

New Insights into the Role and Therapeutic Potential of Heat Shock Protein 70 in Bovine Viral Diarrhea Virus Infection
Abstract
:1. Introduction
2. Bovine Viral Diarrhea Virus and Its Induced Immunosuppression
2.1. BVDV Infection
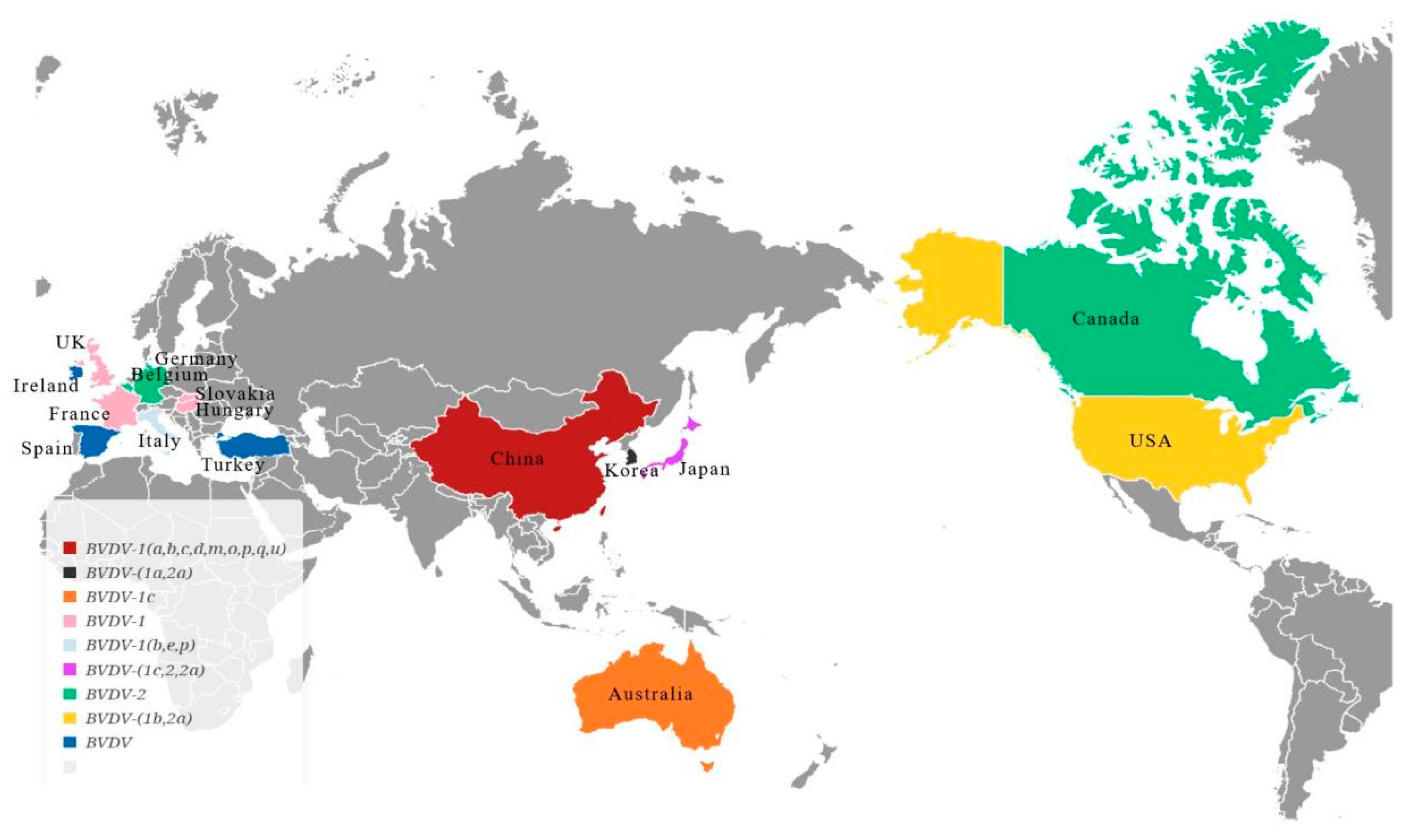
2.2. BVDV Typing and Its Distribution
2.3. BVDV-Induced Immunosuppression in the Body
3. Heat Shock Protein 70 and Its Role in Viral Infection
3.1. Heat Shock Protein 70 Characteristics
3.2. Heat Shock Protein 70’s Role in Viral Replication
3.3. Heat Shock Protein 70 in the Immune Regulation of Antiviral Immunity
3.4. Heat Shock Protein 70 as an Adjuvant for Novel Antiviral Vaccines
4. Role of Heat Shock Protein 70 in Flaviviridae Viruses Infection
4.1. Role of Heat Shock Protein 70 in BVDV Infection
4.2. Role of Heat Shock Protein 70 in ZIKV Infection
4.3. Role of Heat Shock Protein 70 in HCV Infection
4.4. Role of Heat Shock Protein 70 in JEV Infection
4.5. Role of Heat Shock Protein 70 in DENV Infection
4.6. Role of Heat Shock Protein 70 in CSFV Infection
4.7. Role of Heat Shock Protein 70 in WNV Infection
4.8. Role of Heat Shock Protein 70 in EBOV Infection
4.9. Role of Heat Shock Protein 70 in TMUV Infection

{kind=link}
| Virus | In Vitro/In Vivo Study | Mechanism | Reference |
|---|---|---|---|
| BVDV | In vitro | HSP70 promoted BVDV replication in MDBK cells, and the inhibition of HSP70 impeded BVDV replication. The regulation of HSP70 expression has the potential to impact oxidative stress and ERK phosphorylation induced by BVDV. | [25] |
| ZIKV | In vitro | HSP70 mediated ZiKV’s entry, replication, and expulsion from host cells. | [10] |
| In vitro and in vivo | Different cytosolic HSP70 isoforms are recruited to ZIKV-induced compartments and are required for the virus to replicate during pre- and post-entry steps. | [62] | |
| WNV | In vitro | The upregulation of numerous subtypes of HSP70 led to increased viral replication, and HSP70 was found to interact with WNV Cp. | [76] |
| In vitro | HSP70 functions as a negative regulator of the West Nile virus capsid protein through direct interaction. | [77] | |
| HCV | In vitro | The infection of host cells with HCV can increase the expression of HSP70, which can bind to the viral replication complex and facilitate the assembly and replication of viral genes. | [63,64] |
| In vitro | The binding site for NS5A/HSP70 was determined to be a hairpin moiety at the C-terminus of NS5A domain I and showed a corresponding cyclized polyarginine-tagged synthetic peptide (HCV4) that significantly blocks virus production. | [65] | |
| In vitro | HSP70 interacts with NS5A, one of the components of the viral replication complex, thereby affecting the proliferation of the virus. | [66] | |
| CSFV | In vitro | The data suggest that HSP70 is critical in the viral life cycle, particularly during the virus RNA replication period. | [8] |
| In Vivo Study | HSP70 fused with the envelope glycoprotein E0 of the classical swine fever virus, enhancing immune responses in Balb/c mice. | [75] | |
| JEV | In vitro | The upregulation of HSP70 by JEV NS5 plays a dual role by contributing to the antagonism of type I IFN and participating in the anti-apoptotic effect of JEV NS5. This is achieved by preventing p38 MAPK/Stat1-mediated apoptosis induced by IFN-β. | [67] |
| In vitro | The binding of HSP70 to lipid rafts is necessary for JEV infection in Huh7 cells. | [68] | |
| In vitro | JEV usually enters host cells through receptor-mediated clathrin-dependent endocytosis, which requires HSP70/Hsc70 to be composed of at least three subtypes (B, C, and D). | [69] | |
| In vitro | During JEV infection, HSP70 in the cytoplasm colocalized with components of the viral replicase complex (NS3, NS5, and viral dsRNA). Knockout of HSP70 resulted in a significant reduction in JEV genome replication. | [70] | |
| In vitro | Heat shock protein 70, in Neuro2a cells, is a putative receptor of the Japanese encephalitis virus. | [71] | |
| In Vivo Study | Immunizing pigs with HSP70 fusion protein can promote lymphocyte proliferation. | [59] | |
| DENV | In vitro | HSP90 and HSP70 are involved in the entry of the dengue virus as receptor complexes in human cell lines and monocytes/macrophages. | [70] |
| In vitro | HSP70 helps the virus to reproduce by inhibiting the type 1 interferon response. | [70] | |
| In vitro | HSP70/HSc70 is the most critical component of DENV infection. | [72] | |
| In vitro | The use of JHSP70 inhibitors was shown to block dengue virus activity, indicating that HSP70 can be exploited as a host antiviral target. | [73] | |
| EBOV | In vitro | The binding of HSP70 to the EBOV protein promotes its stability. | [78] |
| TMUV | In vitro | HSP70 plays an important role in the TMUV life cycle (virus replication, assembly, and release). The inhibition of HSP70 expression significantly reduces TMUV-induced apoptosis. | [79] |
| Flavivirus | In vitro and in vivo | Small-molecule inhibitors of HSP70 ATPase activity can be used as broad-spectrum inhibitors against flavivirus infection. | [59] |
4.10. Antiviral Effects of the Heat Shock Protein 70 Inhibitor Quercetin
4.10.1. The HSP70 Inhibitor Quercetin in Flaviviridae Virus Infection
| Virus | Mechanism | Reference |
|---|---|---|
| DENV | Quercetin has the best binding energy for NS2B-NS3 protease. | [85] |
| Quercetin interacts with the biological molecules of dengue virus serotype 3 NS2B-NS3 protease to inhibit dengue virus. | [86] | |
| Quercetin showed the most significant preventive effect against DENV infection. In addition, quercetin also showed the strongest binding affinity for the DENV membrane receptor TIM-1 protein in molecular docking analysis. | [87] | |
| Quercetin affects DENV-2 virus replication but does not affect DENV-2 attachment to host cells. | [88] | |
| Quercetin inhibits viral replication in vivo and in vitro in a dose-dependent manner. | [90] | |
| Coinfection wtith COVID-19 and dengue fever | Quercetin inhibits cytokine release through NF-κB, IL-17, and toll-like receptor signaling pathways, alleviates over-immune responses, and eliminates inflammation. | [89] |
| ZIKV | Quercetin prevents the internalization of the virus particles to the host cell, preventing the entry of ZIKV. | [91] |
| Quercetin targets viral replication and almost completely inhibits Zika virus RNA production. | [93] | |
| JEV | Quercetin reduced the JEV RNA copy number. | [92] |
| HCV | Quercetin inhibits the activity of the viral protease non-structural protein 3 (NS3), effectively targeting and reducing HCV RNA and resulting in reduced HCV replication. | [23] |
| Quercetin decreased the production of reactive oxygen and nitrogen species (ROS/RNS) and lipid peroxidation induced by HCV and inhibited HCV replication. Quercetin decreased the increased expression of diglyceride acyltransferase 1 (DGAT1) mRNA after viral infection and inhibited HCV genome replication. Quercetin can affect the morphogenesis of infectious particles and reduce HCV-specific infectivity. The integrity of virions is affected when applied directly to HCV particles, hindering the localization of the HCV core protein in LDs. | [94] | |
| EBOV | Quercetin blocks Ebola virus infection by inhibiting the inhibitory function of VP24 interferon. | [95] |
4.10.2. Quercetin Suppresses HSP70’s Effects in Viral Infection
| Mechanism | Reference | |
|---|---|---|
| BVDV | Quercetin inhibits HSP70 and thus BVDV virus replication (in the early stage of BVDV infection) | [25] |
| CFSV/HCV | Quercetin inhibits viral translation by blocking the induction of the host protein (NS5A) and HSP70 | [8,82] |
| PRRSV | Quercetin inhibits HSP70 and reduces porcine reproductive and respiratory syndrome virus replication in vitro. | [98] |
| JEV | Quercetin-induced downregulation of HSP70 led to a significant recovery of IFN-β-induced apoptosis in NS5-expressing cells. An increase in the phosphorylation of ERK2, p38 MAPK, and STAT1 accompanied this process. | [67] |
| Plant virus | Quercetin inhibits tobacco mosaic virus (TMV) by inhibiting NbHSP70er-1 and NbHSP70c-A and downregulating HSP70 protein expression. | [99] |
| Quercetin can inhibit the proliferation of the potato disease virus. | [99] | |
| Quercetin treatment could downregulate the expression of HSP70 protein in cells and significantly reduce the RNA proliferation of tomato dwarf virus (TBSV). | [100] |
5. Conclusions and Future Perspective
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Efferth, T.; Sauerbrey, A.; Halatsch, M.E.; Ross, D.D.; Gebhart, E. Molecular modes of action of cephalotaxine and homoharringtonine from the coniferous tree Cephalotaxus hainanensis in human tumor cell lines. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 367, 56–67. [Google Scholar] [CrossRef]
- Zeisel, M.B.; Baumert, T.F. Production of infectious hepatitis C virus in tissue culture: A breakthrough for basic and applied research. J. Hepatol. 2006, 44, 436–439. [Google Scholar] [CrossRef]
- Buckwold, V.E.; Beer, B.E.; Donis, R.O. Bovine viral diarrhea virus as a surro-gate model of hepatitis C virus for the evaluation of antiviral agents. Antiviral Res. 2003, 60, 1–15. [Google Scholar] [CrossRef]
- Fautz, R.; Husein, B.; Hechenberger, C. Application of the Neutral Red as-say (NR assay) to monolayer cultures of primary hepatocytes. Rapidcolorimetric viability determination for the unscheduled DNA syn-thesis test (UDS). Mutat. Res. 1991, 235, 173–179. [Google Scholar] [CrossRef]
- Ridpath, J.F. Immunology of BVDV vaccines. Biologicals 2013, 41, 14–19. [Google Scholar] [CrossRef]
- Kregel, K.C. Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186. [Google Scholar] [CrossRef]
- Saini, J.; Sharma, P.K. Clinical, Prognostic and Therapeutic Significance of Heat Shock Proteins in Cancer. Curr. Drug Targets 2018, 19, 1478–1490. [Google Scholar] [CrossRef]
- Zhang, C.; Kang, K.; Ning, P.; Peng, Y.; Lin, Z.; Cui, H.; Cao, Z.; Wang, J.; Zhang, Y. Heat shock protein 70 is associated with CSFV NS5A protein and enhances viral RNA replication. Virology 2015, 482, 9–18. [Google Scholar] [CrossRef]
- Taguwa, S.; Maringer, K.; Li, X.; Bernal-Rubio, D.; Rauch, J.N.; Gestwicki, J.E.; Andino, R.; Fernandez-Sesma, A.; Frydman, J. Defining HSP70 Subnetworks in Dengue Virus Replication Reveals Key Vulnerability in Flavivirus Infection. Cell 2015, 163, 1108–1123. [Google Scholar] [CrossRef]
- Pujhari, S.; Brustolin, M.; Macias, V.M.; Nissly, R.H.; Nomura, M.; Kuchipudi, S.V.; Rasgon, J.L. Heat shock protein 70 (HSP70) mediates Zika virus entry, replication, and egress from host cells. Emerg. Microbes Infect. 2019, 8, 8–16. [Google Scholar] [CrossRef]
- Schweizer, M.; Peterhans, E. Oxidative stress in cells infected with bovine viral diarrhoea virus: A crucial step in the induction of apoptosis. J. Gen. Virol. 1999, 80, 1147–1155. [Google Scholar] [CrossRef]
- Finkielsztein, L.M.; Moltrasio, G.Y.; Caputto, M.E.; Castro, E.F.; Cavallaro, L.V.; Moglioni, A.G. What is known about the antiviral agents active against bovine viral diarrhea virus (BVDV)? Curr. Med. Chem. 2010, 17, 2933–2955. [Google Scholar] [CrossRef]
- Kuca, T.; Passler, T.; Newcomer, B.W.; Neill, J.D.; Galik, P.K.; Riddell, K.P.; Zhang, Y.; Bayles, D.O.; Walz, P.H. Changes Introduced in the Open Reading Frame of Bovine Viral Diarrhea Virus During Serial Infection of Pregnant Swine. Front. Microbiol. 2020, 11, 1138. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, S.; He, B.; Wang, T.; Zhao, S.; Wu, C.; Yue, S.; Zhang, S.; He, M.; Wang, L.; et al. PD-1 blockade inhibits lymphocyte apoptosis and restores proliferation and anti-viral immune functions of lymphocyte after CP and NCP BVDV infection in vitro. Vet. Microbiol. 2018, 226, 74–80. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhou, Y.; Liu, Y. Bovine Viral Diarrhea—Mucosal Disease Control; Harbin Heilongjiang Science and Technology Press: Harbin, China, 2022; Volume 1, p. 8. ISBN 9787571915438. [Google Scholar]
- Di, P.A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and its derivates as antiviral potentials: A comprehensive review. Phytother. Res. 2022, 36, 266–278. [Google Scholar]
- Ridpath, J.F.; Bendfeldt, S.; Neill, J.D.; Liebler-Tenorio, E. Lymphocytopathogenic activity in vitro correlates with high virulence in vivo for BVDV type 2 strains: Criteria for a third biotype of BVDV. Vir. Res. 2006, 118, 62–69. [Google Scholar] [CrossRef]
- Peterhans, E.; Schweizer, M. Pestiviruses: How to outmaneuver your hosts. Vet. Microbiol. 2010, 142, 18–25. [Google Scholar] [CrossRef]
- Villalba, M.; Fredericksen, F.; Otth, C.; Olavarría, V. Transcriptomic analysis of responses to cytopathic bovine viral diarrhea virus-1 (BVDV-1) infection in MDBK cells. Mol. Immunol. 2016, 71, 192–202. [Google Scholar] [CrossRef]
- Song, Q.; Zhao, X.; Cao, C.; Duan, M.; Shao, C.; Jiang, S.; Zhou, B.; Zhou, Y.; Dong, W.; Yang, Y.; et al. Research advances on interferon (IFN) response during BVDV infection. Res. Vet. Sci. 2022, 149, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, C.; van den Hurk, J.; Lecomte, J.; Tijssen, P. Identifcation of a new group of bovine viral diarrhea virus strains associated with severe outbreaks and high mortalities. Virology 1994, 203, 260–268. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Neill, J.D.; Frey, M.; Landgraf, J.G. Phylogenetic, antigenic and clinical characterization of type 2 BVDV from North America. Vet. Microbiol. 2000, 77, 145–155. [Google Scholar] [CrossRef]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral. Hepat. 2012, 19, e81–e88. [Google Scholar] [CrossRef]
- Gangwar, V.; Garg, A.; Lomore, K.; Korla, K.; Bhat, S.S.; Rao, R.P.; Rafiq, M.; Kumawath, R.; Uddagiri, B.V.; Kareenhalli, V.V. Immunomodulatory Effects of a Concoction of Natural Bioactive Compounds-Mechanistic Insights. Biomedicines 2021, 9, 1522. [Google Scholar] [CrossRef]
- Chen, N.; Liu, Y.; Bai, T.; Chen, J.; Zhao, Z.; Li, J.; Shao, B.; Zhang, Z.; Zhou, Y.; Wang, X.; et al. Quercetin Inhibits HSP70 Blocking of Bovine Viral Diarrhea Virus Infection and Replication in the Early Stage of Virus Infection. Viruses 2022, 14, 2365. [Google Scholar] [CrossRef] [PubMed]
- Bachar, S.C.; Mazumder, K.; Bachar, R.; Aktar, A.; Al Mahtab, M. A Review of Medicinal Plants with Antiviral Activity Available in Bangladesh and Mechanistic Insight into Their Bioactive Metabolites on SARS-CoV-2, HIV and HBV. Front. Pharmacol. 2021, 12, 732891. [Google Scholar] [CrossRef]
- Letellier, C.; Kerkhofs, P.; Wellemans, G.; Vanopdenbosch, E. Detection and genotyping of bovine diarrhea virus by reverse transcription-polymerase chain amplifcation of the 5′untranslated region. Vet. Microbiol. 1999, 64, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Luzzago, C.; Lauzi, S.; Ebranati, E.; Giammarioli, M.; Moreno, A.; Cannella, V.; Masoero, L.; Canelli, E.; Guercio, A.; Caruso, C.; et al. Extended genetic diversity of bovine viral diarrhea virus and frequency of genotypes and subtypes in cattle in Italy between 1995 and 2013. Biomed. Res. Int. 2014, 2014, 147145. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, B.W.; Kelling, C.L. Alteration of leukocyte populations in calves concurrently infected with bovine respiratory syncytial virus and bovine viral diarrhea virus. Viral. Immunol. 1999, 12, 323–334. [Google Scholar] [CrossRef]
- Glew, E.J.; Carr, B.V.; Brackenbury, L.S.; Hope, J.C.; Charleston, B.; Howard, C.J. Differential effects of bovine viral diarrhoea virus on monocytes and dendritic cells. J. Gene Virol. 2003, 84, 1771–1780. [Google Scholar] [CrossRef]
- Pedrera, M.; Sánchez-Cordón, P.J.; Romero-Trevejo, J.L.; Risalde, M.A.; Greiser-Wilke, I.; Núñez, A. Morphological Changes and Virus Distribution in the Ileum of Colostrum-Deprived Calves Inoculated with Non-Cytopathic Bovine Viral Diarrhoea Virus Genotype-1. J. Comp. Pathol. 2009, 141, 52–62. [Google Scholar] [PubMed]
- Walz, P.H.; Chamorro, M.F.; Falkenberg, S.; Passler, T.; van der Meer, F.; Woolums, A.R. Bovine viral diarrhea virus: An updated American College of Veterinary Internal Medicine consensus statement with focus on virus biology, hosts, immunosuppression, and vaccination. J. Vet. Intern. Med. 2020, 34, 1690–1706. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Pomp, D.; Donis, R.O. Patterns of cellular gene expression in cells infected with cytopathic or non-cytopathic bovine viral diarrhea virus. Anim. Biotechnol. 2003, 14, 31–49. [Google Scholar]
- Schlesinger, M.J. Heat shock proteins. J. Biol. Chem. 1990, 265, 12111–12114. [Google Scholar] [CrossRef]
- Silver, J.T.; Noble, E.G. Regularion of survival gene HSP70. Cell Stress Chaperones 2012, 17, 1–9. [Google Scholar]
- Li, Z.; Srivastava, P. Heat-shock proteins. Curr. Protoc. Immunol. 2004, 58, A.1T.1–A.1T.6. [Google Scholar]
- Morimoto, R.l.; Sarge, K.D.; Abravaya, K. Transcriptional regulation of heat shock genes. A paradigm for inducible genomic responses. J. Biol. Chem. 1992, 267, 21987–21990. [Google Scholar] [CrossRef]
- Manzoor, R.; Kuroda, K.; Yoshida, R.; Tsuda, Y.; Fujikura, D.; Miyamoto, H.; Kajihara, M.; Kida, H.; Takada, A. Heat shock protein 70 modulates influenza A virus polymerase activity. J. Biol. Chem. 2014, 289, 7599–7614. [Google Scholar]
- Qu, B.; Jia, Y.; Liu, Y.; Wang, H.; Ren, G.; Wang, H. The detection and role of heat shock protein 70 in various nondisease conditions and disease conditions: A literature review. Cell Stress Chaperones 2015, 20, 885–892. [Google Scholar]
- Jindal, S.; Young, R.A. Vaccinia virus infection induces a stress response that leads to association of HSP70 with viral proteins. J. Virol. 1992, 66, 5357–5362. [Google Scholar] [CrossRef]
- Collins, P.L.; Hightower, L.E. Newcastle disease virus stimulates the cellular accumulation of stress (heat shock) mRNAs and proteins. J. Virol. 1982, 44, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Thangue, N.B.; Latchman, D.S. A cellular protein related to heat-shock protein 90 accumulates during herpes simplex virus infection and is overexpressed in transformed cells. Exp. Cell Res. 1988, 178, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.; Abravaya, K.; Morimoto, R.I. Analysis of the specificity and mechanism of transcriptional activation of the human HSP70 gene during infection by DNA viruses. J. Virol. 1991, 65, 5680–5692. [Google Scholar] [CrossRef]
- Jolly, C.; Morimoto, R.I. Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J. Natl. Cancer Inst. 2000, 92, 1564–1572. [Google Scholar] [CrossRef]
- Bolhassani, A.; Agi, E. Heat shock proteins in infection. Clin. Chim. Acta 2019, 498, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Ma, Y.; Zhang, Y.; Li, J.; Shu, Y.; Oglesbee, M. HSP70-dependent antiviral immunity against cytopathic neuronal infection by vesicular stomatitis virus. J. Virol. 2013, 87, 10668–10678. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Shu, Y.; Carsillo, T.; Zhang, J.; Yu, L.; Peterson, C.; Longhi, S.; Girod, S.; Niewiesk, S.; Oglesbee, M. HSP70 and a novel axis of type I interferon-dependent antiviral immunity in the measles virus-infected brain. J. Virol. 2013, 87, 998–1009. [Google Scholar] [CrossRef]
- Dabaghian, M.; Latifi, A.M.; Tebianian, M.; Dabaghian, F.; Ebrahimi, S.M. A truncated C-terminal fragment of Mycobacterium tuberculosis HSP70 enhances cell-mediated immune response and longevity of the total IgG to influenza A virus M2e protein in mice. Antiviral Res. 2015, 120, 23–31. [Google Scholar] [CrossRef]
- Wan, T.; Zhou, X.; Chen, G.; An, H.; Chen, T.; Zhang, W.; Liu, S.; Jiang, Y.; Yang, F.; Wu, Y.; et al. Novel heat shock protein HSP70L1 activates dendritic cells and acts as a Th1 polarizing adjuvant. Blood 2004, 103, 1747–1754. [Google Scholar] [CrossRef]
- Khachatoorian, R.; Ganapathy, E.; Ahmadieh, Y.; Wheatley, N.; Sundberg, C.; Jung, C.L.; Arumugaswami, V.; Raychaudhuri, S.; Dasgupta, A.; French, S.W. The NS5A-binding heat shock proteins HSC70 and HSP70 play distinct roles in the hepatitis C viral life cycle. Virology 2014, 455, 118–127. [Google Scholar] [CrossRef]
- Ho, K.T.; Homma, K.; Takanari, J.; Bai, H.; Kawahara, M.; Nguyen, K.T.K.; Takahashi, M. A standardized extract of Asparagus officinalis stem improves HSP70-mediated redox balance and cell functions in bovine cumulus-granulosa cells. Sci. Rep. 2021, 11, 18175. [Google Scholar] [PubMed]
- Shaul, Y.D.; Seger, R. The MEK/ERK cascade: From signaling specificity to diverse functions. Biochim. Biophys. Acta 2007, 1773, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, M.; Olivier, M. Hydrogen peroxide induces murine macrophage chemokine gene transcription via extracellular signal-regulated kinase-and cyclic adenosine 5′-monophosphate (cAMP)-dependent pathways: Involvement of NF-kappa B, activator protein 1, and cAMP response element binding protein. J. Immunol. 2002, 169, 7026–7038. [Google Scholar] [PubMed]
- Brünnert, D.; Langer, C.; Zimmermann, L.; Bargou, R.C.; Burchardt, M.; Chatterjee, M.; Stope, M.B. The heat shock protein 70 inhibitor VER155008 suppresses the expression of HSP27, HOP and HSP90β and the androgen receptor, induces apoptosis, and attenuates prostate cancer cell growth. J. Cell Biochem. 2020, 121, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Yamane, D.; Zahoor, M.A.; Mohamed, Y.M.; Azab, W.; Kato, K.; Tohya, Y.; Akashi, H. Activation of extracellular signal-regulated kinase in MDBK cells infected with bovine viral diarrhea virus. Arch. Virol. 2009, 154, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kim, D.G.; Kim, S. ERK-dependent phosphorylation of the linker and substrate-binding domain of HSP70 increases folding activity and cell proliferation. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef]
- Qi, Z.; Qi, S.; Gui, L.; Shen, L.; Feng, Z. Daphnetin protects oxidative stress-induced neuronal apoptosis via regulation of MAPK signaling and HSP70 expression. Oncol. Lett. 2016, 12, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.W.; Chen, Y.; Liu, W.; Chen, Y.J.; Fan, Z.M.; Liu, M.; Li, L.J. Inhibition of JAK2/STAT3 Signaling Pathway Suppresses Proliferation of Burkitt’s Lymphoma Raji Cells via Cell Cycle Progression, Apoptosis, and Oxidative Stress by Modulating HSP70. Med. Sci. Monit. 2018, 24, 6255–6263. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, C.; Chen, N.; Li, Y.; Fan, C.; Zhao, S.; Bai, T.; Zhao, Z.; Chen, J.; Su, S.; et al. PD-1 Blockade Restores the Proliferation of Peripheral Blood Lymphocyte and Inhibits Lymphocyte Apoptosis in a BALB/c Mouse Model of cp BVDV Acute Infection. Front. Immunol. 2021, 12, 727254. [Google Scholar] [CrossRef]
- Yang, J.; Xu, Y.; Yan, Y.; Li, W.; Zhao, L.; Dai, Q.; Li, Y.; Li, S.; Zhong, J.; Cao, R.; et al. Small Molecule Inhibitor of ATPase Activity of HSP70 as a Broad-Spectrum Inhibitor against Flavivirus Infections. ACS Infect Dis. 2020, 6, 832–843. [Google Scholar] [CrossRef]
- Ge, F.-f.; Wang, J.; Feng, X.; Sheng, L.P.; Sun, Q.y.; Zhou, J.p.; Chen, P.y.; Liu, P.h. Japanese encephalitis protein vaccine candidates expressing neutralizing epitope and M.T HSP70 induce virus-specific memory B cells and long-lasting antibodies in swine. Vaccine 2008, 26, 5590–5594. [Google Scholar]
- Taguwa, S.; Yeh, M.T.; Rainbolt, T.K.; Nayak, A.; Shao, H.; Gestwicki, J.E.; Andino, R.; Frydman, J. Zika Virus Dependence on Host HSP70 Provides a Protective Strategy against Infection and Disease. Cell Rep. 2019, 26, 906–920.e3. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chen, Y.H.; Chow, L.P.; Tsai, Y.H.; Chen, P.H.; Huang, C.Y.; Chen, W.T.; Hwang, L.H. Heat shock protein 72 is associated with the hepatitis C virus replicase complex and enhances viral RNA replication. J. Biol. Chem. 2010, 285, 28183–28190. [Google Scholar] [CrossRef]
- Parent, R.; Qu, X.; Petit, M.A.; Beretta, L. The heat shock cognate protein 70 is associated with hepatitis C virus particles and modulates virus infectivity. Hepatology 2009, 49, 1798–1809. [Google Scholar] [CrossRef]
- Khachatoorian, R.; Ruchala, P.; Waring, A.; Jung, C.L.; Ganapathy, E.; Wheatley, N.; Sundberg, C.; Arumugaswami, V.; Dasgupta, A.; French, S.W. Structural characterization of the HSP70 interaction domain of the hepatitis C viral protein NS5A. Virology 2015, 475, 46–55. [Google Scholar] [CrossRef]
- Gonzalez, O.; Fontanes, V.; Raychaudhuri, S.; Loo, R.; Loo, J.; Arumugaswami, M.; Sun, R.; Dasgupta, A.; French, S.W. The heat shock protein inhibitor Quercetin attenuates hepatitis C virus production. Hepatology 2009, 50, 1756–1764. [Google Scholar] [CrossRef]
- Weng, J.R.; Hua, C.H.; Chen, C.H.; Huang, S.H.; Wang, C.Y.; Lin, Y.J.; Wan, L.; Lin, C.W. Anti-apoptotic activity of Japanese encephalitis virus NS5 protein in human medulloblastoma cells treated with interferon-β. J. Microbiol. Immunol. Infect. 2018, 51, 456–464. [Google Scholar] [CrossRef]
- Zhu, Y.Z.; Cao, M.M.; Wang, W.B.; Wang, W.; Ren, H.; Zhao, P.; Qi, Z.T. Association of heat-shock protein 70 with lipid rafts is required for Japanese encephalitis virus infection in Huh7 cells. J. Gen. Virol. 2012, 93, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.K.; Yang, T.H.; Chen, T.H.; Yang, C.F.; Chen, W.J. Heat shock cognate protein 70 isoform D is required for clathrin-dependent endocytosis of Japanese encephalitis virus in C6/36 cells. J. Gen. Virol. 2015, 96, 793–803. [Google Scholar] [CrossRef]
- Reyes-del Valle, J.; Chávez-Salinas, S.; Medina, F.; Del Angel, R.M. Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J. Virol. 2005, 79, 4557–4567. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Laxminarayana, S.V.; Chandra, N.; Ravi, V.; Desai, A. Heat shock protein 70 on Neuro2a cells is a putative receptor for Japanese encephalitis virus. Virology 2009, 385, 47–57. [Google Scholar] [CrossRef]
- Vega-Almeida, T.O.; Salas-Benito, M.; Nova-Ocampo, M.A.; Angel, R.M.; Salas-Benito, J.S. Surface proteins of C6/36 cells involved in dengue virus 4 binding and entry. Arch. Virol. 2013, 158, 1189–1207. [Google Scholar] [CrossRef] [PubMed]
- Padwad, Y.S.; Mishra, K.P.; Jain, M.; Chanda, S.; Ganju, L. Dengue virus infection activates cellular chaperone HSP70 in THP-1 cells: Downregulation of HSP70 by siRNA revealed decreased viral replication. Viral. Immunol. 2010, 23, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Howe, M.K.; Speer, B.L.; Hughes, P.F.; Loiselle, D.R.; Vasudevan, S.; Haystead, T.A. An inducible heat shock protein 70 small molecule inhibitor demonstrates anti-dengue virus activity, validating HSP70 as a host antiviral target. Antiviral Res. 2016, 130, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, X.; Jing, J.; Shi, B.; Wang, S.; Zhou, B.; Chen, P. HSP70 Fused with the Envelope Glycoprotein E0 of Classical Swine Fever Virus Enhances Immune Responses in Balb/c Mice. Bing Du Xue Bao 2015, 31, 363–369. (In Chinese) [Google Scholar]
- Pastorino, B.; Boucomont-Chapeaublanc, E.; Peyrefitte, C.N.; Belghazi, M.; Fusaï, T.; Rogier, C.; Tolou, H.J.; Almeras, L. Identification of cellular proteome modifications in response to West Nile virus infection. Mol. Cell Proteomics 2009, 8, 1623–1637. [Google Scholar] [CrossRef]
- Oh, W.K.; Song, J. HSP70 functions as a negative regulator of West Nile virus capsid protein through direct interaction. Biochem. Biophys. Res. Commun. 2006, 347, 994–1000. [Google Scholar] [CrossRef]
- García-Dorival, I.; Wu, W.; Armstrong, S.D.; Barr, J.N.; Carroll, M.W.; Hewson, R.; Hiscox, J.A. Elucidation of the Cellular Interactome of Ebola Virus Nucleoprotein and Identification of Therapeutic Targets. J. Proteome Res. 2016, 15, 4290–4303. [Google Scholar] [CrossRef]
- Dai, G.; Han, K.; Huang, X.; Zhang, L.; Liu, Q.; Yang, J.; Liu, Y.; Li, Y.; Zhao, D. Heat shock protein 70 (HSP70) plays important role in tembusu virus infection. Vet. Microbiol. 2022, 267, 109377. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Yang, J.X.; Maria, T.C.; Zhou, B.; Xiao, F.L.; Wang, M.; Mao, Y.J.; Li, Y. Quercetin improves immune function in Arbor Acre broilers through activation of NF-κB signaling pathway. Poult. Sci. 2020, 99, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Khachatoorian, R.; Arumugaswami, V.; Raychaudhuri, S.; Yeh, G.K.; Maloney, E.M.; Wang, J.; Dasgupta, A.; French, S.W. Divergent antiviral effects of bioflavonoids on the hepatitis C virus life cycle. Virology 2012, 433, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Storniolo, A.; Raciti, M.; Cucina, A.; Bizzarri, M.; Renzo, L. Quercetin affects HSP70/IRE1αmediated protection from death induced by endoplasmic reticulum stress. Oxid. Med. Cell Longev. 2015, 2015, 645157. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, E.J.; Min, K.H.; Hur, G.Y.; Lee, S.H.; Lee, Y.; Kim, J.H.; Shin, C.; Shim, J.J.; In, K.H.; et al. Quercetin Enhances Chemosensitivity to Gemcitabine in Lung Cancer Cells by Inhibiting Heat Shock Protein 70 Expression. Clin Lung Cancer 2015, 16, e235–e243. [Google Scholar] [CrossRef]
- Senthilvel, P.; Lavanya, P.; Kumar, K.M.; Swetha, R.; Anitha, P.; Bag, S.; Sarveswari, S.; Vijayakumar, V.; Ramaiah, S.; Anbarasu, A. Flavonoid from Carica papaya inhibits NS2B-NS3 protease and prevents Dengue 2 viral assembly. Bioinformation 2013, 9, 889–895. [Google Scholar] [CrossRef]
- De Sousa, L.R.; Wu, H.; Nebo, L.; Fernandes, J.B.; Da Silva, M.F.; Kiefer, W.; Kanitz, M.; Bodem, J.; Diederich, W.E.; Schirmeister, T.; et al. Flavonoids as noncompetitive inhibitors of Dengue virus NS2B-NS3 protease: Inhibition kinetics and docking studies. Bioorg. Med. Chem. 2015, 23, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Chakraborty, U.; Kool, A.; Chakravarti, M.; Das, S.; Ghosh, S.; Thakur, L.; Khuranna, A.; Nayak, D.; Basu, B.; et al. In-vitro antiviral action of Eupatorium perfoliatum against dengue virus infection: Modulation of mTOR signaling and autophagy. J. Ethnopharmacol. 2022, 282, 114627. [Google Scholar] [CrossRef]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; Abubakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 560. [Google Scholar] [CrossRef]
- Zheng, W.; Wu, H.; Wang, T.; Zhan, S.; Liu, X. Quercetin for COVID-19 and DENGUE co-infection: A potential therapeutic strategy of targeting critical host signal pathways triggered by SARS-CoV-2 and DENV. Brief Bioinform. 2021, 22, bbab199. [Google Scholar] [CrossRef] [PubMed]
- Qamar, M.T.U.; Mumtaz, A.; Naseem, R.; Ali, A.; Fatima, T.; Jabbar, T.; Ahmad, Z.; Ashfaq, U.A. Molecular Docking Based Screening of Plant Flavonoids as Dengue NS1 Inhibitors. Bioinformation 2014, 10, 460–465. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral activity of baicalein and quercetin against the Japanese encephalitis virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Zou, M.; Liu, H.; Li, J.; Yao, X.; Chen, Y.; Ke, C.; Liu, S. Structure-activity relationship of flavonoid bifunctional inhibitors against Zika virus infection. Biochem. Pharmacol. 2020, 177, 113962. [Google Scholar] [CrossRef]
- Rojas, Á.; Del Campo, J.A.; Clement, S.; Lemasson, M.; García-Valdecasas, M.; Gil-Gómez, A.; Ranchal, I.; Bartosch, B.; Bautista, J.D.; Rosenberg, A.R.; et al. Effect of Quercetin on Hepatitis C Virus Life Cycle: From Viral to Host Targets. Sci. Rep. 2016, 6, 31777. [Google Scholar] [CrossRef]
- Fanunza, E.; Iampietro, M.; Distinto, S.; Corona, A.; Quartu, M.; Maccioni, E.; Horvat, B.; Tramontano, E. Quercetin Blocks Ebola Virus Infection by Counteracting the VP24 Interferon-Inhibitory Function. Antimicrob. Agents Chemother. 2020, 64, e00530-20. [Google Scholar] [CrossRef]
- He, Y.; Tan, S.-L.; Tareen, S.U.; Vijaysri, S.; Langland, J.O.; Jacobs, B.L.; Katze, M.G. Regulation of mRNA translation and cellular signaling by hepatitis C virus nonstructural protein NS5A. J. Virol. 2001, 75, 5090–5098. [Google Scholar] [CrossRef]
- Tellinghuisen, T.L.; Marcotrigiano, J.; Gorbalenya, A.E.; Rice, C.M. The NS5A protein of hepatitis C virus is a zinc metalloprotein. J. Biol. Chem. 2004, 279, 48576–48587. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xiao, S.; Liu, X.; Wang, L.; Ji, Q.; Mo, D.; Chen, Y. Inhibition of HSP70 reduces porcine reproductive and respiratory syndrome virus replication in vitro. BMC Microbiol. 2014, 14, 64. [Google Scholar] [CrossRef]
- Wang, J.; Hao, K.; Yu, F.; Shen, L.; Wang, F.; Yang, J.; Su, C. Field application of nanoliposomes delivered quercetin by inhibiting specific HSP70 gene expression against plant virus disease. J. Nanobiotechnol. 2022, 20, 16. [Google Scholar] [CrossRef]
- Wang, R.Y.; Stork, J.; Nagy, P.D. A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes. J. Virol. 2009, 83, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.R.; Serrano, M.A.; Efferth, T.; Alvarez, M.; Marin, J.J. Effect of cantharidin, cephalotaxine and homoharringtonine on “in vitro” models of hepatitis B virus (HBV) and bovine viral diarrhoea virus (BVDV) replication. Planta Med. 2007, 73, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.L.; Hou, C.C.; Du, C.; Zhu, J.Q. Molecular cloning and expression analysis of five heat shock protein 70 (HSP70) family members in Lateolabrax maculatus with Vibrio harveyi infection. Fish Shellfish Immunol. 2017, 60, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Kalra, E.K. Nutraceutical—Definition and introduction. AAPS PharmSci. 2003, 5, E25. [Google Scholar] [CrossRef] [PubMed]
- Aronson, J.K. Defining ‘nutraceuticals’: Neither nutritious nor pharmaceutical. Br. J. Clin. Pharmacol. 2017, 83, 8–19. [Google Scholar] [CrossRef]
- Ge, H.; Wang, Y.F.; Xu, J.; Gu, Q.; Liu, H.B.; Xiao, P.G.; Zhou, J.; Liu, Y.; Yang, Z.; Su, H. Anti-influenza agents from Traditional Chinese Medicine. Nat. Prod. Rep. 2010, 27, 1758–1780. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, N.; Bai, T.; Wang, S.; Wang, H.; Wu, Y.; Liu, Y.; Zhu, Z. New Insights into the Role and Therapeutic Potential of Heat Shock Protein 70 in Bovine Viral Diarrhea Virus Infection. Microorganisms 2023, 11, 1473. https://doi.org/10.3390/microorganisms11061473
Chen N, Bai T, Wang S, Wang H, Wu Y, Liu Y, Zhu Z. New Insights into the Role and Therapeutic Potential of Heat Shock Protein 70 in Bovine Viral Diarrhea Virus Infection. Microorganisms. 2023; 11(6):1473. https://doi.org/10.3390/microorganisms11061473
Chicago/Turabian StyleChen, Nannan, Tongtong Bai, Shuang Wang, Huan Wang, Yue Wu, Yu Liu, and Zhanbo Zhu. 2023. "New Insights into the Role and Therapeutic Potential of Heat Shock Protein 70 in Bovine Viral Diarrhea Virus Infection" Microorganisms 11, no. 6: 1473. https://doi.org/10.3390/microorganisms11061473