zn-contaminated + Mycolicibacterium sp. Pb-113; —Zn-contaminated + Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
 —rhizome biomass at planting; —rhizome biomass at the end of cultivation; —non-inoculated control; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—rhizome biomass at planting; —rhizome biomass at the end of cultivation; —non-inoculated control; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—rhizome biomass at planting; —rhizome biomass at the end of cultivation; —non-inoculated control; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—rhizome biomass at planting; —rhizome biomass at the end of cultivation; —non-inoculated control; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.


 —non-inoculated; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—non-inoculated; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—non-inoculated; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.
—non-inoculated; —Mycolicibacterium sp. Pb113; —Chitinophaga sp. Zn19. Values represent means, and bars represent confidence interval (n ≥ 3); different letters mean significant difference between treatments at p ≤ 0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Pb113 | Zn19 |
|---|---|---|
| PGPR traits: | ||
| nitrogen fixation | + | + |
| phosphorous mobilization | − | − |
| siderophore synthesis | + | + |
| IAA production | + | + |
| Metal resistance (MIC, mmol/L) | ||
| Zn | 3.0 | 2.5 |
| Pb | ≥5.0 | 2.0 |
| Soil | Inoculation | Observed Features | Chao1 | Shannon Index | Simpson Index | Faith PD |
|---|---|---|---|---|---|---|
| Uncontaminated | Non-inoculated | 294 ± 64 | 293.60 ± 64.31 | 7.5813 ± 0.2880 | 0.9928 ± 0.0013 | 29.96 ± 7.95 |
| Pb113 | 378 ± 16 | 379.38 ± 16.09 | 8.0089 ± 0.0316 | 0.9949 ± 0.0000 | 32.52 ± 2.96 | |
| Zn19 | 298 ± 5 | 298.10 ± 4.10 | 7.6623 ± 0.0471 | 0.9935 ± 0.0002 | 28.10 ± 2.20 | |
| Zn-contaminated | Non-inoculated | 294 ± 16 | 295.17 ±17.21 | 7.5218 ± 0.0345 | 0.9922 ± 0.0003 | 26.78 ± 5.88 |
| Pb113 | 346 ± 14 | 347.55 ± 14.42 | 7.7014 ± 0.0351 | 0.9930 ± 0.0018 | 36.66 ± 5.39 | |
| Zn19 | 414 ± 6 | 414.56 ± 6.29 | 7.9681 ± 0.0554 | 0.9942 ± 0.0007 | 30.41 ± 3.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muratova, A.; Golubev, S.; Romanova, V.; Sungurtseva, I.; Nurzhanova, A. Effect of Heavy-Metal-Resistant PGPR Inoculants on Growth, Rhizosphere Microbiome and Remediation Potential of Miscanthus × giganteus in Zinc-Contaminated Soil. Microorganisms 2023, 11, 1516. https://doi.org/10.3390/microorganisms11061516
Muratova A, Golubev S, Romanova V, Sungurtseva I, Nurzhanova A. Effect of Heavy-Metal-Resistant PGPR Inoculants on Growth, Rhizosphere Microbiome and Remediation Potential of Miscanthus × giganteus in Zinc-Contaminated Soil. Microorganisms. 2023; 11(6):1516. https://doi.org/10.3390/microorganisms11061516
Chicago/Turabian StyleMuratova, Anna, Sergey Golubev, Valeria Romanova, Irina Sungurtseva, and Asil Nurzhanova. 2023. "Effect of Heavy-Metal-Resistant PGPR Inoculants on Growth, Rhizosphere Microbiome and Remediation Potential of Miscanthus × giganteus in Zinc-Contaminated Soil" Microorganisms 11, no. 6: 1516. https://doi.org/10.3390/microorganisms11061516