

Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk and Humans in Eastern Tanzania: Genetic Diversity and Inter-Host Transmission
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Site
2.3. Study Design
2.4. Sample Collection
2.5. Identification
2.6. DNA Extraction and Whole Genome Sequencing
2.7. Bioinformatic Analysis
3. Results
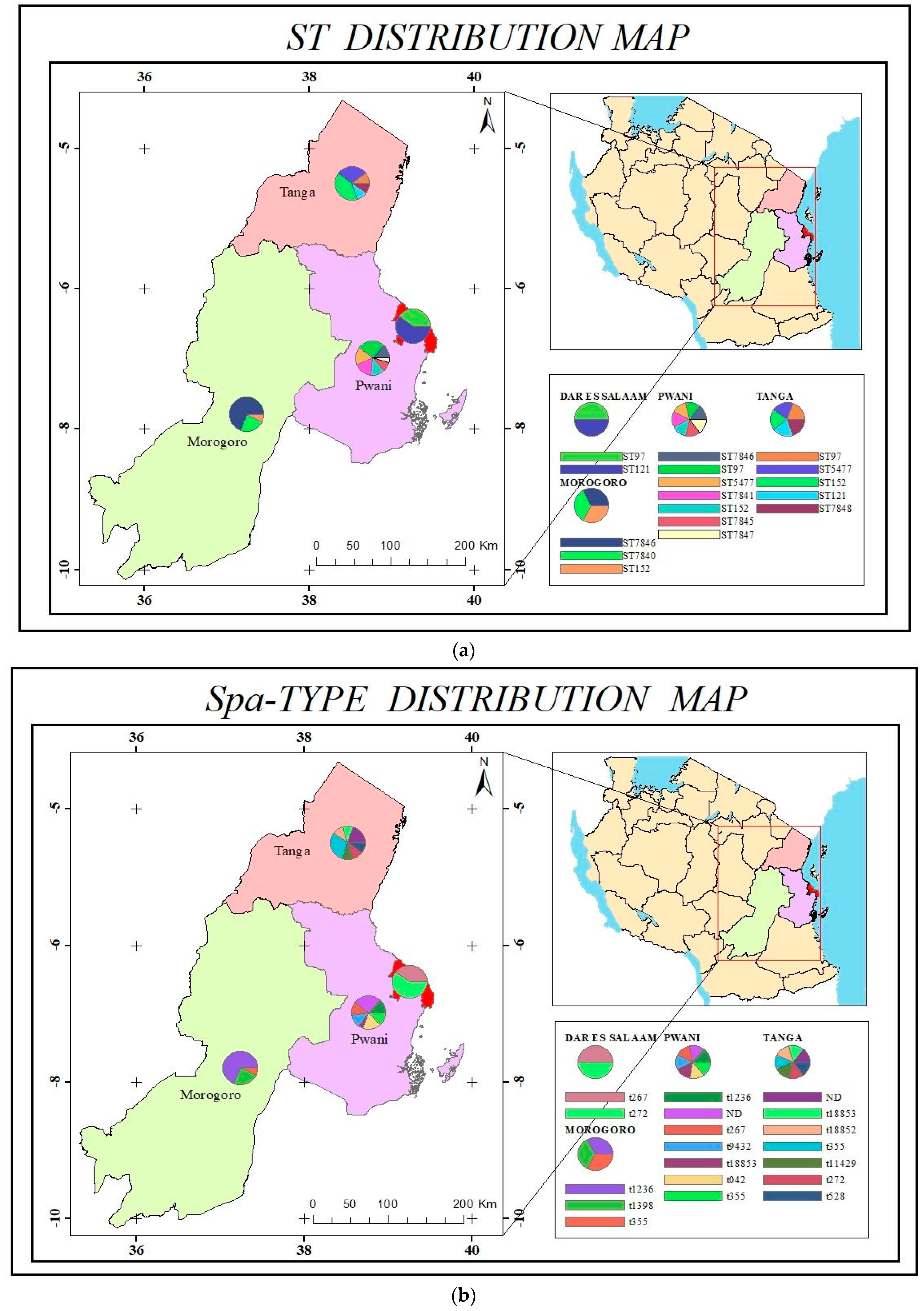
3.1. Isolates Characteristics, Species Identification, and Multilocus Sequence Typing
3.2. Antimicrobial Resistance, Virulence, Leukocidin Genes, and Spa Typing
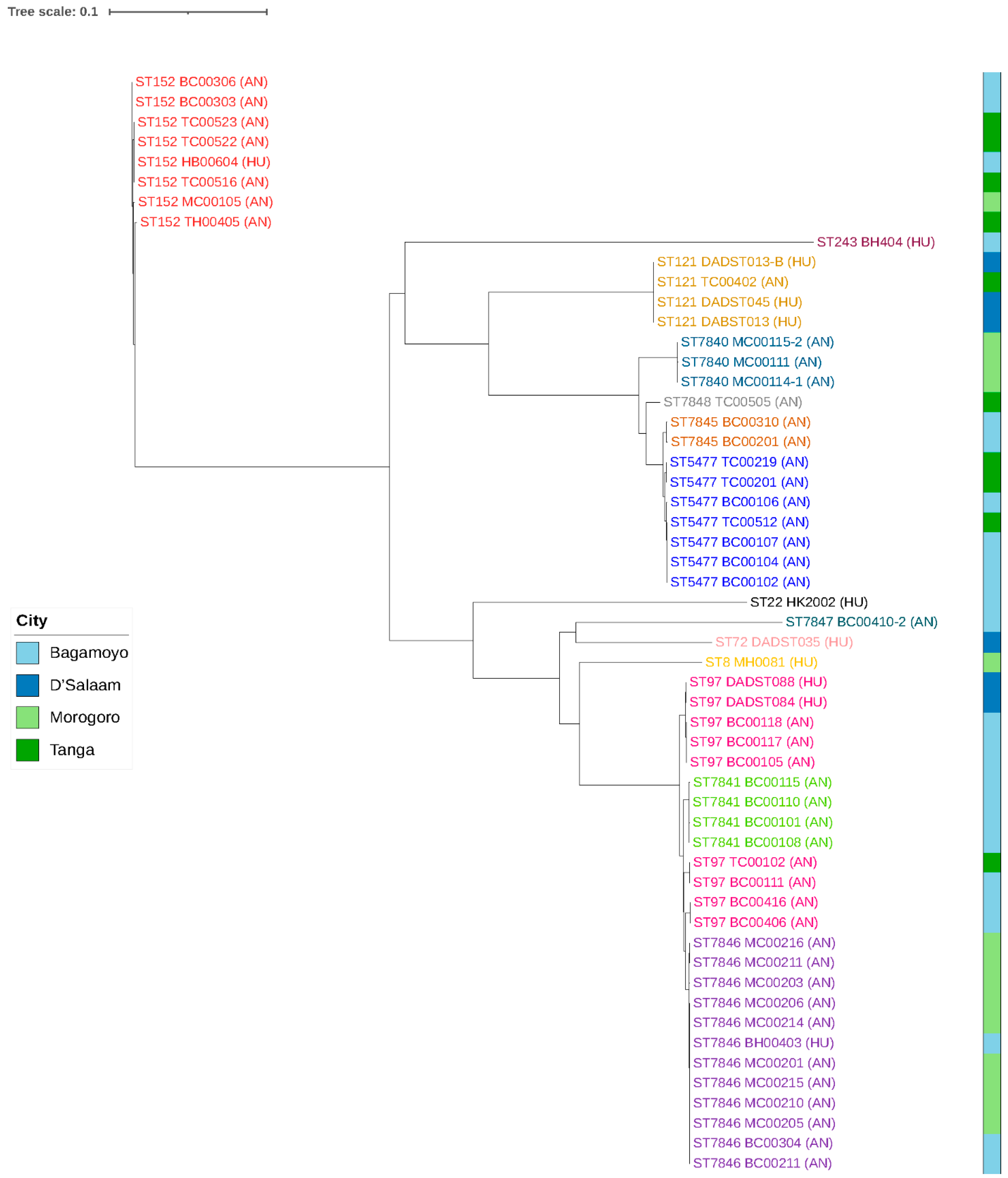
3.3. Single Nucleotide Polymorphisms (SNPs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blomberg, B.; Mwakagile, D.S.; Urassa, W.K.; Maselle, S.Y.; Mashurano, M.; Digranes, A.; Harthug, S.; Langeland, N. Surveillance of Antimicrobial Resistance at a Tertiary Hospital in Tanzania. BMC Public Health 2004, 4, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivaria, F.M.; Noordhuizen, J. A retrospective study of the aetiology and temporal distribution of bovine clinical mastitis in smallholder dairy herds in the Dar es Salaam region of Tanzania. Vet. J. 2007, 173, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.L.; Kamphuis, C.; Martins CM, M.R.; Barreiro, J.R.; Tomazi, T.; Gameiro, A.H.; Hogeveen Hdos Santos, M.V. Bovine subclinical mastitis reduces milk yield and economic return. Livest. Sci. 2018, 210, 25–32. [Google Scholar] [CrossRef]
- Tamba, M.; Rocca, R.; Prosperi, A.; Pupillo, G.; Bassi, P.; Galletti, G.; Martini, E.; Santi, A.; Casadei, G.; Arrigoni, N. Evaluation of Control Program against Streptococcus agalactiae Infection in Dairy Herds during 2019–2021 in Emilia-Romagna Region, Northern Italy. Front. Vet. Sci. 2022, 9, 904527. [Google Scholar] [CrossRef] [PubMed]
- Budd, K.E.; McCoy, F.; Monecke, S.; Cormican, P.; Mitchell, J.; Keane, O.M. Extensive Genomic Diversity among Bovine-Adapted Staphylococcus aureus: Evidence for a Genomic Rearrangement within CC97. PLoS ONE 2015, 10, e0134592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakwinska, O.; Giddey, M.; Moreillon, M.; Morisset, D.; Waldvogel, A.; Moreillon, P. Staphylococcus aureus Host Range and Human-Bovine Host Shift. Appl. Environ. Microbiol. 2011, 77, 5908–5915. [Google Scholar] [CrossRef] [Green Version]
- Campos, B.; Pickering, A.; Rocha, C.; Lis Souza, A.; Fabres-Klein, P.A.; de Oliveira Mendes, M.H.; Fitzgerald, T.A.; Ross, J.; de Oliveira Barros Ribon, A. Diversity and pathogenesis of Staphylococcus aureus from bovine mastitis: Current understanding and future perspectives. BMC Vet. Res. 2022, 18, 115. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Scott, N.L.; Sordillo, L.M. Ability of Staphylococcusaureus Coagulase Genotypes to Resist Neutrophil Bactericidal Activity and Phagocytosis. Infect. Immun. 1994, 62, 5679–5682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabello, R.F.; Moreira, B.M.; Lopes RM, M.; Teixeira, L.M.; Riley, L.W.; Castro, A.C.D. Multilocus sequence typing of Staphylococcus aureus isolates recovered from cows with mastitis in Brazilian dairy herds. J. Med. Microbiol. 2007, 56 Pt 11, 1505–1511. [Google Scholar] [CrossRef] [Green Version]
- Monistero, V.; Graber, H.U.; Pollera, C.; Cremonesi, P.; Castiglioni, B.; Bottini, E.; Ceballos-Marquez, A.; Lasso-Rojas, L.; Kroemker, V.; Wente, N.; et al. Staphylococcus aureus Isolates from Bovine Mastitis in Eight Countries: Genotypes, Detection of Genes Encoding Different Toxins and Other Virulence Genes. Toxins 2018, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.; Kock, M.M.; Ehlers, M.M. Molecular Characterization of Staphylococcus aureus Isolated from Bovine Mastitis and Close Human Contacts in South African Dairy Herds: Genetic Diversity and Inter-Species Host Transmission. Front. Microbiol. 2017, 8, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndahetuye, J.B.; Leijon, M.; Båge, R.; Artursson, K.; Persson, Y. Genetic Characterization of Staphylococcus aureus from Subclinical Mastitis Cases in Dairy Cows in Rwanda. Front. Vet. Sci. 2021, 8, 1319. [Google Scholar] [CrossRef] [PubMed]
- Müller-Premru, M.; Strommenger, B.; Alikadic, N.; Witte, W.; Friedrich, A.W.; Seme, K.; Svent, K.N.; Smrke, D.; Spik, V.; Gubina, M. New strains of community-acquired methicillin-resistant Staphylococcus aureus with Panton–Valentine leukocidin causing an outbreak of severe soft tissue infection in a football team. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Abdulgader, S.M.A.; Shittu, A.O.; Nicol, M.P.; Kaba, M. Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: A systematic review. Front. Microbiol. 2015, 6, 348. [Google Scholar] [CrossRef]
- Juhász-Kaszanyitzky, E.; Jánosi, S.; Somogyi Pdán, A.; van der Graaf-van Bloois, L.; van Duijkeren, E.; Wagenaar, J.A. MRSA transmission between cows and humans. Emerg. Infect. Dis. 2007, 13, 630–632. [Google Scholar] [CrossRef]
- Algammal, A.M.; El-Tarabili, E.; Mohamed, E.; Ghobashy, R.M.; Madeha, O.I.; Helmy, Y.A. Prevalence, Antimicrobial Resistance Profiles, Virulence and Enterotoxins-Determinant Genes of MRSA Isolated from Subclinical Bovine Mastitis in Egypt. Pathogens 2020, 9, 362. [Google Scholar] [CrossRef]
- A Dinges, M.M.; Orwin, M.; Schlievert, M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Younis, A.; Krifucks, O.; Fleminger, G.H.; Elimelech, D.G.; Natan, S.; Arthur, L.G. Staphylococcus aureus leucocidin, a virulence factor in bovine mastitis. J. Dairy Res. 2005, 72, 188–194. [Google Scholar] [CrossRef]
- El Bayomi, R.M.; Ahmed, H.A.; Awadallah, M.A.; Mohsen, R.A.; Abd El-Ghafar, A.E.; Abdelrahman, M.A. Occurrence, Virulence Factors, Antimicrobial Resistance, and Genotyping of Staphylococcus aureus Strains Isolated from Chicken Products and Humans. Vector Borne Zoonotic Dis. 2016, 16, 157–164. [Google Scholar] [CrossRef]
- Votintseva, A.A.; Fung, R.M.; Ruth, R.K.; Kyle, G.H.; Wyllie, D.H.; Bowden, R.C.; Derrick, W.; Walker, A.S. Prevalence of Staphylococcus aureus protein A (spa) mutants in the community and hospitals in Oxfordshire. BMC Microbiol. 2014, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Hakimi Alni, R.; Mohammadzadeh, A.; Mahmoodi, P. Molecular typing of Staphylococcus aureus of different origins based on the polymorphism of the spa gene: Characterization of a novel spa type. 3 Biotech 2018, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Ngoa, U.A.; Kösters, K.; Köck, R.; Adegnika, A.A.; Kremsner, P.G.; Lell, B.; Peters, G.; Mellmann, A.; Becker, K. Virulence factors and genotypes of Staphylococcus aureus from infection and carriage in Gabon. Clin. Microbiol. Infect. 2011, 17, 1507–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nhatsave, N.; Garrine, M.M.; Augusto, M.; Arsénia, J.; Cossa, A.; Vaz, R.; Ombi, A.; Zimba, T.F.; Alfredo, H.; Mandomando, I.; et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique. Microorganisms 2021, 9, 1684. [Google Scholar] [CrossRef] [PubMed]
- Essack, S.Y.; Desta, A.T.; Abotsi, R.E.; Agoba, E.E. Antimicrobial resistance in the WHO African region: Current status and roadmap for action. J. Public Health 2016, 39, 8–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shittu, A.O.; Taiwo, F.F.; Froböse, N.J.; Schwartbeck, B.; Niemann, S.; Mellmann, A.; Schaumburg, F. Genomic analysis of Staphylococcus aureus from the West African Dwarf (WAD) goat in Nigeria. Antimicrob. Resist. Infect. Control. 2021, 10, 122. [Google Scholar] [CrossRef]
- Kimang’a, A.N. A Situational Analysis of Antimicrobial Drug Resistance in Africa: Are We Losing the Battle? Ethiop. J. Health Sci. 2012, 22, 135–143. [Google Scholar]
- United republic of Tanzania, Ministry of Livestock and Fisheries. Tanzania Livestock Sector Analysis (2016-2017-2031-2032). 2017. Available online: https://www.mifugouvuvi.go.tz/uploads/projects/1553602287-LIVESTOCK%20SECTOR%20ANALYSIS.pdf (accessed on 8 December 2022).
- Sudhanthiramani, S.; Swetha, C.S.; Bharathy, S. Prevalence of antibiotic resistant Staphylococcus aureus from raw milk samples collected from the local vendors in the region of Tirupathi, India. Vet. World 2015, 8, 478–481. [Google Scholar] [CrossRef] [Green Version]
- Boerlin, P.; Boerlin, P.; Kuhnert, P.; Hüssy, D.; Schaellibaum, M. Methods for identification of Staphylococcus aureus isolates in cases of bovine mastitis. J. Clin. Microbiol. 2003, 41, 767–771. [Google Scholar] [CrossRef] [Green Version]
- Spoor, L.E.; McAdam, P.R.; Weinert, L.A.; Rambaut, A.; Hasman, H.; Aarestrup, F.M.; Kearns, K.A.; Larsen, A.R.; Skov, L.A.; Fitzgerald, J.R. Livestock Origin for a Human Pandemic Clone of Community-Associated Methicillin-Resistant Staphylococcus aureus. MBio 2013, 4, e00356-13. [Google Scholar] [CrossRef] [Green Version]
- Yebra, G.; Harling-Lee, J.D.; Lycett, S.; Aarestrup, F.M.; Larsen, G.; Cavaco, L.M.; Seo, K.S.; Abraham, S.; Norris, J.M.; Schmidt, T.; et al. Multiclonal human origin and global expansion of an endemic bacterial pathogen of livestock. Proc. Natl. Acad. Sci. USA 2022, 119, 5. [Google Scholar] [CrossRef]
- Ruffing, U.; Alabi, A.; Kazimoto, T.; Vubil, D.C.; Akulenko, R.; Abdulla, S.; Alonso, P.; Bischoff, M.; Germann, A.; Grobusch, M.P.; et al. Community-Associated Staphylococcus aureus from Sub-Saharan Africa and Germany: A Cross-Sectional Geographic Correlation Study. Sci. Rep. 2017, 7, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekonnen, S.A.; Lam, T.J.G.M.; Hoekstra, J.; Rutten, V.P.M.G.; Tessema, T.S.; Broens, E.M.; Riesebos, A.E.; Spaninks, M.P.; Koop, G. Characterization of Staphylococcus aureus isolated from milk samples of dairy cows in small holder farms of North-Western Ethiopia. BMC Vet. Res. 2018, 14, 246. [Google Scholar] [CrossRef] [PubMed]
- Sangeda, R.Z.; Baha, A.; Erick, A.; Mkumbwa, S.; Bitegeko, A.; Sillo, H.B.; Fimbo, A.M.; Chambuso, M.; Mbugi, E.V. Consumption Trends of Antibiotic for Veterinary Use in Tanzania: A Longitudinal Retrospective Survey from 2010–2017. Front. Trop. Dis. 2021, 2, 694082. [Google Scholar] [CrossRef]
- Penadés, M.; Viana, D.; García-Quirós, A.; Muñoz-Silvestre, A.; Moreno-Grua, E.; Pérez-Fuentes, S.; Pascual, J.J.; Corpa, J.M.; Selva, L. Differences in virulence between the two more prevalent Staphylococcus aureus clonal complexes in rabbitries (CC121 and CC96) using an experimental model of mammary gland infection. Vet. Res. 2020, 51, 11. [Google Scholar] [CrossRef] [Green Version]
- Egyir, B.; Hadjirin, N.; Gupta, S.; Owusu, F.; Agbodzi, B.; Adogla-Bessa, T.; Addo, K.; Stegger, M.; Larsen, A.R.; Holmes, M. Whole-genome sequence profiling of antibiotic-resistant Staphylococcus aureus isolates from livestock and farm attendants in Ghana. J. Glob. Antimicrob. Resist. 2020, 22, 527–532. [Google Scholar] [CrossRef]
- Vasquez, M.T.; Lubkin, A.; Reyes-Robles, T.; Day, C.J.; Lacey, K.A.; Jennings, M.P.; Torres, V.J. Dentification of a domain critical for Staphylococcus aureus LukED receptor targeting and lysis of erythrocytes. J. Biol. Chem. 2020, 295, 17241–17250. [Google Scholar] [CrossRef]
- Ortega, E.; Abriouel, H.; Lucas, R.; Gálvez, A. Multiple roles of Staphylococcus aureus enterotoxins: Pathogenicity, superantigenic activity, and correlation to antibiotic resistance. Toxins 2010, 2, 2117–2131. [Google Scholar] [CrossRef] [Green Version]
- Ahmadrajabi, R.; Layegh-Khavidaki, S.; Kalantar-Neyestanaki, D.; Fasihi, Y. Molecular analysis of immune evasion cluster (IEC) genes and intercellular adhesion gene cluster (ICA) among methicillin-resistant and methicillin-sensitive isolates of Staphylococcus aureus. J. Prev. Med. Hyg. 2017, 58, E308–E314. [Google Scholar]
- Verkaik, N.J.; Benard, M.; Boelens, H.A.; de Vogel, C.P.; Nouwen, J.L.; Verbrugh, H.A.; Melles, D.C.; van Belkum, A.; van Wamel, W.J. Immune evasion cluster-positive bacteriophages are highly prevalent among human Staphylococcus aureus strains, but they are not essential in the first stages of nasal colonization. Clin. Microbiol. Infect. 2011, 17, 343–348. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| (a) | ||||
|---|---|---|---|---|
| Sample Type | Source | Region | No. of Isolates | Sequence Type (n) |
| Milk | Cow | Tanga | 9 | ST97 (1) |
| ST5477 (3) | ||||
| ST121 (1) | ||||
| ST7848 (1) | ||||
| ST152 (3) | ||||
| Milk | Cow | Bagamoyo | 21 | ST7841 (4) |
| ST5477 (5) | ||||
| ST97 (5) | ||||
| ST7845 (2) | ||||
| ST7846 (2) | ||||
| ST152 (2) | ||||
| ST7847 (1) | ||||
| Milk | Cow | Morogoro | 13 | ST152 (1) |
| ST7840 (3) | ||||
| ST7846 (9) | ||||
| (b) | ||||
| Sample Type | Source | Region | No. of Isolates | Sequence Type (n) |
| Nasal Swab | Human | Bagamoyo | 4 | ST22 (1) |
| ST7846 (1) | ||||
| ST243 (1) | ||||
| ST152 (1) | ||||
| Tanga | 1 | ST152 | ||
| Morogoro | 1 | ST8 | ||
| Dar es Salaam | 6 | ST72 (1) | ||
| ST121 (3) | ||||
| ST97 (2) | ||||
| Sample ID | Origin | Region | ST | CC | AMR Genes | Virulence Genes | Toxin Genes | Leukocide Genes | Spa Type |
|---|---|---|---|---|---|---|---|---|---|
| BC00211 | M | BAG | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| BC00304 | M | BAG | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00201 | M | MOR | blaZ, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00203 | M | MOR | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00205 | M | MOR | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00210 | M | MOR | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00211 | M | MOR | 7846 | 97 | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 |
| MC00214 | M | MOR | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00215 | M | MOR | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00216 | M | MOR | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| MC00206 | M | MOR | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| BH00403 | HWA | BAG | blaZ, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t1236 | ||
| TC00102 | M | TAN | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | ND | ||
| BC00105 | M | BAG | blaZ, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn | lukD, lukE | t267 | ||
| BC00117 | M | BAG | blaZ, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn | lukD, lukE | t267 | ||
| BC00118 | M | BAG | blaZ, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn | lukD, lukE | t267 | ||
| BC00406 | M | BAG | 97 | 97 | blaZ, qacG, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t9432 |
| BC00416 | M | BAG | blaZ, qacG, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t9432 | ||
| BC00111 | M | BAG | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t9432 | ||
| DADST084 | HNA | DSM | blaZ | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn | lukD, lukE | t267 | ||
| DADST088 | HNA | DSM | blaZ | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn | lukD, lukE | t267 | ||
| TC00201 | M | TAN | blaZ, str | aur, edinB, hlgA, hlgB, hlgC, splA, splB | sei, sem, sen, tst | lukE | t18853 | ||
| TC00219 | M | TAN | blaZ, str | aur, edinB, hlgA, hlgB, hlgC, splA, splB | sei, sem, sen, seo, tst | lukE | t18852 | ||
| TC00512 | M | TAN | blaZ | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, sen, seu, tst | lukD, lukE | ND | ||
| BC00102 | M | BAG | 5477 | 5477 | blaZ | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, sen, seo, seu, tst | lukE | ND |
| BC00104 | M | BAG | blaZ | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, seu, tst | lukE | ND | ||
| BC00106 | M | BAG | blaZ | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, seo, tst | lukE | ND | ||
| BC00107 | M | BAG | blaZ | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, sen, seo, tst | lukE | ND | ||
| BC00108 | M | BAG | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB, splE | ND | lukD, lukE | t042 | ||
| BC00110 | M | BAG | blaZ, str | aur, hlgA, hlgB, hlgC, splA, splB, splE | ND | lukD, lukE | t042 | ||
| BC00101 | M | BAG | 7841 | 97 | str | aur, hlgA, hlgB, hlgC, splA, splB, splE | ND | lukD, lukE | t042 |
| BC00115 | M | BAG | blaZ | aur, hlgA, hlgB, hlgC, splA, splB | ND | lukD, lukE | t042 | ||
| MC00111 | M | MOR | erm(C) | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, seu | lukE | t1398 | ||
| MC00114 | M | MOR | 7840 | 5477-like | ND | aur, edinB, hlgA, hlgB, hlgC, splA, splB | sei | lukE | t1398 |
| MC00115 | M | MOR | ND | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, sen, seo, seu, | lukE | t1398 | ||
| TC00516 | M | TAN | blaZ, dfrG, erm(C) | edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 | ||
| TC00522 | M | TAN | blaZ, dfrG, erm(C) | aur, edinB, hlgA | sak, scn | lukF-PV, lukS-PV | t355 | ||
| TC00523 | M | TAN | blaZ, dfrG, erm(C) | aur, edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 | ||
| BC00303 | M | BAG | 152 | 152 | dfrG, erm(C), tet(K) | aur, edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 |
| BC00306 | M | BAG | blaZ, dfrG, erm(C), tet(K) | edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 | ||
| MC00105 | M | MOR | blaZ, dfrG, tet(K) | edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 | ||
| TH00405 | HWA | TAN | blaZ, tet(K) | hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t11429 | ||
| HB00604 | HWA | BAG | blaZ, dfrG, erm(C) | aur, edinB, hlgA, hlgB | sak, scn | lukF-PV, lukS-PV | t355 | ||
| TC00402 | M | TAN | blaZ, dfrG, erm(C) | aur, edinC, hlgA, hlgB, hlgC, splA, splB | sak, scn, seg, sei, sem, sen, seo, seu, eta, etb | ND | t272 | ||
| DABST013 | HNA | DSM | 121 | 121 | blaZ, dfrG, erm(C) | aur, edinC, hlgA, hlgB, hlgC, splA | sak, scn, seg, sei, sem, sen, seu, eta, etb | ND | t272 |
| DADST045 | HNA | DSM | blaZ, dfrG, erm(C), fosB, tet(K) | aur, edinC, hlgA, hlgB, hlgC, splA, splB | sak, scn, sei, sem, sen, seo, seu, eta, etb | ND | t272 | ||
| DADST013B | HNA | DSM | dfrG, erm(C) | aur, hlgA, hlgB, splA, splB | sak, scn, sei, eta | ND | t272 | ||
| BC00310 | M | BAG | blaZ | aur, edinB, hlgA, hlgB, hlgC | ND | lukE | ND | ||
| BC00201 | M | BAG | 7845 | 5477-like | blaZ, str, tet(K) | aur, edinB, hlgA, hlgB, hlgC, splA, splB | ND | lukE | t18853 |
| BC00410-2 | M | BAG | 7847 | qacG | aur, hlgA, hlgB, hlgC, splA, splB, splE | ND | lukE | ND | |
| TC00505 | M | TAN | 7848 | 5477-like | tet(K) | aur, edinB, hlgA, hlgC, splA, splB | sei, sem, sen, seo | lukE | t528 |
| HK2002 | HWA | BAG | 22 | blaZ | aur, hlgA, hlgB, hlgC | sak, scn, seg, sei, sem, sen, seo, seu, tst | ND | t223 | |
| BH00404 | HWA | BAG | 243 | ND | aur, hlgA, hlgB, hlgC, splE | seg, sei, sem, sen, seo, seu | lukF-PV, lukS-PV | t021 | |
| MH00801 | HWA | MOR | 8 | aac(6’)-aph(″), blaZ, dfrG, erm(C), mecA, qacD, tet(K) | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn, seb, sej, sek, seq, ser | lukD, lukE | t1476 | |
| DADST035 | HNA | DSM | 72 | blaZ, dfrG | aur, hlgA, hlgB, hlgC, splA, splB, splE | sak, scn, sec, seg, sei, sel, sem, sen, seo, seu, tst | lukD, lukE | t148 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mzee, T.; Kumburu, H.; Kazimoto, T.; Leekitcharoenphon, P.; van Zwetselaar, M.; Masalu, R.; Mlaganile, T.; Sonda, T.; Wadugu, B.; Mushi, I.; et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk and Humans in Eastern Tanzania: Genetic Diversity and Inter-Host Transmission. Microorganisms 2023, 11, 1505. https://doi.org/10.3390/microorganisms11061505
Mzee T, Kumburu H, Kazimoto T, Leekitcharoenphon P, van Zwetselaar M, Masalu R, Mlaganile T, Sonda T, Wadugu B, Mushi I, et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk and Humans in Eastern Tanzania: Genetic Diversity and Inter-Host Transmission. Microorganisms. 2023; 11(6):1505. https://doi.org/10.3390/microorganisms11061505
Chicago/Turabian StyleMzee, Tutu, Happiness Kumburu, Theckla Kazimoto, Pimlapas Leekitcharoenphon, Marco van Zwetselaar, Rose Masalu, Tarsis Mlaganile, Tolbert Sonda, Boaz Wadugu, Ignass Mushi, and et al. 2023. "Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk and Humans in Eastern Tanzania: Genetic Diversity and Inter-Host Transmission" Microorganisms 11, no. 6: 1505. https://doi.org/10.3390/microorganisms11061505