
The Lanthipeptide Synthetase-like Protein CA_C0082 Is an Effector of Agr Quorum Sensing in Clostridium acetobutylicum


Abstract
:1. Introduction
2. Methods
2.1. Bacterial Strains and Media
2.2. Plasmids, Primers, DNA Techniques
2.3. Construction of Mutants Using Clostron Technology
2.4. Generation of Complementation Vectors
2.5. Generation of a GusA Reporter Plasmid
2.6. Spore Assays and Detection of Granulose
2.7. Addition of Synthetic AgrD Signaling Peptide
2.8. Analysis of Fermentation Products
2.9. GusA Reporter Assay
2.10. Phylogenetic Analysis
2.11. Statistical Analysis
3. Results
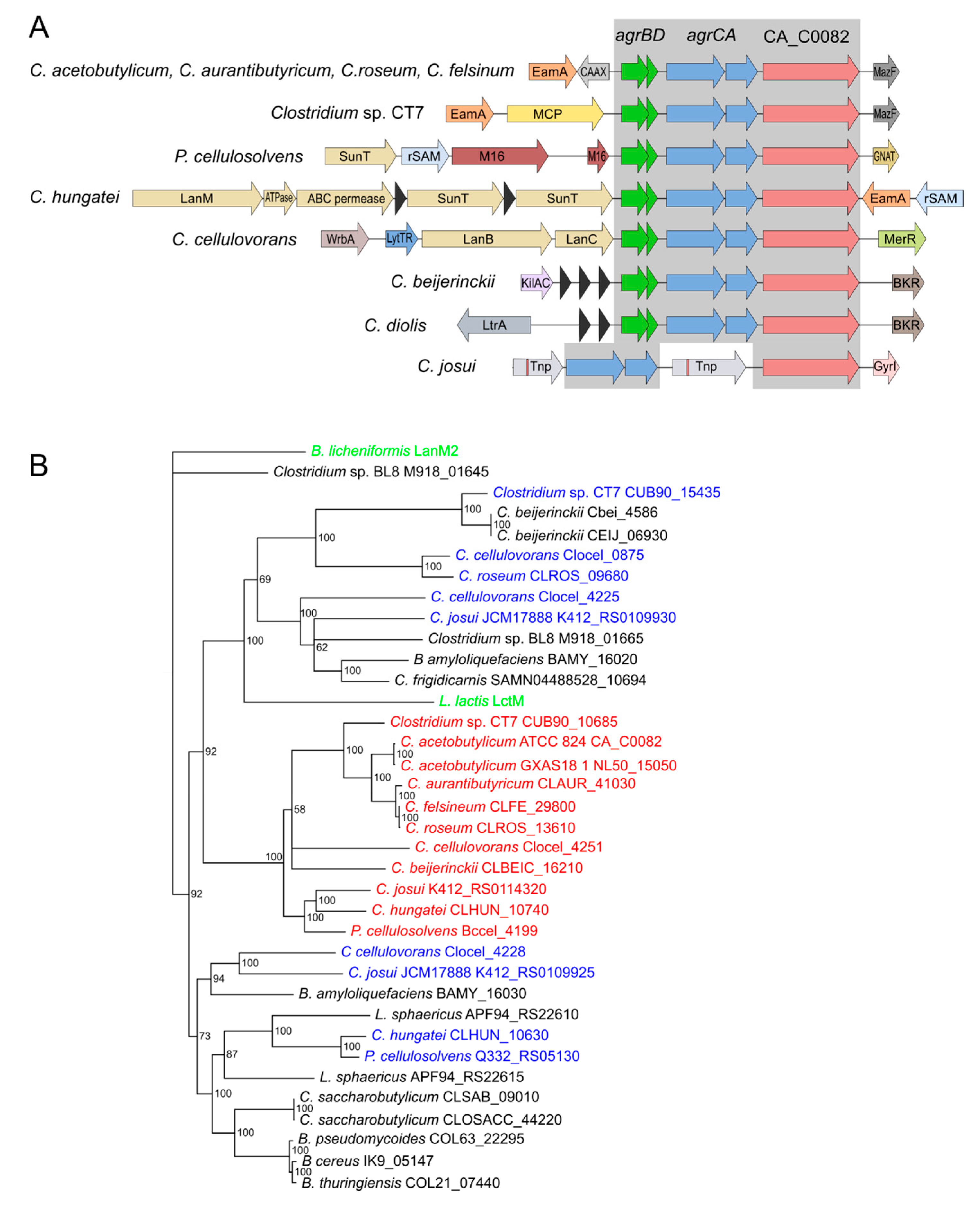
3.1. CA_C0082 and Its Close Homologs Lack a Zinc Binding Motif and Are Associated with Agr Quorum Sensing
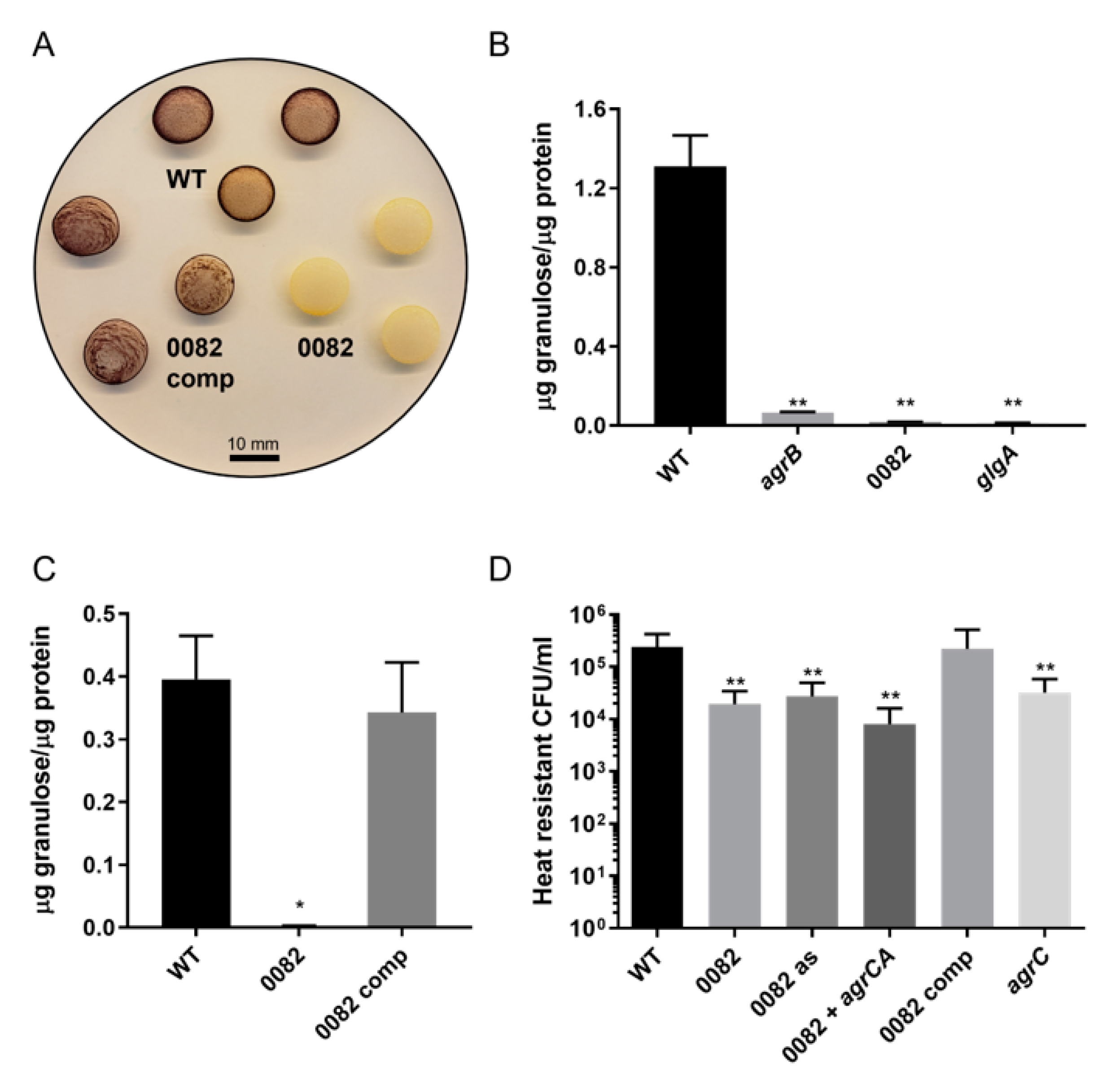
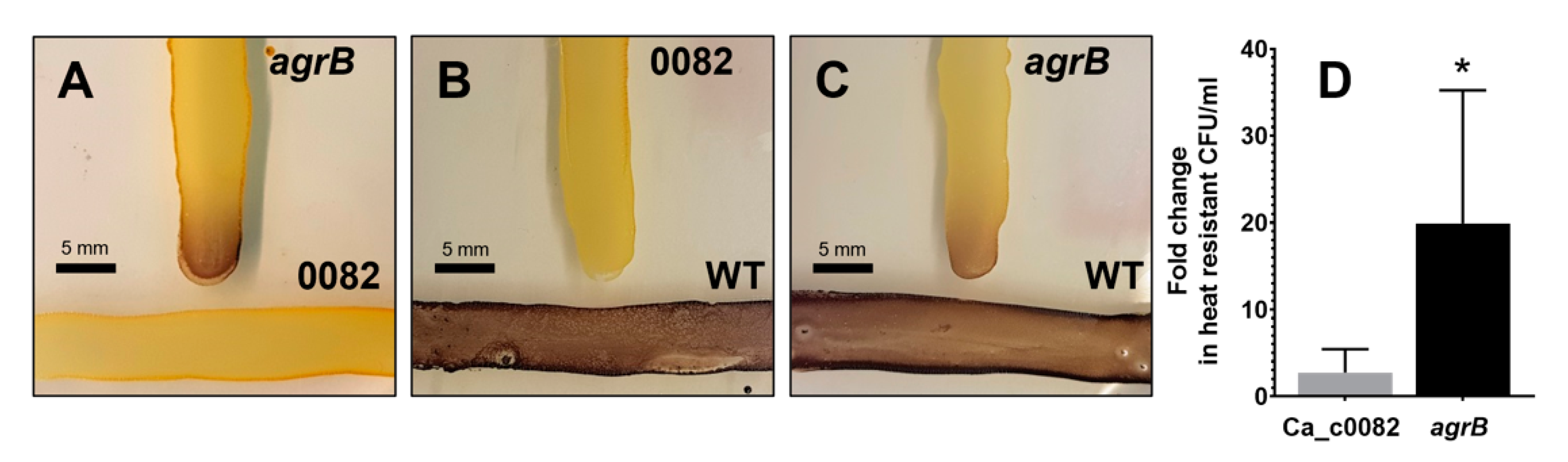
3.2. Generation and Characterization of CA_C0082 Mutants
3.3. CA_C0082 Is Not Required for Active AIP Synthesis
3.4. CA_C0082 Expression Is Controlled by Agr Quorum Sensing
3.5. Lack of Granulose Replicates the Sporulation Phenotype of Agr and CA_C0082 Mutants
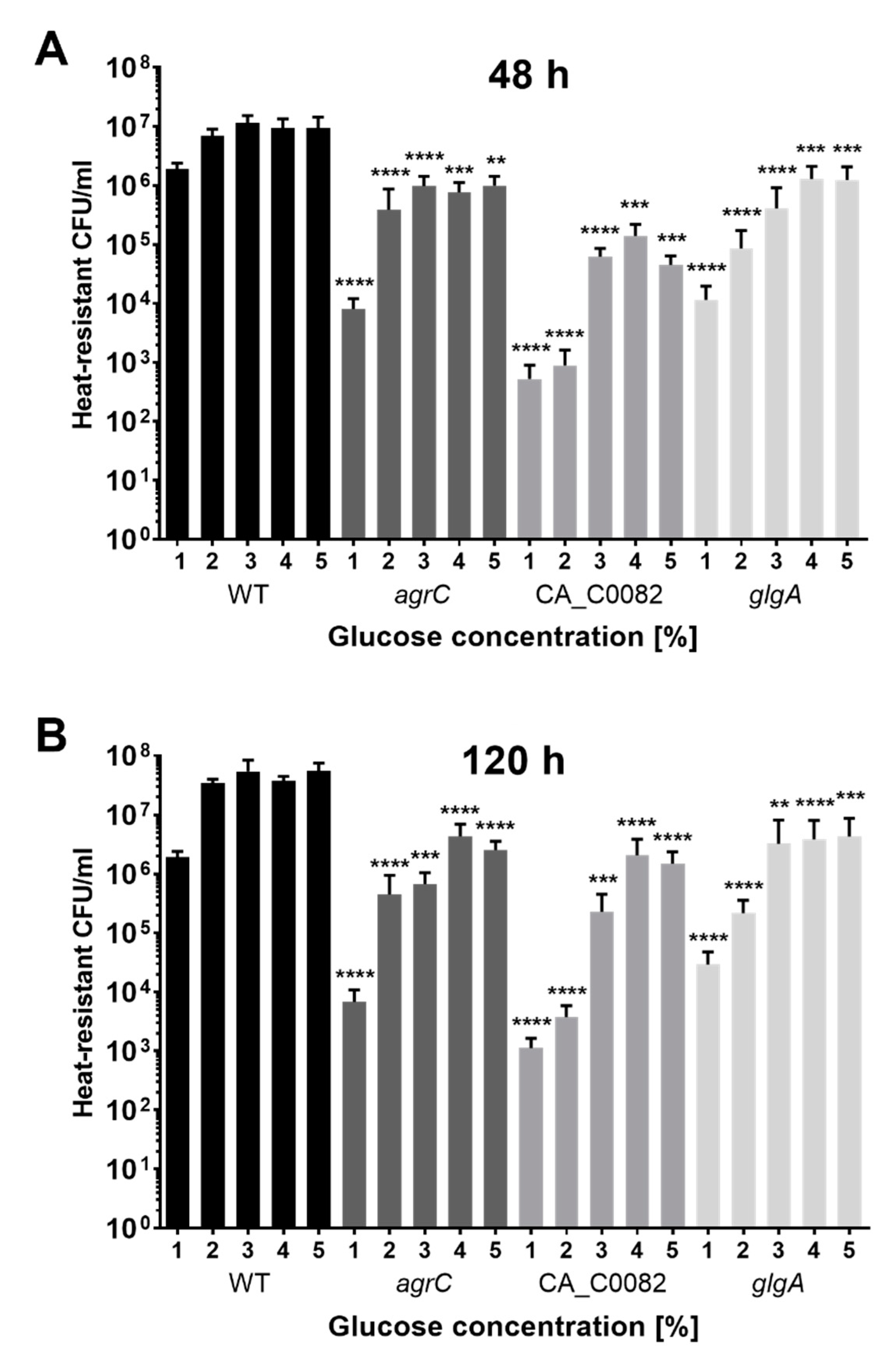
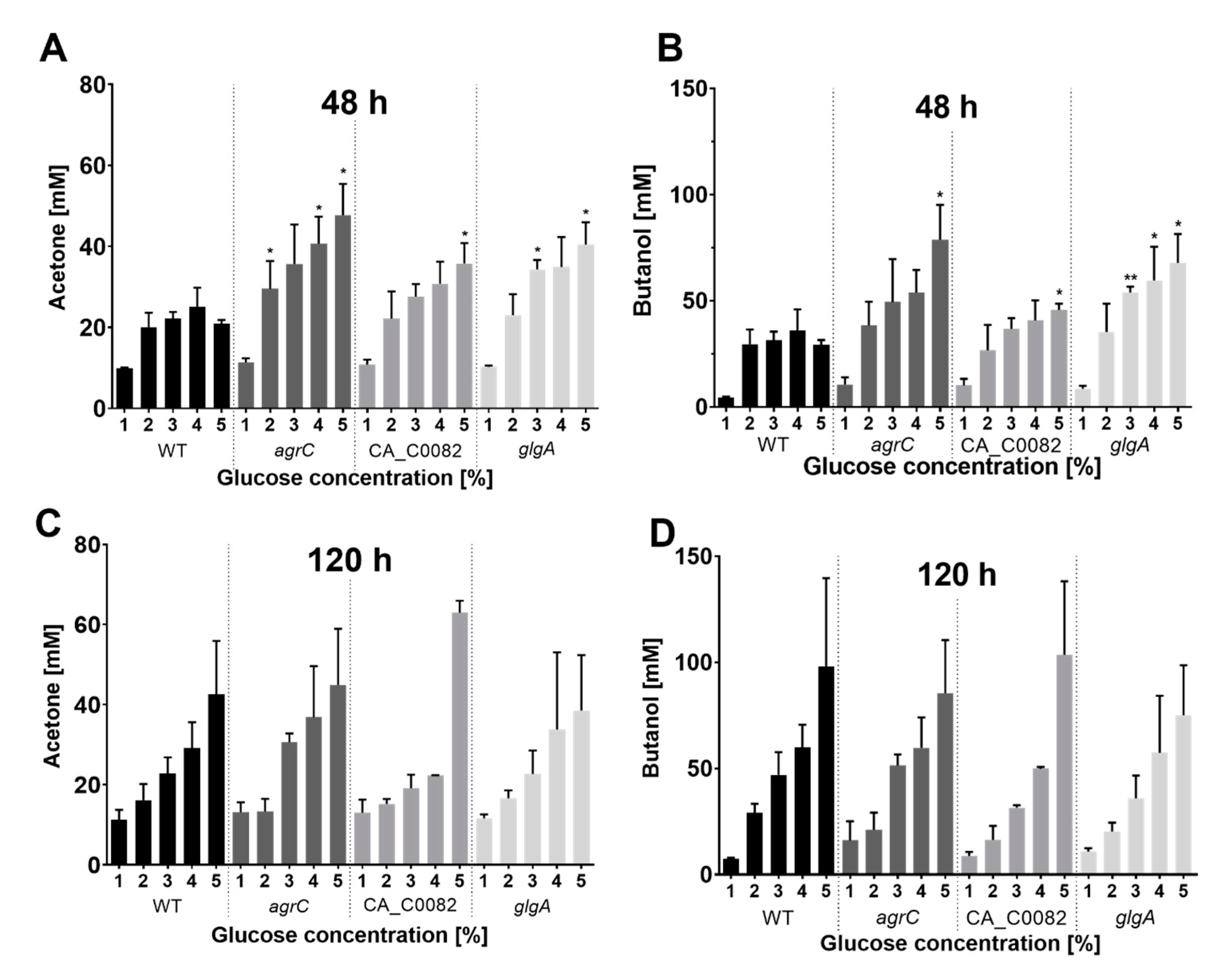
3.6. Increased Early Formation of Solvents by CA_C0082 and Agr Mutants Is Linked to the Absence of Granulose
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, D.T.; Woods, D.R. Acetone-butanol fermentation revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef]
- Al-Hinai, M.A.; Jones, S.W.; Papoutsakis, E.T. The Clostridium sporulation programs: Diversity and preservation of endospore differentiation. Microbiol. Mol. Biol. Rev. 2015, 79, 19–37. [Google Scholar] [CrossRef] [Green Version]
- Dürre, P. Physiology and sporulation in Clostridium. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reysenbach, A.L.; Ravenscroft, N.; Long, S.; Jones, D.T.; Woods, D.R. Characterization biosynthesis and regulation of granulose in Clostridium acetobutylicum. Appl. Environ. Microbiol. 1986, 52, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dürre, P.; Hollergschwandner, C. Initiation of endospore formation in Clostridium acetobutylicum. Anaerobe 2004, 10, 69–74. [Google Scholar] [CrossRef]
- Harris, L.M.; Welker, N.E.; Papoutsakis, E.T. Northern, morphological and fermentation analysis of spo0A inactivation and overexpression in Clostridium acetobutylicum ATCC 824. J. Bacteriol. 2002, 184, 3586–3597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravagnani, A.; Jennert, K.C.; Steiner, E.; Grünberg, R.; Jefferies, J.R.; Wilkinson, S.R.; Young, D.I.; Tidswell, E.C.; Brown, D.P.; Youngman, P.; et al. Spo0A directly controls the switch from acid to solvent production in solvent-forming clostridia. Mol. Microbiol. 2000, 37, 1172–1185. [Google Scholar] [CrossRef] [Green Version]
- Steiner, E.; Dago, A.E.; Young, D.I.; Heap, J.T.; Minton, N.P.; Hoch, J.A.; Young, M. Multiple orphan histidine kinases interact directly with Spo0A to control the initiation of endospore formation in Clostridium acetobutylicum. Mol. Microbiol. 2011, 80, 641–654. [Google Scholar] [CrossRef] [Green Version]
- Kotte, A.-K. RNPP-Type Quorum Sensing in Clostridium acetobutylicum. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2013. [Google Scholar]
- Steiner, E.; Scott, J.; Minton, N.P.; Winzer, K. An agr quorum sensing system that regulates granulose formation and sporulation in Clostridium acetobutylicum. Appl. Environ. Microbiol. 2012, 78, 1113–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotte, A.-K.; Severn, O.; Bean, Z.; Schwarz, K.; Minton, N.P.; Winzer, K. RRNPP-type quorum sensing affects solvent formation and sporulation in Clostridium acetobutylicum. Microbiology 2020, 166, 579–592. [Google Scholar] [CrossRef]
- Wuster, A.; Babu, M.M. Conservation and evolutionary dynamics of the agr cell-to-cell communication system across Firmicutes. J. Bacteriol. 2008, 190, 743–746. [Google Scholar] [CrossRef] [Green Version]
- Le, K.Y.; Otto, M. Quorum-sensing regulation in staphylococci—An overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Geisinger, E. Quorum sensing in staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef]
- Letzel, A.C.; Pidot, S.J.; Hertweck, C. Genome mining for ribosomally synthesized and post-translationally modified peptides (RiPPs) in anaerobic bacteria. BMC Genom. 2014, 15, 983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repka, L.M.; Chekan, J.R.; Nair, S.K.; van der Donk, W.A. Mechanistic understanding of lanthipeptide biosynthetic enzymes. Chem. Rev. 2017, 117, 5457–5520. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.W.; Morris, J.G. Oxygen and the growth and metabolism of Clostridium acetobutylicum. J. Gen. Microbiol. 1971, 68, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Carter, G.P.; Minton, N.P. The ClosTron, a universal gene knock-out system for the genus Clostridium. J. Microbiol. Methods 2007, 70, 452–464. [Google Scholar] [CrossRef]
- Ehsaan, M.; Kuit, W.; Zhang, Y.; Cartman, S.T.; Heap, J.T.; Winzer, K.; Minton, N.P. Mutant generation by allelic exchange and genome resequencing of the biobutanol organism Clostridium acetobutylicum ATCC 824. Biotechnol. Biofuels 2016, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Minton, N.P. A modular system for Clostridium shuttle plasmids. J. Microbiol. Methods 2009, 78, 79–85. [Google Scholar] [CrossRef]
- Heap, J.T.; Cartman, S.T.; Kuehne, S.A.; Cooksley, C.; Minton, N.P. ClosTron-targeted mutagenesis. Methods Mol. Biol. 2010, 646, 165–182. [Google Scholar]
- Wallace, B.D.; Roberts, A.B.; Pollet, R.M.; Ingle, J.D.; Biernat, K.A.; Pellock, S.J.; Venkatesh, M.K.; Guthrie, L.; O’Neal, S.K.; Robinson, S.J.; et al. Structure and inhibition of microbiome β-glucuronidases essential to the alleviation of cancer drug toxicity. Chem. Biol. 2015, 22, 1238–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar]
- Sukumaran, J.; Holder, M.T. DendroPy: A Python library for phylogenetic computing. Bioinformatics 2010, 26, 1569–1571. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.; Patton, G.C.; van der Donk, W.A. Mutants of the zinc ligands of lacticin 481 synthetase retain dehydration activity but have impaired cyclization activity. Biochemistry 2007, 46, 6268–6276. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yu, Y.; Vélasquez, J.E.; van der Donk, W.A. Evolution of lanthipeptide synthetases. Proc. Natl. Acad. Sci. USA 2012, 109, 18361–18366. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y.; Li, B.; Claesen, J.; Shi, Y.; Bibb, M.J.; van der Donk, W.A. Discovery of unique lanthionine synthetases reveals new mechanistic and evolutionary insights. PLoS Biol. 2010, 8, e1000339. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; van der Donk, W.A. Biosynthesis of the class III lantipeptide catenulipeptin. ACS Chem. Biol. 2012, 7, 1529–1535. [Google Scholar] [CrossRef]
- You, Y.O.; Levengood, M.R.; Ihnken, L.A.; Knowlton, A.K.; van der Donk, W.A. Lacticin 481 synthetase as a general serine/threonine kinase. ACS Chem. Biol. 2009, 4, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.O.; van der Donk, W.A. Mechanistic investigations of the dehydration reaction of lacticin 481 synthetase using site-directed mutagenesis. Biochemistry 2007, 46, 5991–6000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Gao, Y.; Zhao, F.; Wang, J.; Teng, K.; Zhang, J.; Zhong, J. Dissecting the catalytic and substrate binding activity of a class II lanthipeptide synthetase BovM. Biochem. Biophys. Res. Commun. 2014, 450, 1126–1132. [Google Scholar] [CrossRef]
- Kies, S.; Vuong, C.; Hille, M.; Peschel, A.; Meyer, C.; Götz, F.; Otto, M. Control of antimicrobial peptide synthesis by the agr quorum sensing system in Staphylococcus epidermidis: Activity of the lantibiotic epidermin is regulated at the level of precursor peptide processing. Peptides 2003, 24, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Hoover, S.E.; Perez, A.J.; Tsui, H.-C.T.; Sinha, D.; Smiley, D.L.; DiMarchi, R.D.; Winkler, M.E.; Lazazzera, B.A. A new quorum sensing system (TprA/PhrA) for Streptococcus pneumoniae D39 that regulates a lantibiotic biosynthesis gene cluster. Mol. Microbiol. 2015, 97, 229–243. [Google Scholar] [CrossRef]
- Kuipers, O.P.; Beerthuyzen, M.M.; de Ruyter, P.G.; Luesink, E.J.; de Vos, W.M. Autoregulation of nisin biosynthesis in Lactococcus lactis by signal transduction. J. Biol. Chem. 1995, 270, 27299–27304. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T. Applied acetone-butanol fermentation. In Clostridia: Biotechnology and Medical Applications; Bahl, H., Dürre, P., Eds.; Wiley-VCH: Weinheim, Germany, 2001; pp. 125–168. [Google Scholar]
- Hoch, J.A. Regulation of the phosphorelay and the initiation of sporulation in Bacillus subtilis. Annu. Rev. Microbiol. 1993, 47, 441–465. [Google Scholar] [CrossRef]
- Grossman, A.D. Genetic networks controlling the initiation of sporulation and the development of genetic competence in Bacillus subtilis. Annu. Rev. Genet. 1995, 29, 477–508. [Google Scholar] [CrossRef]
- Lazazzera, B.A. Quorum sensing and starvation: Signals for entry into stationary phase. Curr. Opin. Microbiol. 2000, 3, 177–182. [Google Scholar] [CrossRef]
- Rainey, F.A.; Hollen, B.J.; Small, A.; Genus, I. Clostridium. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; de Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; Volume 3, pp. 738–828. [Google Scholar]
- Horino, H.; Fujita, T.; Tonouchi, A. Description of Anaerobacterium chartisolvens gen. nov. sp. nov., an obligately anaerobic bacterium from Clostridium rRNA cluster III isolated from soil of a Japanese rice field, and reclassification of Bacteroides cellulosolvens Murray et al. 1984 as Pseudobacteroides cellulosolvens gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1296–1303. [Google Scholar]
- Tracy, B.P.; Jones, S.W.; Papoutsakis, E.T. Inactivation of σE and σG in Clostridium acetobutylicum illuminates their roles in clostridial-cell-form biogenesis granulose synthesis solventogenesis, and spore morphogenesis. J. Bacteriol. 2011, 193, 1414–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, N.A.; Kim, S.J.; Li, J.S.; Cai, W.; Koshino, H.; Zhang, W. The industrial anaerobe Clostridium acetobutylicum uses polyketides to regulate butanol production and differentiation. Nat. Commun. 2017, 15, 1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Properties | Source/Reference |
|---|---|---|
| E. coli Top10 | F-mcrA Δ(mrr-hsdRMS-mcrBC) Φ80lacZΔM15 ΔlacX74 recA1 araD139 Δ(ara leu)7697 galU galK rpsL (StrR) endA1 nupG | Invitrogen |
| E. coli Top 10 pAN2 | E. coli Top 10 with methylation plasmid pAN2 containing the ϕ3TI methyltransferase | [18] |
| C. acetobutylicum ATCC 824 | C. acetobutylicum ATCC 824 wild type | Prof. Hubert Bahl, University of Rostock (COSMIC-strain) |
| C. acetobutylicum CA_C0082::CTermB_s | C. acetobutylicum ATCC 824 CA_C0082 ClosTron mutant; ClosTron inserted in “sense” orientation between base pairs 1119/1120 | This study |
| C. acetobutylicum CA_C0082::CTermB_as | C. acetobutylicum ATCC 824 CA_C0082 ClosTron mutant; ClosTron inserted in “antisense” orientation between base pairs 538/539 | This study |
| C. acetobutylicum agrA::CTermB | C. acetobutylicum ATCC 824 agrC ClosTron mutant; unable to mediate a quorum sensing response | [10] |
| C. acetobutylicum agrB::CTermB | C. acetobutylicum ATCC 824 agrB ClosTron mutant; unable to produce AIP (quorum sensing peptide) | [10] |
| C. acetobutylicum agrC::CTermB | C. acetobutylicum ATCC 824 agrC ClosTron mutant; unable to sense AIP (quorum sensing peptide) | [10] |
| C. acetobutylicum ΔglgA | C. acetobutylicum ATCC 824 glycogen synthase (glgA) in-frame deletion mutant | [19] |
| C. acetobutylicum pMTL85141 | ATCC 824 wild type with empty pMTL85141 vector | This study |
| C. acetobutylicum CA_C0082::CTermB_s pMTL85141 | CA_C0082 mutant with empty pMTL85141 vector | This study |
| C. acetobutylicum CA_C0082::CTermB_s pMTL85141-CA_C0082 | CA_C0082 mutant with pMTL85141-CA_C0082 complementation vector (native CA_C0082 promoter) | This study |
| C. acetobutylicum CA_C0082::CTermB_s pMTL85141-agrCA | CA_C0082 mutant with pMTL8514-agrCA complementation vector | This study |
| C. acetobutylicum agrA::CTermB pMTL85143 | agrA mutant with empty pMTL85143 (ferredoxin promoter) | This study |
| C. acetobutylicum agrA::CTermB pMTL85143-CA_C0082 | agrA mutant with pMTL85143-CA_C0082 vector (ferredoxin promoter) | This study |
| C. acetobutylicum pMTL-JL1-PCA_C0082 | ATCC 824 wild type expressing CA_C0082-gusA transcriptional reporter fusion | This study |
| C. acetobutylicum agrA::CTermB pMTL-JL1-PCA_C0082 | agrA mutant expressing CA_C0082-gusA transcriptional reporter fusion | This study |
| C. acetobutylicum agrB::CTermB pMTL-JL1-PCA_C0082 | agrB mutant expressing CA_C0082-gusA transcriptional reporter fusion | This study |
| Plasmid | Relevant Properties | Source |
|---|---|---|
| pAN2 | Plasmid containing ϕ3TI methyltransferase | [18] |
| pMTL007C-E2::CA_C0082-538|539A | ClosTron plasmid retargeted to CA_C0082 1 | This study |
| pMTL007C-E2::CA_C0082-1119|1120S | ClosTron plasmid retargeted to CA_C0082 1 | This study |
| pMTL85141 | Clostridium modular plasmid containing a catPresistance marker | [20] |
| pMTL85143 | pMTL85141 with C. sporogenes ferredoxin promoter upstream of a multiple cloning site | Ref. [11], Dr Ying Zhang, Univ. of Nottingham |
| pMTL85141-CA_C0082 | pMTL85141 containing CA_C0082 coding region and 88 bp non-coding upstream promoter region | This study |
| pMTL85141-agrCA | pMTL85141 containing the C. acetobutylicum agrCA operon under control of its native promoter | [10] |
| pMTL85143-CA_C0082 | pMTL85143 containing the CA_C0082 coding region under control of consitutive C. sporogenes ferredoxin promoter | This study |
| pMTL-JL1 | pMTL85143 derivative conatining the Streptococcus agalactiae gusA gene under control of the C. sporogenes ferredoxin promoter | Dr Jessica Locke, Univ. of Nottingham |
| pMTL-JL1-PCA_C0082 | CA_C0082 β-glucuronidase reporter; pMTL-JL1 derivative containing a transcriptional fusion of the C. acetobutylicum CA_C0082 promoter region and Streptococcus agalactiae gusA | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humphreys, J.R.; Bean, Z.; Twycross, J.; Winzer, K. The Lanthipeptide Synthetase-like Protein CA_C0082 Is an Effector of Agr Quorum Sensing in Clostridium acetobutylicum. Microorganisms 2023, 11, 1460. https://doi.org/10.3390/microorganisms11061460
Humphreys JR, Bean Z, Twycross J, Winzer K. The Lanthipeptide Synthetase-like Protein CA_C0082 Is an Effector of Agr Quorum Sensing in Clostridium acetobutylicum. Microorganisms. 2023; 11(6):1460. https://doi.org/10.3390/microorganisms11061460
Chicago/Turabian StyleHumphreys, Jonathan R., Zak Bean, Jamie Twycross, and Klaus Winzer. 2023. "The Lanthipeptide Synthetase-like Protein CA_C0082 Is an Effector of Agr Quorum Sensing in Clostridium acetobutylicum" Microorganisms 11, no. 6: 1460. https://doi.org/10.3390/microorganisms11061460