
Antimicrobial Resistance Risk Assessment of Vibrio parahaemolyticus Isolated from Farmed Green Mussels in Singapore

 , , , and
, , , and
Abstract
:1. Introduction
2. Methodology
2.1. Survey Data Collection for Exposure Assessment
2.2. Sample Collection
2.2.1. Farm and Retail Sampling
2.2.2. Sample Processing
2.2.3. Presumptive Vibrio Species Direct Plate Counting and Phenotypic Screening of Haemolytic Strains
2.2.4. Purification of Bacterial Strains and Glycerol Stocking
2.2.5. 16s rRNA Gene Amplification and Sequencing
2.2.6. Antimicrobial Susceptibility Testing
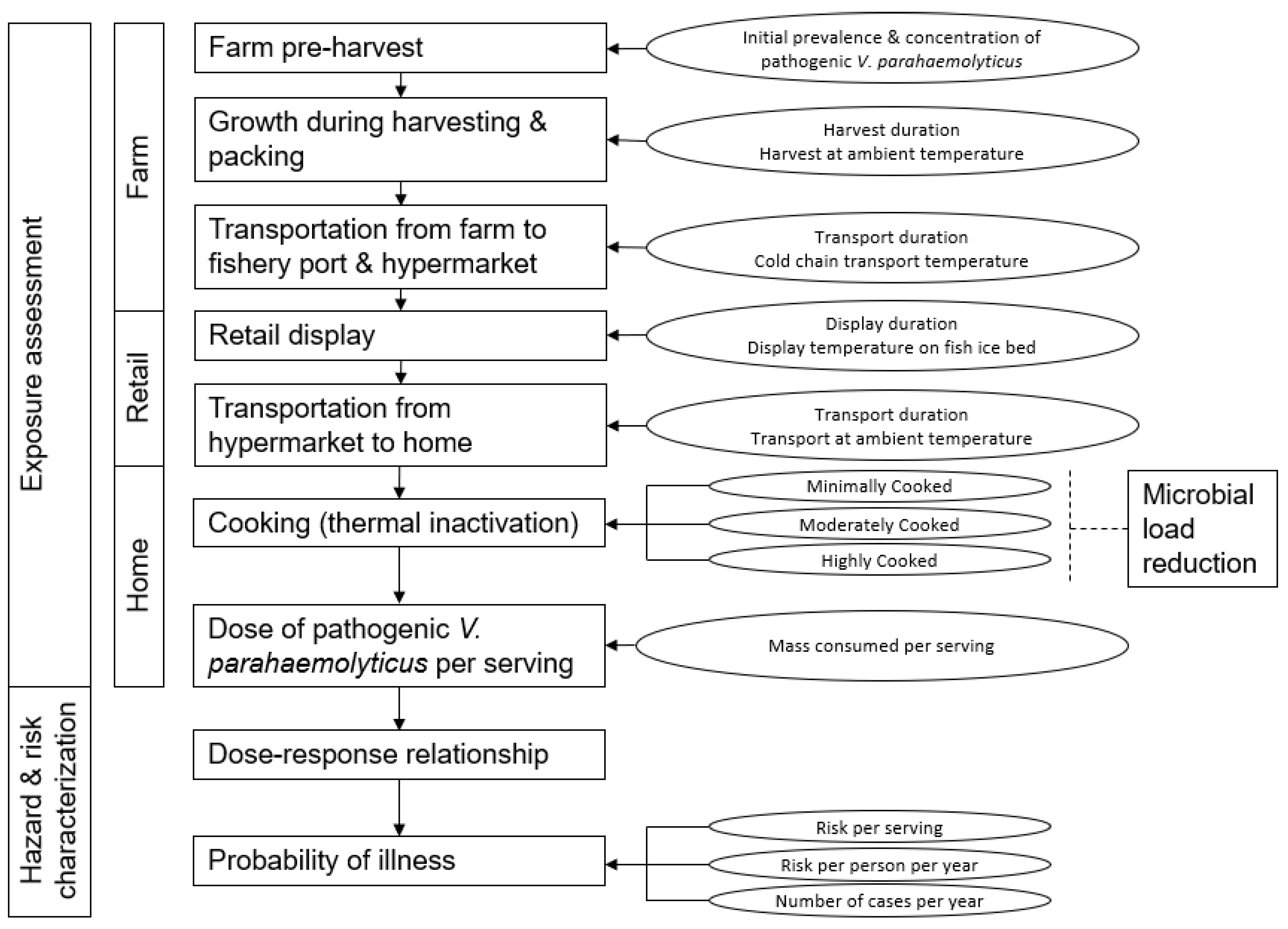
2.3. Risk Modelling Framework
- Green mussels harvested from the farm were processed on site before being packed on ice. Products were firstly transported to the fishery port, where they were sorted and eventually sent to the hypermarket. At the hypermarket, the seafood was packaged in plastic and placed on a chiller for display.
- Equal survivability fitness was assumed across different V. parahaemolyticus strains.
- Cross- and co-resistance traits of V. parahaemolyticus strains to the other studied resistance traits were disregarded within each ARRA for specific antimicrobial resistance.
- Only direct exposure from contaminated green mussels through consumption was considered. Indirect transmission modes of infection and transmission to workers in the food chain were disregarded.
- Variable human-host immune responses to V. parahaemolyticus infection were disregarded.
2.4. Hazard Identification
2.5. Exposure Assessment
2.5.1. V. parahaemolyticus Growth Rate Modelling and Adjustment Factors
2.5.2. Hemolytic V. parahaemolyticus Occurrence and Concentration Levels at Pre-Harvest and Retail
2.5.3. Parameters for Harvesting and Transportation to the Hypermarket Retailer
2.5.4. Parameters for Retail Display and Transportation to Home
2.5.5. Parameters for Preparation, Cooking and Consumption of Green Mussels
2.6. Hazard Characterisation
Dose–Response Relationship
2.7. Risk Characterisation
3. Results
3.1. Haemolytic, Ampicillin, Penicillin G, and Tetracycline Resistant Vibrio parahaemolyticus Occurrence and Concentration Levels
3.2. Risk Estimate Outputs across ARRAs
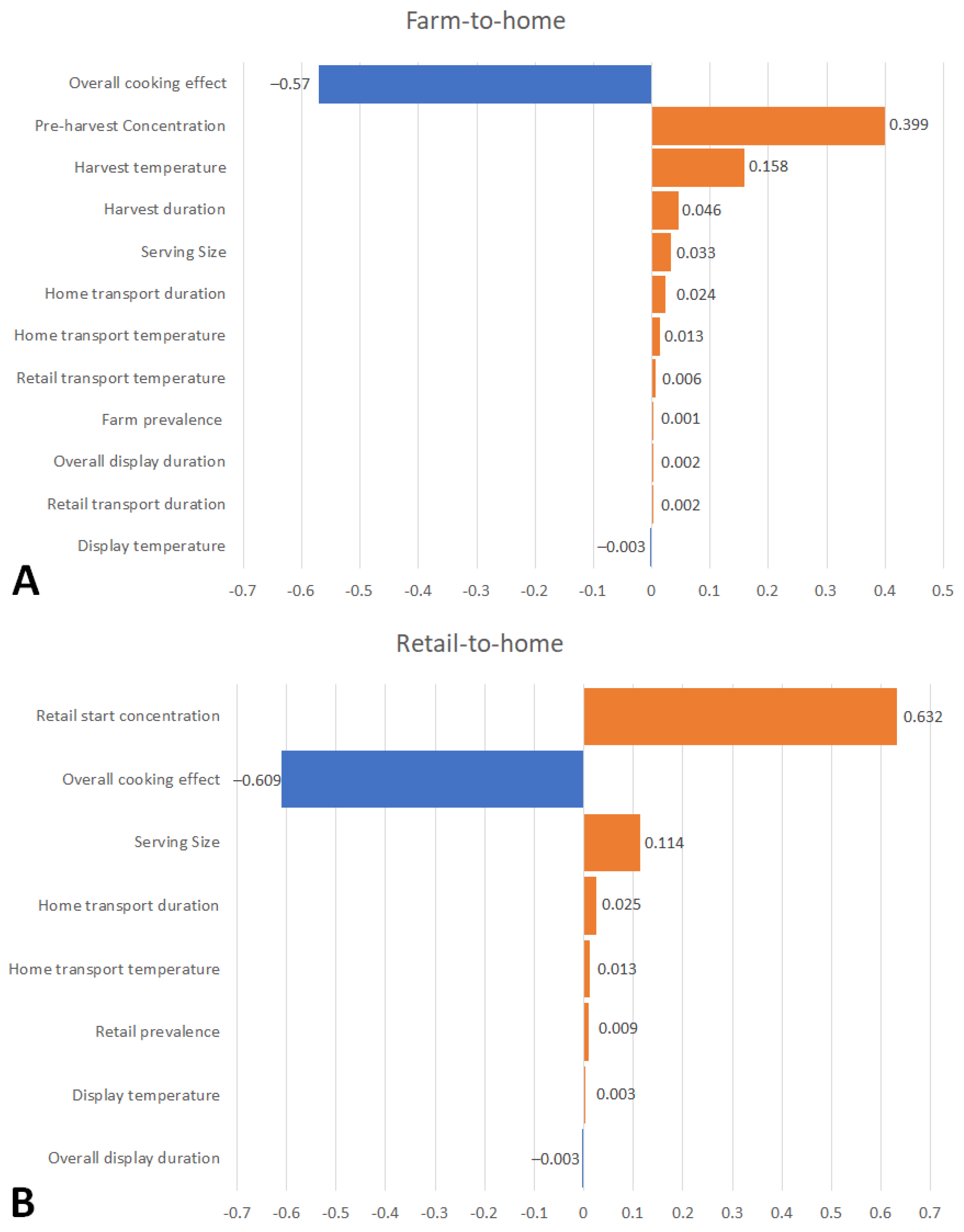
3.3. Sensitivity Analysis
4. Discussion
4.1. Occurrence and Concentration Trends of Vibrio parahaemolyticus in Green Mussels
4.2. Comparative Analysis of ARRA Risk Estimate Outputs to Other Studies
4.3. Comparison Analysis among ARRAs
4.4. Sensitivity Analysis of Intervention Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fujino, T.; Okuno, Y.; Nakada, D.; Aoyama, A.; Fukai, K.; Mukai, T.; Ueho, T. On the bacteriological examination of shirasu-food poisoning. Med. J. Osaka Univ. 1953, 4, 299–304. [Google Scholar]
- Shimohata, T.; Takahashi, A. Diarrhea induced by infection Of Vibrio parahaemolyticus. J. Med. Investig. 2010, 57, 179–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Urtaza, J.; Lozano-Leon, A.; DePaola, A.; Ishibashi, M.; Shimada, K.; Nishibuchi, M.; Liebana, E. Characterization of pathogenic Vibrio parahaemolyticus isolates from clinical sources in Spain and comparison with Asian and North American pandemic isolates. J. Clin. Microbiol. 2004, 42, 4672–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, G.B.; Ramamurthy, T.; Bhattacharya, S.K.; Dutta, B.; Takeda, Y.; Sack, D.A. Global dissemination of Vibrio parahaemolyticus serotype O3:K6 and its serovariants. Clin. Microbiol. Rev. 2007, 20, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Ni, Y.X.; Miwatani, T. Purification and characterization of a hemolysin produced by a clinical isolate of Kanagawa phenomenon-negative Vibrio parahaemolyticus and related to the thermostable direct hemolysin. Infect. Immun. 1988, 56, 961–965. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Kato, T.; Obara, Y.; Akiyama, S. In vitro hemolytic characteristic of Vibrio parahaemolyticus: Its close correlation with human pathogenicity. J. Bacteriol. 1969, 100, 1147–1149. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, S. Sixty years from the discovery of Vibrio parahaemolyticus and some recollections. Biocontrol Sci. 2011, 16, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Twedt, R.M.; Novelli, R.E.; Spaulding, P.L.; Hall, H.E. Comparative hemolytic activity of Vibrio parahaemolyticus and related Vibrios. Infect. Immun. 1970, 1, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D. Vibrio vulnificus. In The Biology of Vibrios; Thompson, F.L., Austin, B., Swings, J., Eds.; ASM Press: Washington, DC, USA, 2006; pp. 349–366. [Google Scholar]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Mazel, D.; Davies, J. Antibiotic resistance in microbes. Cell. Mol. Life Sci. 1999, 56, 742–754. [Google Scholar] [CrossRef] [PubMed]
- De Melo, L.M.; Almeida, D.; Hofer, E.; Dos Reis, C.M.; Theophilo, G.N.; Santos, A.; Vieira, R.H. Antibiotic resistance of Vibrio parahaemolyticus isolated from pond-reared Litopenaeus vannamei marketed in Natal, Brazil. Braz. J. Microbiol. 2011, 42, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food. Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, F.; Walker, R.D.; Janes, M.E.; Prinyawiwatkul, W.; Ge, B. Antimicrobial susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus isolates from Louisiana Gulf and retail raw oysters. Appl. Environ. Microbiol. 2007, 73, 7096–7098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, W.S.; Leisner, J.J.; Rusul, G.; Radu, S.; Rassip, A. Survival of Vibrio spp. including inoculated V. cholerae 0139 during heat-treatment of cockles (Anadara granosa). Int. J. Food. Microbiol. 1998, 42, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Huang, Y.; Chen, H. Inactivation of Vibrio parahaemolyticus and Vibrio vulnificus in oysters by high-hydrostatic pressure and mild heat. Food. Microbiol. 2012, 32, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.; Gennari, M.; Ghidini, V.; Trento, I.; Manfrin, A.; Borrego, J.J.; Lleo, M.M. High incidence of antibiotic multi-resistant bacteria in coastal areas dedicated to fish farming. Mar. Pollut. Bull. 2013, 70, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhang, Y.; Jin, D.; Ding, G.; Luo, Y.; Zhang, J.; Mie, L.; Zhu, M. Molecular characterization and antibiotic susceptibility of Vibrio vulnificus in retail shrimps in Hangzhou, People’s Republic of China. J. Food Prot. 2013, 76, 2063–2068. [Google Scholar] [CrossRef]
- Chua, F.H.C. Aquaculture health management in Singapore: Current status and future directions. In Health Management in Asian Aquaculture; Subasinghe, R.P., Arthur, J.R., Shariff, M., Eds.; FAO: Rome, Italy, 1996; pp. 115–126. [Google Scholar]
- Kruse, H.; Sørum, H. Transfer of multiresistance plasmids between bacteria of diverse origin in natural micro-environments. Appl. Environ. Microbiol. 1994, 60, 4015–4021. [Google Scholar] [CrossRef] [Green Version]
- Ludher, E. Food and the City: Overcoming Challenges for Food Security; Center for Liveable Cities Publications: Singapore, 2018. [Google Scholar]
- Shen, Y.; Ma, K.; Yue, G.H. Status, challenges and trends of aquaculture in Singapore. Aquaculture 2021, 533, 736210. [Google Scholar] [CrossRef]
- FAO; WHO. Part I- Microbiological risk assessment of Vibrio parahaemolyticus in raw oysters. In Risk Assessment of Vibrio Parahaemolyticus in Seafood; Interpretative Summary and Technical Report; FAO: Rome, Italy, 2011; 193p. [Google Scholar]
- FAO; WHO. Part II—Microbiological risk assessment of Vibrio parahaemolyticus in Andara granosa (bloody clam). In Risk Assessment of Vibrio Parahaemolyticus in Seafood; Interpretative Summary and Technical Report; FAO: Rome, Italy, 2011; 193p. [Google Scholar]
- Food and Drug Administration (US FDA). Quantitative Risk Assessment on the Public Health Impact of Pathogenic Vibrio parahaemolyticus in Raw Oysters. [WWW Document]. FDA. 2005. Available online: http://www.fda.gov/downloads/Food/FoodScienceResearch/UCM196915.pdf/ (accessed on 14 October 2021).
- Malcolm, T.T.H.; Cheah, Y.K.; Radzi, C.W.J.W.M.; Kasim, F.A.; Kantilal, H.K.; John, T.Y.H.; Martinez-Urtaza, J.; Nakaguchi, Y.; Nishibuchi, M.; Son, R. Detection and quantification of pathogenic Vibrio parahaemolyticus in shellfish by using multiplex PCR and loop-mediated isothermal amplification assay. Food Control 2015, 47, 664–671. [Google Scholar] [CrossRef]
- Sani, N.A.; Ariyawansa, S.; Babji, A.S.; Hashim, J.K. The risk assessment of Vibrio parahaemolyticus in cooked black tiger shrimps (Penaeus mondon) in Malaysia. Food Control 2013, 31, 546–552. [Google Scholar] [CrossRef]
- Sobrinho, P.D.S.C.; Destro, M.T.; Franco, B.D.G.M.; Landgraf, M. A quantitative risk assessment model for Vibrio parahaemolyticus in raw oysters in Sao Paulo State, Brazil. Int. J. Food Microbiol. 2014, 180, 69–77. [Google Scholar] [CrossRef]
- Yamamoto, A.; Vuddhakul, V.; Charernjiratragul, W.; Vose, D.; Osaka, K.; Shigematsu, M.; Toyofuku, H.; Yamamoto, S.; Nishibuchi, M. Quantitative modeling for risk assessment of Vibrio parahaemolyticus in bloody clams in southern Thailand. Int. J. Food Microbiol. 2008, 124, 70–78. [Google Scholar] [CrossRef]
- Notermans, S.; Mead, G.C. Incorporation of elements of quantitative risk analysis in the HACCP system. Int. J. Food Microbiol. 1996, 30, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Snary, E.L.; Kelly, L.A.; Davison, H.C.; Teale, C.J.; Woolridge, M. Antimicrobial resistance: A microbial risk assessment perspective. J. Antimicrob. Chemother. 2004, 53, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Pichler, M.; Coskun, O.K.; Ortega-Arbulu, A.-S.; Conci, N.; Worheide, G.; Vargas, S.; Orsi, W.D. A 16s rRMA sequencing and analysis protocol for the illumina MiniSeq platform. Microbiologyopen 2018, 7, e00611. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Proposed Guideline, CLSI Document M45-P; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2005. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Codex Alimentarius. Guidelines for risk analysis of foodborne antimicrobial resistance. CAC-GL 2011, 77, 1–29. [Google Scholar]
- Miles, D.W.; Ross, T.; Olley, J.; McMeekin, T.A. Development and evaluation of a predictive model for the effect of temperature and water activity on the growth rate of Vibrio parahaemolyticus. Int. J. Food Microbiol. 1997, 38, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.M.G.; Zhong, Y.; Hu, C.C.; Ong, K.H.; Khor, W.C.; Schlundt, J.; Aung, K.T. Quantitative Risk Evaluation of Antimicrobial-Resistant Vibrio parahaemolyticus Isolated from Farmed Grey Mullets in Singapore. Pathogens 2023, 12, 93. [Google Scholar] [CrossRef]
- Meteorological Service Singapore (MSS). Annual Climate Assessment Reports. [WWW Document]. MSS. 2019. Available online: http://www.weather.gov.sg/wp-content/uploads/2020/03/Annual-Climate-Assessment-Report-2019.pdf/ (accessed on 14 October 2021).
- Iwahori, J.; Yamamoto, A.; Suzuki, H.; Yamamoto, T.; Tsutsui, T.; Motoyama, K.; Sawada, M.; Matsushita, T.; Hasegawa, A.; Osaka, K.; et al. Quantitative risk assessment of Vibrio parahaemolyticus in finfish: A model of raw horse mackerel consumption in Japan. Risk. Anal. 2010, 30, 1817–1832. [Google Scholar] [CrossRef]
- Jouhara, H.; Nannou, T.; Ghazal, H.; Kayyali, R.; Tassou, S.A.; Lester, S. Temperature and energy performance of open refrigerated display cabinets using heat pipe shelves. Energy Procedia 2017, 123, 273–280. [Google Scholar] [CrossRef]
- Tan, C.W.; Malcolm, T.T.H.; Premarathne, J.M.K.J.K.; New, C.Y.; Kuan, C.H.; Thung, T.Y.; Chang, W.S.; Loo, Y.Y.; Rukayadi, Y.; Nakaguchi, Y.; et al. Preliminary quantitative microbial risk assessment of pathogenic Vibrio parahaemolyticus in short mackerel in Malaysia. Microb. Risk. Anal. 2019, 12, 11–19. [Google Scholar] [CrossRef]
- Malcolm, T.T.H.; Cheah, Y.K.; Radzi, C.W.J.W.M.; Kantilal, H.K.; Martinez-Urtaza, J.; Nishibuchi, M.; Son, R. Microbial risk assessment of Vibrio parahaemolyticus in bloody clams in Malaysia: A preliminary model from retail to consumption. Microb. Risk. Anal. 2016, 4, 43–51. [Google Scholar] [CrossRef]
- Health Promotion Board (HPB). National Nutrition Survey 2010 Singapore. [WWW Document]. HPB. 2010. Available online: https://www.hpb.gov.sg/docs/default-source/pdf/nns-2010-report.pdf?sfvrsn=18e3f172_2 (accessed on 14 October 2021).
- Singapore Department of Statistics (SDS). Population and Population Structure. [WWW Document]. SDS. 2019. Available online: https://www.singstat.gov.sg/find-data/search-by-theme/population/population-and-population-structure/latest-data (accessed on 14 October 2021).
- Burger, J.; Fleischer, J.; Gochfeld, M. Fish, shellfish, and meat meals of the public in Singapore. Environ. Res. 2003, 92, 254–261. [Google Scholar] [CrossRef]
- United States Food and Drug Administration (US FDA). Refrigerator thermometers: Cold Facts about Food Safety. [WWW Document]. FDA. 2017. Available online: https://www.fda.gov/food/resourcesforyou/consumers/ucm253954.htm (accessed on 8 October 2021).
- Bej, A.K.; Patterson, D.P.; Brasher, C.W.; Vickery, M.C.; Jones, D.D.; Kaysner, C.A. Detection of total and hemolysin-producing Vibrio parahaemolyticus in shellfish using multiplex PCR amplification of tl, tdh and trh. J. Microbiol. Methods 1999, 36, 215–225. [Google Scholar] [CrossRef]
- Allam, B.; Pales-Espinosa, E. Bivalve immunity and response to infections: Are we looking at the right place? Fish Shellfish Immunol. 2016, 53, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; Gomez-Chiarri, M.; Castillo, M.G.; Figueras, A.; Fiorito, G.; Moreira, R.; Novoa, B.; Pallavicini, A.; Ponte, G.; Roumbedakis, K.; et al. Immunity in Molluscs: Recognition and effector mechanisms, with a focus on Bivalvia. In Advances in Comparative Immunology; Cooper, E., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 225–341. [Google Scholar]
- Boonyawantang, A.; Mahakarnchanakul, W.; Rachtanapun, C.; Boonsupthip, W. Behavior of pathogenic Vibrio parahaemolyticus in prawn in response to temperature in laboratory and factory. Food Control 2012, 26, 479–485. [Google Scholar] [CrossRef]
- Desmarchelier, P.M. Pathogenic vibrios. In Foodborne Microorganisms of Public Health Importance; Hocking, A.D., Arnold, G., Jenson, I., Newton, K., Sutherland, P., Eds.; AIFST (NSW Branch): Sydney, Australia, 1997; pp. 285–312. [Google Scholar]
- Dupray, E.; Cormier, M. Optimal enrichment time for 537 isolation of Vibrio parahaemolyticus from seafood. Appl. Environ. Microbiol. 1983, 46, 1234–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, S.G.; Bayen, S.; Burger, D.; Kellym, B.C.; Han, P.; Babovic, V.; Gin, K.Y. Occurrence and distribution of bacteria indicators, chemical tracers and pathogenic Vibrios in Singapore coastal waters. Mar. Pollut. Bull. 2017, 114, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Bilung, L.M.; Radu, S.; Bahaman, A.R.; Rahim, R.A.; Napis, S.; Wong, C.V.L.M.; Tanil, G.B.; Nishibuchi, M. Detection of Vibrio parahaemolyticus in cockle (Anadara granosa) by PCR. FEMS Microbiol. Lett. 2005, 252, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Gurpreet, K.; Tee, G.H.; Amal, N.M.; Paramesarvathy, R.; Karuthan, C. Incidence and determinants of acute diarrhoea in Malaysia: A population-based study. J. Health Popul. Nutr. 2011, 29, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, A.E.I.; Verhoeff-Bakkenes, L.; Nauta, M.J.; de Jonge, R. Cross-contamination in the kitchen: Effect of hygiene measures. J. Appl. Microbiol. 2008, 105, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collineau, L.; Boerlin, P.; Carson, C.A.; Chapman, B.; Fazil, A.; Hetman, B.; McEwen, S.A.; Parmley, E.J.; Reid-Smith, R.; Taboada, E.N.; et al. Integrating whole-genome sequencing data into quantitative risk assessment of foodborne antimicrobial resistance: A review of opportunities and challenges. Front. Microbiol. 2019, 10, 1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collineau, L.; Chapman, B.; Bao, X.; Sivapathasundaram, B.; Carson, C.A.; Fazil, A.; Reid-Smith, R.J.; Smith, B.A. A farm-to-fork quantitative risk assessment model for Salmonella Heidelberg resistant to third-generation cephalosporins in broiler chickens in Canada. Int. J. Food Microbiol. 2020, 330, 108559. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Equation | References |
|---|---|---|---|
| Exposure assessment | |||
| Growth rate equations | |||
| Growth rate in broth model (Log10/min) | [36] | ||
| Growth rate (Log10/h) | - | ||
| Growth rate adjustment | [25] | ||
| Initial occurrence and concentration equations | |||
| Occurrence of haemolytic Vp | RiskBeta (positives + 1, negatives + 1) | [37] | |
| # | Total concentration of Vp in shellfish (Log10/g) | - | |
| Growth during harvest | |||
| Harvest time (h) | RiskTriang (7, 7.5, 8) | Author’s input | |
| Harvest temperature (K) | RiskPert (299.05, 301.55, 305.45) | [38] | |
| Concentration of Vp (Log10/g) | - | ||
| Growth during transport to retail | |||
| Transport time (h) | RiskUniform (13.5, 14.5) | Author’s input | |
| Transport temperature (K) | RiskPert (276.15, 279.15, 282.15) | [39] | |
| Concentration of Vp | - | ||
| Growth during retail display | |||
| Display time for majority of purchases (h) | RiskUniform (0.5, 3.5) | Author’s input | |
| Display time for remaining purchases (h) | RiskPert (3.5, 5.5, 12.5) | Author’s input | |
| Overall display time (h) | Author’s input | ||
| Display temperature (K) | RiskPert (275.85, 278.15, 279.15) | [40] | |
| Concentration of Vp (Log10/g) | - | ||
| Growth during transport to home | |||
| Transport time (h) | RiskTriang (0.25, 0.5, 0.75) | [41] | |
| Transport temperature (K) | RiskPert (299.05, 301.55, 305.45) | [38] | |
| Concentration of Vp (Log10/g) | - | ||
| Preparation and cooking | |||
| Minimally cooked scenario (Log10/g) | RiskUniform (0, −2) | [15,42] | |
| Moderately cooked scenario (Log10/g) | RiskUniform (−2,−5) | [42] | |
| Highly cooked scenario (Log10/g) | RiskUniform (−5,−7) | [16] | |
| Overall cooked scenario (Log10/g) | Cookingminimal −2.5% Cookingmoderate −2.5% Cookinghighly −95% | [24] | |
| & | Concentration of Vp (Log10/g) | - | |
| Serving Size (grams) | RiskTriang (0, 17.2, 90.47) | [43] | |
| Dose (CFU) | - | ||
| Hazard characterisation | |||
| Beta Poisson dose-response | [25] | ||
| @ | Occurrence change from farm to retail | - | |
| Probability of illness per serving | @ | - | |
| Risk characterisation | |||
| Singapore’s population | 5,703,569 | [44] | |
| Population proportion consuming shellfish | 0.937 | [45] | |
| Population proportion consuming green mussel | Author’s input | ||
| Exposed population | - | ||
| Number of meals per week | RiskNormal (8.29, 8.323) | [45] | |
| Probability of illness per person per year | - | ||
| Cases per year | - | ||
| FDA Model | α | β | Probability Weight |
|---|---|---|---|
| 1 | 1.47 × 106 | 3.53 × 1014 | 3.40 × 10−4 |
| 2 | 1.26 × 107 | 7.20 × 1014 | 4.12 × 10−3 |
| 3 | 6.37 × 102 | 1.65 × 1010 | 2.06 × 10−2 |
| 4 | 3.58 × 101 | 5.42 × 108 | 5.49 × 10−2 |
| 5 | 2.08 × 101 | 1.99 × 108 | 8.23 × 10−2 |
| 6 | 1.49 × 101 | 8.78 × 107 | 6.58 × 10−2 |
| 7 | 1.06 × 101 | 2.99 × 107 | 2.20 × 10−2 |
| 8 | 3.89 | 2.28 × 108 | 6.90 × 10−4 |
| 9 | 1.31 | 2.93 × 107 | 8.23 × 10−3 |
| 10 | 5.20 × 10−1 | 3.61 × 106 | 4.12 × 10−2 |
| 11 | 4.70 × 10−1 | 1.50 × 106 | 1.10 × 10−1 |
| 12 | 6.00 × 10−1 | 1.31 × 106 | 1.65 × 10−1 |
| 13 | 1.00 | 1.80 × 106 | 1.32 × 10−1 |
| 14 | 8.59 | 1.30 × 107 | 4.39 × 10−2 |
| 15 | 1.50 × 10−1 | 2.33 × 105 | 3.40 × 10−4 |
| 16 | 1.90 × 10−1 | 2.29 × 105 | 4.12 × 10−3 |
| 17 | 2.50 × 10−1 | 2.36 × 105 | 2.06 × 10−2 |
| 18 | 3.20 × 10−1 | 2.57 × 105 | 5.49 × 10−2 |
| 19 | 4.30 × 10−1 | 3.04 × 105 | 8.23 × 10−2 |
| 20 | 6.90 × 10−1 | 4.34 × 105 | 6.58 × 10−2 |
| 21 | 6.92 | 4.49 × 106 | 2.20 × 10−2 |
| VP | Marine Coastal Farm | Hypermarket | Farm Water | ||
|---|---|---|---|---|---|
| Sample Total | Meat | Sample Total | Meat | ||
| Occurrence: haemolytic | 45 | 31/45 (69) | 45 | 41/45 (91) | 6/6 (100) |
| Mean concentration: haemolytic | - | 3.0 (0.10) | - | 5.9 (0.12) | 2.8 (0.17) |
| Occurrence: AMP-R | 45 | 14/45 (31) | 45 | 30/45 (67) | 4/6 (67) |
| Mean concentration: AMP-R | - | 2.8 (0.13) | - | 5.3 (0.13) | 2.7 (0.25) |
| Occurrence; PENG-R | 45 | 15/45 (33) | 45 | 34/45 (76) | 5/6 (83) |
| Mean concentration: PENG-R | - | 2.7 (0.17) | - | 4.8 (0.11) | 2.3 (0.34) |
| Occurrence: TET-R | 45 | 25/45 (56) | 45 | 40/45 (89) | 5/6 (83) |
| Mean concentration: TET-R | - | 1.6 (0.081) | - | 2.5 (0.075) | 1.0 (0.075) |
| Farm-To-Home | Retail-To-Home | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Haemolytic | Ampicillin | Penicillin G | Tetracycline | Haemolytic | Ampicillin | Penicillin G | Tetracycline | |||
| Occurrence (Farm/Retail) | 6.8 × 10−1 (5.7 × 10−1, 7.9 × 10−1) | 3.2 × 10−1 (2.1 × 10−1, 4.3 × 10−1) | 3.4 × 10−1 (2.3 × 10−1, 4.6 × 10−1) | 5.5 × 10−1 (4.3 × 10−1, 6.7 × 10−1) | 8.9 × 10−1 (8.1 × 10−1, 9.6 × 10−1) | 6.6 × 10−1 (5.4 × 10−1, 7.7 × 10−1) | 7.5 × 10−1 (6.4 × 10−1, 8.4 × 10−1) | 8.7 × 10−1 (7.9 × 10−1, 9.4 × 10−1) | ||
| Concentration (LogCFU/g) | Farm | Pre-harvest | 3.0 (2.1, 3.9) | 2.8 (2.0, 3.6) | 2.7 (1.6, 3.8) | 1.60 (9.3 × 10−1, 2.3) | * N.A. | N.A. | N.A. | N.A. |
| Post-harvest | 5.1 (4.1, 6.2) | 4.9 (4.0, 5.9) | 4.8 (3.6, 6.0) | 3.7 (2.9, 4.5) | N.A. | N.A. | N.A. | N.A. | ||
| Retail | Retail-start | 5.1 (4.1, 6.2) | 5.0 (4.0, 5.9) | 4.8 (3.7, 6.0) | 3.7 (2.9, 4.6) | 5.9 (4.6, 7.1) | 5.3 (4.1, 6.5) | 4.8 (3.8, 5.9) | 2.5 (1.7, 3.3) | |
| Retail-end | 5.2 (4.1, 6.2) | 5.0 (4.0, 5.9) | 4.8 (3.7, 6.0) | 3.7 (2.9, 4.6) | 5.9 (4.6, 7.1) | 5.3 (4.2, 6.5) | 4.8 (3.8, 5.9) | 2.5 (1.7, 3.3) | ||
| Home | 5.3 (4.2, 6.3) | 5.1 (4.1, 6.1) | 5.0 (3.8, 6.1) | 3.8 (3.0, 4.7) | 6.0 (4.7, 7.3) | 5.5 (4.3, 6.7) | 5.0 (3.9, 6.0) | 2.6 (1.8, 3.4) | ||
| Cooking | Average | 2.5 × 10−1 (0, 1.3) | 1.9 × 10−1 (0, 1.0) | 1.9 × 10−1 (0, 1.1) | 8.7 × 10−2 (0, 0) | 5.6 × 10−1 (0, 2.2) | 3.2 × 10−1 (0, 1.6) | 1.8 × 10−1 (0, 9.7 × 10−1) | 4.3 × 10−2 (0, 0) | |
| Minimally cooked | 4.3 (2.9, 5.7) | 4.1 (2.7, 5.4) | 4.0 (2.5, 5.5) | 2.8 (1.6, 4.1) | 5.0 (3.4, 6.6) | 4.5 (3.0, 6.0) | 4.0 (2.6, 5.4) | 1.6 (3.9 × 10−1, 2.8) | ||
| Moderately cooked | 1.8 (4.1 × 10−2,, 3.5) | 1.6 (0, 3.3) | 1.5 (0, 3.3) | 6.1 × 10−1 (0, 2.0) | 2.5 (6.2 × 10−1, 4.4) | 2.0 (1.4 × 10−1, 3.8) | 1.5 (0, 3.2) | 9.9 × 10−2 (0, 7.0 × 10−1) | ||
| Highly cooked | 1.0 × 10−1 (0, 7.0 × 10−1) | 5.5 × 10−2 (0, 4.4 × 10−1) | 5.9 × 10−2 (0, 4.7 × 10−1) | 2.4 × 10−4 (0, 0) | 4.0 × 10−1 (0, 1.6) | 1.7 × 10−1 (0, 1.0) | 4.9 × 10−2 (0, 4.0 × 10−1) | 0 (0, 0) | ||
| Farm-To-Home | Retail-To-Home | ||||||
|---|---|---|---|---|---|---|---|
| Pill,serving | Pill,yearly | Ncases | Pill,serving | Pill,yearly | Ncases | ||
| Haemolytic | Average | 5.7 × 10−3 (0, 2.9 × 10−4) | 3.4 × 10−2 (0, 9.2 × 10−2) | 1.7 × 102 (0, 4.4 × 102) | 1.3 × 10−2 (0, 2.7 × 10−3) | 7.4 × 10−2 (0, 5.9 × 10−1) | 3.6 × 102 (0, 2.8 × 103) |
| Minimally cooked | 2.2 × 10−1 (3.5 × 10−3, 6.6 × 10−1 ) | 8.0 × 10−1 (0, 1) | 3.9 × 103 (0, 4.8 × 103) | 4.5 × 10−1 (1.6 × 10−2, 8.9 × 10−1) | 8.3 × 10−1 (0, 1) | 4.0 × 103 (0, 4.8 × 103) | |
| Moderately cooked | 9.2 × 10−3 (1.6 × 10−6, 4.4 × 10−2) | 3.3 × 10−1 (0, 1) | 1.6 × 103 (0, 4.8 × 103) | 4.4 × 10−2 (2.6 × 10−5, 2.6 × 10−1) | 5.1 × 10−1 (0, 1) | 2.4 × 103 (0, 4.8 × 103) | |
| Highly cooked | 1.8 × 10−5 (0, 7.5 × 10−5) | 6.7 × 10−3 (0, 3.0 × 10−2) | 3.2 × 101 (0, 1.4 × 102) | 2.0 × 10−4 (0, 6.7 × 10−4) | 4.3 × 10−2 (0, 2.5 × 10−1) | 2.1 × 102 (0, 1.2 × 103) | |
| Haemolytic and AMP-R | Average | 3.4 × 10−3 (0, 1.2 × 10−4) | 2.8 × 10−2 (0, 3.9 × 10−2) | 1.4 × 102 (0, 1.9 × 102) | 6.1 × 10−3 (0, 5.0 × 10−4) | 4.0 × 10−2 (0, 1.5 × 10−1) | 1.9 × 102 (0, 7.2 × 102) |
| Minimally cooked | 1.3 × 10−1 (1.9 × 10−3, 4.5 × 10−1) | 7.8 × 10−1 (0, 1) | 3.8 × 103 (0, 4.8 × 103) | 2.3 × 10−1 (3.9 × 10−3, 6.2 × 10−1) | 8.1 × 10−1 (0, 1) | 3.9 × 103 (0, 4.8 × 103) | |
| Moderately cooked | 4.2 × 10−3 (0, 1.9 × 10−2) | 2.5 × 10−1 (0, 1) | 1.2 × 103 (0, 4.8 × 103) | 1.3 × 10−2 (3.6 × 10−6, 6.8 × 10−2) | 3.6 × 10−1 (0, 1) | 1.7 × 103 (0, 4.8 × 103) | |
| Highly cooked | 6.2 × 10−6 (0, 2.8 × 10−5) | 2.6 × 10−3 (0, 9.9 × 10−3) | 1.3 × 101 (0, 4.8 × 101) | 3.4 × 10−5 (0, 1.3 × 10−4) | 1.1 × 10−2 (0, 5.2 × 10−2) | 5.5 × 101 (0, 2.5 × 102) | |
| Haemolytic and PENG-R | Average | 3.5 × 10−3 (0, 1.4 × 10−4) | 2.8 × 10−2 (0, 4.4 × 10−2) | 1.4 × 102 (0, 2.1 × 102) | 4.1 × 10−3 (0, 1.4 × 10−4) | 2.9 × 10−2 (0, 4.5 × 10−2) | 1.4 × 102 (0, 2.2 × 102) |
| Minimally cooked | 1.3 × 10−1 (1.1 × 10−3, 4.9 × 10−1) | 7.6 × 10−1 (0, 1) | 3.7 × 103 (0, 4.8 × 103) | 1.6 × 10−1 (1.7 × 10−3, 5.8 × 10−1) | 7.8 × 10−1 (0, 1) | 3.7 × 103 (0, 4.8 × 103) | |
| Moderately cooked | 4.8 × 10−3 (0, 2.1 × 10−2) | 2.4 × 10−1 (0, 1) | 1.2 × 103 (0, 4.8 × 103) | 5.1 × 10−3 (0, 2.3 × 10−2) | 2.6 × 10−1 (0, 1) | 1.2 × 103 (0, 4.8 × 103) | |
| Highly cooked | 8.5 × 10−6 (0, 3.2 × 10−5) | 3.3 × 10−3 (0, 1.1 × 10−2) | 1.6 × 101 (0, 5.3 × 101) | 7.6 × 10−6 (0, 3.2 × 10−5) | 3.1 × 10−3 (0, 1.0 × 10−2) | 1.5 × 101 (0, 5.1 × 101) | |
| Haemolytic and TET-R | Average | 7.2 × 10−4 (0, 0) | 1.7 × 10−2 (0, 0) | 7.9 × 101 (0, 0) | 7.2 × 10−5 (0, 0) | 7.1 × 10−3 (0, 0) | 3.4 × 101 (0, 0) |
| Minimally cooked | 2.8 × 10−2 (1.6 × 10−4, 1.4 × 10−1) | 5.9 × 10−1 (0, 1) | 2.8 × 103 (0, 4.8 × 103) | 2.8 × 10−3 (1.1 × 10−5, 1.3 × 10−2) | 2.7 × 10−1 (0, 1) | 1.3 × 103 (0, 4.8 × 103) | |
| Moderately cooked | 3.1 × 10−4 (0, 1.3 × 10−3) | 6.6 × 10−2 (0, 4.2 × 10−1) | 3.2 × 102 (0, 2.0 × 103) | 1.8 × 10−5 (0, 8.5 × 10−5) | 7.1 (0, 3.4 × 10−2) | 3.4 × 101 (0, 1.6 × 102) | |
| Highly cooked | 3.4 × 10−8 (0, 0) | 1.5 × 10−5 (0, 0) | 7.3 × 10−2 (0, 0) | 0 (0, 0) | 0 (0, 0) | 0 (0, 0) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ong, H.M.G.; Zhong, Y.; Hu, C.; Ong, K.H.; Khor, W.C.; Schlundt, J.; Aung, K.T. Antimicrobial Resistance Risk Assessment of Vibrio parahaemolyticus Isolated from Farmed Green Mussels in Singapore. Microorganisms 2023, 11, 1498. https://doi.org/10.3390/microorganisms11061498
Ong HMG, Zhong Y, Hu C, Ong KH, Khor WC, Schlundt J, Aung KT. Antimicrobial Resistance Risk Assessment of Vibrio parahaemolyticus Isolated from Farmed Green Mussels in Singapore. Microorganisms. 2023; 11(6):1498. https://doi.org/10.3390/microorganisms11061498
Chicago/Turabian StyleOng, Hong Ming Glendon, Yang Zhong, Chengcheng Hu, Kar Hui Ong, Wei Ching Khor, Joergen Schlundt, and Kyaw Thu Aung. 2023. "Antimicrobial Resistance Risk Assessment of Vibrio parahaemolyticus Isolated from Farmed Green Mussels in Singapore" Microorganisms 11, no. 6: 1498. https://doi.org/10.3390/microorganisms11061498